Los dinosaurios son variados desde el punto de vista taxonómico , morfológico y ecológico . Las aves, con más de 11.000 especies vivas , se encuentran entre los grupos de vertebrados más diversos. Utilizando evidencia fósil, los paleontólogos han identificado más de 900 géneros distintos y más de 1.000 especies diferentes de dinosaurios no aviares. Los dinosaurios están representados en todos los continentes tanto por especies existentes (aves) como por restos fósiles. Durante la primera mitad del siglo XX, antes de que las aves fueran reconocidas como dinosaurios, la mayor parte de la comunidad científica creía que los dinosaurios eran lentos y de sangre fría . Sin embargo, la mayoría de las investigaciones realizadas desde la década de 1970 han indicado que los dinosaurios eran animales activos con metabolismos elevados y numerosas adaptaciones para la interacción social. Algunos eran herbívoros , otros carnívoros . La evidencia sugiere que todos los dinosaurios ponían huevos y que la construcción de nidos era un rasgo compartido por muchos dinosaurios, tanto aviares como no aviares.
Si bien los dinosaurios eran ancestralmente bípedos , muchos grupos extintos incluían especies cuadrúpedos , y algunos pudieron cambiar entre estas posturas. Las estructuras de exhibición elaboradas, como cuernos o crestas, son comunes a todos los grupos de dinosaurios, y algunos grupos extintos desarrollaron modificaciones esqueléticas , como armaduras óseas y espinas . Si bien el linaje aviar (aves) de los dinosaurios que sobrevive en la actualidad es generalmente pequeño debido a las limitaciones del vuelo, muchos dinosaurios prehistóricos (avianos y no aviares) tenían cuerpos grandes: se estima que los dinosaurios saurópodos más grandes alcanzaron longitudes de 39,7 metros (130 pies) y alturas de 18 m (59 pies) y fueron los animales terrestres más grandes de todos los tiempos. La idea errónea de que los dinosaurios no aviares eran uniformemente gigantescos se basa en parte en el sesgo de preservación , ya que es más probable que los huesos grandes y resistentes duren hasta que se fosilicen. Muchos dinosaurios eran bastante pequeños, algunos medían unos 50 centímetros (20 pulgadas) de largo.
Los primeros fósiles de dinosaurios fueron reconocidos a principios del siglo XIX, y Sir Richard Owen acuñó el nombre "dinosaurio" (que significa "lagarto terrible") en 1842 para referirse a estos "grandes lagartos fósiles". [7] [8] [9] Desde entonces, los esqueletos de dinosaurios fósiles montados han sido atracciones importantes en museos de todo el mundo, y los dinosaurios se han convertido en una parte duradera de la cultura popular . Los grandes tamaños de algunos dinosaurios, así como su naturaleza aparentemente monstruosa y fantástica, han asegurado su aparición habitual en libros y películas más vendidos, como Jurassic Park . El persistente entusiasmo público por los animales ha resultado en una importante financiación para la ciencia de los dinosaurios, y los medios de comunicación cubren periódicamente los nuevos descubrimientos.
La investigación realizada por Matthew G. Baron, David B. Norman y Paul M. Barrett en 2017 sugirió una revisión radical de la sistemática de los dinosaurios. Análisis filogenético de Baron et al. recuperó los Ornithischia por estar más cerca de los Theropoda que los Sauropodomorpha, en contraposición a la unión tradicional de terópodos con sauropodomorfos. Esto haría que los saurópodos y sus parientes quedaran fuera de los dinosaurios tradicionales, por lo que redefinieron Dinosauria como el último ancestro común de Triceratops horridus , Passer domesticus y Diplodocus carnegii , y todos sus descendientes, para garantizar que los saurópodos y sus parientes sigan incluidos como dinosaurios. También resucitaron el clado Ornithoscelida para referirse al grupo que contiene Ornithischia y Theropoda. [15] [16]
Usando una de las definiciones anteriores, los dinosaurios pueden describirse generalmente como arcosaurios con las extremidades traseras erguidas debajo del cuerpo . [17] Otros animales prehistóricos, incluidos pterosaurios , mosasaurios , ictiosaurios , plesiosaurios y dimetrodon , aunque a menudo se los concibe popularmente como dinosaurios, no están clasificados taxonómicamente como dinosaurios. [18] Los pterosaurios están relacionados lejanamente con los dinosaurios, siendo miembros del clado Ornithodira . Los otros grupos mencionados son, al igual que los dinosaurios y los pterosaurios, miembros de Sauropsida (el clado de reptiles y aves), excepto Dimetrodon (que es un sinápsido ). Ninguno de ellos tenía la postura erguida de las extremidades traseras característica de los verdaderos dinosaurios. [19]
Los dinosaurios fueron los vertebrados terrestres dominantes de la Era Mesozoica , especialmente de los períodos Jurásico y Cretácico. Otros grupos de animales estaban restringidos en tamaño y nichos; Los mamíferos , por ejemplo, rara vez superaban el tamaño de un gato doméstico, y generalmente eran carnívoros del tamaño de roedores y presas pequeñas. [20] Los dinosaurios siempre han sido reconocidos como un grupo extremadamente variado: se han identificado con seguridad más de 900 géneros de dinosaurios no aviares (2018) con 1124 especies (2016). Las estimaciones sitúan el número total de géneros de dinosaurios conservados en el registro fósil en 1850, casi el 75% aún sin descubrir, [21] [22] [23] y el número que alguna vez existió (dentro o fuera del registro fósil) en 3.400. [24] Una estimación de 2016 sitúa el número de especies de dinosaurios que viven en el Mesozoico entre 1.543 y 2.468, [25] [26] en comparación con el número de aves modernas (dinosaurios aviares) en 10.806 especies. [27]
Los dinosaurios extintos, así como las aves modernas, incluyen géneros que son herbívoros y otros carnívoros, incluidos los que se alimentan de semillas, los que se alimentan de peces, los insectívoros y los omnívoros. Si bien los dinosaurios eran ancestralmente bípedos (como lo son todas las aves modernas), algunos evolucionaron hasta convertirse en cuadrúpedos y otros, como Anchisaurus e Iguanodon , podían caminar con la misma facilidad sobre dos o cuatro patas. Las modificaciones craneales, como cuernos y crestas, son rasgos comunes de los dinosaurios, y algunas especies extintas tenían armadura ósea. Aunque los géneros más conocidos destacan por su gran tamaño, muchos dinosaurios mesozoicos eran del tamaño de los humanos o más pequeños, y las aves modernas son generalmente de tamaño pequeño. Los dinosaurios hoy habitan en todos los continentes, y los fósiles muestran que habían alcanzado una distribución global a más tardar en la época del Jurásico Temprano . [28] Las aves modernas habitan en la mayoría de los hábitats disponibles, desde terrestres hasta marinos, y hay evidencia de que algunos dinosaurios no aviares (como el Microraptor ) podían volar o al menos planear, y otros, como los espinosáuridos , tenían hábitos semiacuáticos . [29]
Características anatómicas distintivas.
Si bien los descubrimientos recientes han hecho que sea más difícil presentar una lista universalmente aceptada de sus características distintivas, casi todos los dinosaurios descubiertos hasta ahora comparten ciertas modificaciones en el esqueleto ancestral del arcosaurio, o son claramente descendientes de dinosaurios más antiguos que muestran estas modificaciones. Aunque algunos grupos posteriores de dinosaurios presentaron versiones modificadas de estos rasgos, se consideran típicos de Dinosauria; los primeros dinosaurios los tenían y los transmitieron a sus descendientes. Estas modificaciones, que se originan en el ancestro común más reciente de un determinado grupo taxonómico, se denominan sinapomorfias de dicho grupo. [30]
Diagrama etiquetado de un cráneo de arcosaurio típico, el cráneo de Dromaeosaurus
Una evaluación detallada de las interrelaciones de los arcosaurios realizada por Sterling Nesbitt [31] confirmó o encontró las siguientes doce sinapomorfias inequívocas, algunas conocidas previamente:
En el cráneo , hay una fosa supratemporal (excavación) delante de la fenestra supratemporal , la abertura principal en el techo posterior del cráneo.
Epífisis , apófisis que apuntan oblicuamente hacia atrás en las esquinas superiores traseras de las vértebras anteriores (frontales) del cuello , detrás del atlas y el eje , las dos primeras vértebras del cuello.
Vértice de una cresta deltopectoral (una proyección en la que se insertan los músculos deltopectorales ) ubicada en o más del 30% de la longitud del húmero (hueso de la parte superior del brazo)
Radio , un hueso del antebrazo, más corto que el 80% de la longitud del húmero.
El cuarto trocánter (proyección donde el músculo caudofemoral se inserta en el eje posterior interno) en el fémur (hueso del muslo) es un reborde afilado
Cuarto trocánter asimétrico, con margen distal inferior que forma un ángulo más pronunciado con respecto a la diáfisis.
En el astrágalo y el calcáneo , los huesos superiores del tobillo, la faceta articular proximal, la superficie de conexión superior, pues el peroné ocupa menos del 30% del ancho transversal del elemento.
Los exoccipitales (huesos en la parte posterior del cráneo) no se juntan a lo largo de la línea media en el piso de la cavidad endocraneal, el espacio interno de la caja del cráneo.
En la pelvis, las superficies articulares proximales del isquion con el ilion y el pubis están separadas por una gran superficie cóncava (en la parte superior del isquion, una parte de la articulación abierta de la cadera se encuentra entre los contactos con el hueso púbico y el ilion)
La cresta cnemial de la tibia (parte que sobresale de la superficie superior de la tibia) se arquea anterolateralmente (se curva hacia el frente y el lado externo)
Distinta cresta orientada proximodistalmente (vertical) presente en la cara posterior del extremo distal de la tibia (la superficie posterior del extremo inferior de la tibia)
Superficie articular cóncava para el peroné del calcáneo (la superficie superior del calcáneo, donde toca el peroné, tiene un perfil hueco)
Nesbitt encontró varias sinapomorfias potenciales adicionales y descartó varias sinapomorfias sugeridas anteriormente. Algunos de estos también están presentes en los silesáuridos , que Nesbitt recuperó como grupo hermano de Dinosauria, incluido un gran trocánter anterior, metatarsianos II y IV de longitud subigual, contacto reducido entre el isquion y el pubis, la presencia de una cresta cnemial en la tibia y de un proceso ascendente en el astrágalo, y muchos otros. [10]
Articulaciones de la cadera y posturas de las patas traseras de: (de izquierda a derecha) reptiles típicos (extendidos), dinosaurios y mamíferos (erectos) y rauisuquios (erectos en forma de pilar)
Los dinosaurios comparten una variedad de otras características esqueléticas. Sin embargo, debido a que son comunes a otros grupos de arcosaurios o no estaban presentes en todos los primeros dinosaurios, estas características no se consideran sinapomorfias. Por ejemplo, como diápsidos , los dinosaurios ancestralmente tenían dos pares de fenestras infratemporales (aberturas en el cráneo detrás de los ojos) y, como miembros del grupo de diápsidos Archosauria, tenían aberturas adicionales en el hocico y la mandíbula inferior. [32] Además, ahora se sabe que varias características que antes se pensaba que eran sinapomorfias aparecieron antes que los dinosaurios, o estuvieron ausentes en los primeros dinosaurios y evolucionaron de forma independiente por diferentes grupos de dinosaurios. Estos incluyen una escápula u omóplato alargada; un sacro compuesto por tres o más vértebras fusionadas (tres se encuentran en algunos otros arcosaurios, pero sólo dos en Herrerasaurus ); [10] y un acetábulo perforado , o cavidad de la cadera, con un agujero en el centro de su superficie interior (cerrado en Saturnalia tupiniquim , por ejemplo). [33] [34] Otra dificultad para determinar las características distintivas de los dinosaurios es que los primeros dinosaurios y otros arcosaurios de la época del Triásico Tardío a menudo son poco conocidos y eran similares en muchos aspectos; Estos animales a veces han sido identificados erróneamente en la literatura. [35]
Los dinosaurios se paran con sus extremidades traseras erguidas de una manera similar a la mayoría de los mamíferos modernos , pero distintos de la mayoría de los otros reptiles, cuyas extremidades se extienden hacia ambos lados. [36] Esta postura se debe al desarrollo de un hueco orientado lateralmente en la pelvis (generalmente una cavidad abierta) y una correspondiente cabeza distinta orientada hacia adentro en el fémur. [37] Su postura erguida permitió a los primeros dinosaurios respirar fácilmente mientras se movían, lo que probablemente permitió niveles de resistencia y actividad que superaron los de los reptiles "extendidos" . [38] Las extremidades erectas probablemente también ayudaron a apoyar la evolución del gran tamaño al reducir las tensiones de flexión en las extremidades. [39] Algunos arcosaurios no dinosaurios, incluidos los rauisuquios , también tenían extremidades erectas, pero lo lograban mediante una configuración de "pilar erecto" de la articulación de la cadera, donde en lugar de tener una proyección desde el fémur insertado en una cavidad en la cadera, el El hueso pélvico superior se rotó para formar un estante sobresaliente. [39]
historia del estudio
Historia precientífica
Los fósiles de dinosaurios se conocen desde hace milenios, aunque no se reconocía su verdadera naturaleza. Los chinos los consideraban huesos de dragón y los documentaron como tales. Por ejemplo, Huayang Guo Zhi (華陽國志), un diccionario geográfico compilado por Chang Qu (常璩) durante la dinastía Jin Occidental (265-316), informó del descubrimiento de huesos de dragón en Wucheng, en la provincia de Sichuan . [40] Los aldeanos del centro de China llevan mucho tiempo desenterrando "huesos de dragón" fosilizados para utilizarlos en medicinas tradicionales . [41] En Europa , generalmente se creía que los fósiles de dinosaurios eran restos de gigantes y otras criaturas bíblicas . [42]
Las descripciones académicas de lo que ahora se reconocerían como huesos de dinosaurio aparecieron por primera vez a finales del siglo XVII en Inglaterra. Parte de un hueso, que ahora se sabe que fue el fémur de un Megalosaurus , [43] fue recuperado de una cantera de piedra caliza en Cornwell cerca de Chipping Norton , Oxfordshire, en 1676. El fragmento fue enviado a Robert Plot , profesor de química en la Universidad. de Oxford y primer conservador del Museo Ashmolean , que publicó una descripción en su The Natural History of Oxford-shire (1677). [44] Identificó correctamente el hueso como la extremidad inferior del fémur de un animal grande, y reconoció que era demasiado grande para pertenecer a cualquier especie conocida. Por lo tanto, concluyó que se trataba del fémur de un humano enorme, tal vez un titán u otro tipo de gigante que aparece en las leyendas. [45] [46] Edward Lhuyd , amigo de Sir Isaac Newton , publicó Lithophylacii Britannici ichnographia (1699), el primer tratamiento científico de lo que ahora sería reconocido como un dinosaurio cuando describió y nombró a un diente de saurópodo , " Rutellum impicatum " . , [47] [48] que se había encontrado en Caswell, cerca de Witney , Oxfordshire. [49]
Entre 1815 y 1824, el reverendo William Buckland , primer lector de Geología de la Universidad de Oxford, recolectó más huesos fosilizados de Megalosaurus y se convirtió en la primera persona en describir un dinosaurio no aviar en una revista científica . [43] [50] El segundo género de dinosaurios no aviares identificado, Iguanodon , fue, según la leyenda, descubierto en 1822 por Mary Ann Mantell , la esposa del geólogo inglés Gideon Mantell , quien de hecho había requerido restos años antes. Gideon Mantell reconoció similitudes entre sus fósiles y los huesos de las iguanas modernas . Publicó sus hallazgos en 1825. [51] [52]
El estudio de estos "grandes lagartos fósiles" pronto adquirió gran interés para los científicos europeos y americanos, y en 1842 el paleontólogo inglés Sir Richard Owen acuñó el término "dinosaurio", usándolo para referirse a la "tribu distinta o suborden de Reptiles Saurios" que entonces estaban siendo reconocidos en Inglaterra y en todo el mundo. [7] [8] [9] [53] [54] El término se deriva del griego antiguo δεινός (deinos) 'terrible, potente o terriblemente grande', y σαῦρος (sauros) 'lagarto o reptil'. [53] [55] Aunque el nombre taxonómico a menudo se ha interpretado como una referencia a los dientes, garras y otras características temibles de los dinosaurios, Owen pretendía que también evocara su tamaño y majestuosidad. [56] Owen reconoció que los restos que se habían encontrado hasta ahora, Iguanodon , Megalosaurus e Hylaeosaurus , compartían características distintivas, y por eso decidió presentarlos como un grupo taxonómico distinto. Según aclaró el geólogo e historiador británico Hugh Torrens, Owen había dado una presentación sobre reptiles fósiles ante la Asociación Británica para el Avance de la Ciencia en 1841, pero informes de la época muestran que Owen no mencionó la palabra "dinosaurio", ni reconoció a los dinosaurios. como un grupo distinto de reptiles en su dirección. Presentó los Dinosaurios sólo en la versión de texto revisada de su charla publicada en abril de 1842. [7] [8] Con el respaldo del Príncipe Alberto , esposo de la Reina Victoria , Owen estableció el Museo de Historia Natural de Londres , para exhibir el patrimonio nacional. colección de fósiles de dinosaurios y otras exhibiciones biológicas y geológicas. [57]
Descubrimientos en América del Norte
En 1858, William Parker Foulke descubrió el primer dinosaurio americano conocido, en pozos de margas en la pequeña ciudad de Haddonfield, Nueva Jersey . (Aunque ya se habían encontrado fósiles antes, no se había discernido correctamente su naturaleza). La criatura se llamó Hadrosaurus foulkii . Fue un hallazgo extremadamente importante: Hadrosaurus fue uno de los primeros esqueletos de dinosaurio casi completos encontrados ( el primero fue en 1834, en Maidstone, Inglaterra ), y era claramente una criatura bípeda. Este fue un descubrimiento revolucionario ya que, hasta ese momento, la mayoría de los científicos habían creído que los dinosaurios caminaban sobre cuatro patas, como otros lagartos. Los descubrimientos de Foulke provocaron una ola de interés por los dinosaurios en los Estados Unidos, conocida como manía por los dinosaurios. [58]
La manía de los dinosaurios quedó ejemplificada por la feroz rivalidad entre Edward Drinker Cope y Othniel Charles Marsh , quienes corrieron para ser los primeros en encontrar nuevos dinosaurios en lo que se conoció como las Guerras de los Huesos . Esta lucha entre los dos científicos duró más de 30 años y terminó en 1897 cuando Cope murió después de gastar toda su fortuna en la caza de dinosaurios. Muchos ejemplares valiosos de dinosaurios resultaron dañados o destruidos debido a los duros métodos de la pareja: sus excavadores, por ejemplo, utilizaban a menudo dinamita para desenterrar huesos. Los paleontólogos modernos encontrarían estos métodos toscos e inaceptables, ya que las voladuras destruyen fácilmente la evidencia fósil y estratigráfica. A pesar de sus métodos poco refinados, las contribuciones de Cope y Marsh a la paleontología fueron enormes: Marsh desenterró 86 nuevas especies de dinosaurios y Cope descubrió 56, un total de 142 nuevas especies. La colección de Cope se encuentra ahora en el Museo Americano de Historia Natural de la ciudad de Nueva York, mientras que la de Marsh se encuentra en el Museo Peabody de Historia Natural de la Universidad de Yale . [59]
La Segunda Guerra Mundial provocó una pausa en la investigación paleontológica; Después de la guerra, la atención de la investigación también se desvió cada vez más hacia los mamíferos fósiles en lugar de los dinosaurios, a los que se consideraba lentos y de sangre fría. [60] [61] Sin embargo, a finales de la década de 1960, el campo de la investigación de los dinosaurios experimentó un aumento en la actividad que continúa. [62] Varios estudios fundamentales condujeron a esta actividad. Primero, John Ostrom descubrió el terópodo dromeosáurido parecido a un pájaro Deinonychus y lo describió en 1969. Su anatomía indicaba que era un depredador activo que probablemente era de sangre caliente, en marcado contraste con la imagen entonces predominante de los dinosaurios. [60] Al mismo tiempo, Robert T. Bakker publicó una serie de estudios que también abogaban por estilos de vida activos en los dinosaurios basados en evidencia anatómica y ecológica (ver § Fisiología), [63] [64] que se resumieron posteriormente en su libro de 1986 The Dinosaur Herejías . [sesenta y cinco]
Las nuevas revelaciones fueron respaldadas por un aumento en los descubrimientos de dinosaurios. Los paleontólogos que trabajan en regiones previamente inexploradas, como la India, América del Sur, Madagascar, la Antártida y, más significativamente, China, han realizado nuevos descubrimientos importantes sobre dinosaurios. Entre terópodos, sauropodomorfos y ornitisquios, el número de géneros nombrados comenzó a aumentar exponencialmente en la década de 1990. [21] A partir de 2008, [actualizar]cada año se nombraban más de 30 nuevas especies de dinosaurios. [66] Al menos los sauropodomorfos experimentaron un aumento adicional en el número de especies nombradas en la década de 2010, con un promedio de 9,3 nuevas especies nombradas cada año entre 2009 y 2020. Como consecuencia, se nombraron más sauropodomorfos entre 1990 y 2020 que en todos los años anteriores combinados. [67] Estas nuevas localidades también condujeron a mejoras en la calidad general de los especímenes, y las nuevas especies reciben cada vez más nombres no de fósiles fragmentados sino de esqueletos más completos, a veces de varios individuos. Mejores especímenes también condujeron a que las nuevas especies fueran invalidadas con menos frecuencia. [66] Las localidades asiáticas han producido los especímenes de terópodos más completos, [68] mientras que las localidades de América del Norte han producido los especímenes de sauropodomorfos más completos. [67]
Antes del renacimiento de los dinosaurios, los dinosaurios se clasificaban principalmente utilizando el sistema tradicional basado en rangos de la taxonomía de Linneo . El renacimiento también estuvo acompañado por la aplicación cada vez más extendida de la cladística , un método de clasificación más objetivo basado en la ascendencia y los rasgos compartidos, que ha demostrado ser tremendamente útil en el estudio de la sistemática y la evolución de los dinosaurios. El análisis cladístico, entre otras técnicas, ayuda a compensar un registro fósil a menudo incompleto y fragmentario. [69] [70] Los libros de referencia que resumen el estado de la investigación de los dinosaurios, como The Dinosauria de David B. Weishampel y sus colegas , hicieron que el conocimiento fuera más accesible [71] y estimularon un mayor interés en la investigación de los dinosaurios. La publicación de la primera y segunda ediciones de The Dinosauria en 1990 y 2004, y de un artículo de revisión de Paul Sereno en 1998, estuvieron acompañadas de un aumento en el número de árboles filogenéticos de dinosaurios publicados. [72]
Los fósiles de dinosaurios no se limitan a huesos, sino que también incluyen huellas o restos mineralizados de cubiertas de piel, órganos y otros tejidos. De estos, los revestimientos cutáneos basados en proteínas de queratina se conservan más fácilmente debido a su estructura molecular hidrófoba y reticulada . [73] Se conocen fósiles de cubiertas de piel a base de queratina o cubiertas de piel ósea de la mayoría de los grupos principales de dinosaurios. Desde el siglo XIX se han encontrado fósiles de dinosaurios con impresiones de piel escamosa. Samuel Beckles descubrió una extremidad anterior de saurópodo con piel conservada en 1852 que se atribuyó incorrectamente a un cocodrilo; Marsh lo atribuyó correctamente en 1888 y fue objeto de más estudios por parte de Reginald Hooley en 1917. [74] Entre los ornitisquios, en 1884 Jacob Wortman encontró impresiones de piel en el primer espécimen conocido de Edmontosaurus annectens , que fueron en gran parte destruidas durante la excavación del espécimen. [75] Owen y Hooley describieron posteriormente impresiones de la piel de Hypsilophodon e Iguanodon en 1885 y 1917. [74] Desde entonces, las impresiones de escamas se han encontrado con mayor frecuencia entre los hadrosáuridos, donde las impresiones se conocen de casi todo el cuerpo en múltiples especímenes. [76]
A partir de los años 90, importantes descubrimientos de fósiles excepcionalmente conservados en depósitos conocidos como Lagerstätten de conservación contribuyeron a la investigación sobre los tejidos blandos de los dinosaurios. [77] [78] Principalmente entre ellas estaban las rocas que produjeron las biotas Jehol (Cretácico temprano) y Yanliao (Jurásico medio a tardío) del noreste de China, de las cuales cientos de especímenes de dinosaurios con impresiones de estructuras parecidas a plumas (ambas estrechamente relacionados con las aves y otros, ver § Origen de las aves) han sido descritos por Xing Xu y sus colegas. [79] [80] En los reptiles y mamíferos vivos, las estructuras celulares que almacenan pigmentos conocidas como melanosomas son parcialmente responsables de producir la coloración. [81] [82] Se han informado rastros químicos de melanina y melanosomas de formas características en las plumas y escamas de los dinosaurios Jehol y Yanliao, incluidos terópodos y ornitisquios. [83] Esto ha permitido múltiples reconstrucciones de cuerpo completo de la coloración de los dinosaurios , como las de Sinosauropteryx [84] y Psittacosaurus [85] por Jakob Vinther y sus colegas, y técnicas similares también se han extendido a fósiles de dinosaurios de otras localidades. [81] (Sin embargo, algunos investigadores también han sugerido que los melanosomas fosilizados representan restos bacterianos. [86] [87] ) El contenido del estómago en algunos dinosaurios Jehol y Yanliao estrechamente relacionados con las aves también ha proporcionado indicaciones indirectas de la dieta y la anatomía del sistema digestivo (p. ej. , cultivos ). [88] [89] Se ha informado más evidencia concreta de la anatomía interna en Scipionyx de Pietraroja Plattenkalk de Italia. Preserva porciones de los intestinos, el colon, el hígado, los músculos y la tráquea. [90]
Al mismo tiempo, una línea de trabajo dirigida por Mary Higby Schweitzer , Jack Horner y sus colegas informaron varios casos de proteínas y tejidos blandos conservados dentro de fósiles de huesos de dinosaurios. Schweitzer y otros habían encontrado varias estructuras mineralizadas que probablemente representaban glóbulos rojos y fibras de colágeno en huesos de tiranosáuridos ya en 1991. [91] [92] [93] Sin embargo, en 2005, Schweitzer y sus colegas informaron que un fémur de Tyrannosaurus En su interior se conservaba tejido blando y flexible, incluidos vasos sanguíneos , matriz ósea y tejido conectivo (fibras óseas) que habían conservado su estructura microscópica. [94] Este descubrimiento sugirió que los tejidos blandos originales podrían conservarse a lo largo del tiempo geológico, [73] habiéndose propuesto múltiples mecanismos. [95] Más tarde, en 2009, Schweitzer y sus colegas informaron que un fémur de Brachylophosaurus conservaba microestructuras similares, y las técnicas inmunohistoquímicas (basadas en la unión de anticuerpos ) demostraron la presencia de proteínas como colágeno, elastina y laminina . [96] Ambos especímenes produjeron secuencias de proteínas de colágeno que eran viables para análisis filogenéticos moleculares , que los agruparon con aves como era de esperar. [96] [97] También se ha informado de la extracción de ADN fragmentario para ambos fósiles, [98] junto con un espécimen de Hypacrosaurus . [99] En 2015, Sergio Bertazzo y sus colegas informaron sobre la preservación de fibras de colágeno y glóbulos rojos en ocho especímenes de dinosaurios del Cretácico que no mostraron ningún signo de preservación excepcional, lo que indica que el tejido blando puede preservarse con más frecuencia de lo que se pensaba anteriormente. [100] Se han rechazado las sugerencias de que estas estructuras representan biopelículas bacterianas [101] , [102] pero la contaminación cruzada sigue siendo una posibilidad difícil de detectar. [103]
Los dinosaurios se separaron de sus ancestros arcosaurios durante las épocas del Triásico Medio y Tardío, aproximadamente 20 millones de años después de que la devastadora extinción del Pérmico-Triásico acabara con aproximadamente el 96% de todas las especies marinas y el 70% de las especies de vertebrados terrestres hace aproximadamente 252 millones de años. [104] [105] Los fósiles de dinosaurios más antiguos conocidos a partir de restos sustanciales datan de la época Carniana del período Triásico y se han encontrado principalmente en las Formaciones Ischigualasto y Santa María de Argentina y Brasil, y en la Formación Pebbly Arkose de Zimbabwe. [106]
La Formación Ischigualasto ( fechada radiométricamente entre 231 y 230 millones de años [107] ) ha producido el saurisquio temprano Eoraptor , originalmente considerado un miembro de Herrerasauridae [108] pero ahora considerado como un sauropodomorfo temprano, junto con los herrerasáuridos Herrerasaurus y Sanjuansaurus . y los sauropodomorfos Chromogisaurus , Eodromaeus y Panphagia . [109] El probable parecido de Eoraptor con el ancestro común de todos los dinosaurios sugiere que los primeros dinosaurios habrían sido depredadores pequeños y bípedos . [110] [111] [112] La Formación Santa María (fechada radiométricamente como más antigua, con 233,23 millones de años [113] ) ha producido los herrerasáuridos Gnathovorax y Staurikosaurus , junto con los sauropodomorfos Bagualosaurus , Buriolestes , Guaibasaurus , Macrocollum , Nhandumirim . , Pampadromaeus , Saturnalia y Unaysaurus . [109] La Formación Pebbly Arkose, que es de edad incierta pero probablemente era comparable a las otras dos, ha producido el sauropodomorfo Mbiresaurus , junto con un herrerasáurido sin nombre. [106]
Restos menos bien conservados de los sauropodomorfos Jaklapallisaurus y Nambalia , junto con el saurisquio temprano Alwalkeria , se conocen en las formaciones Upper Maleri y Lower Maleri de la India. [114] La Formación Chañares de Argentina, de edad Carniense, preserva ornitodiranos primitivos parecidos a dinosaurios, como Lagosuchus y Lagerpeton en Argentina , lo que la convierte en otro sitio importante para comprender la evolución de los dinosaurios. Estos ornitodiranos apoyan el modelo de los primeros dinosaurios como depredadores pequeños y bípedos. [109] [115] Los dinosaurios pueden haber aparecido ya en la época anisiana del Triásico, hace aproximadamente 243 millones de años, que es la edad del Nyasasaurus de la Formación Manda de Tanzania. Sin embargo, sus fósiles conocidos son demasiado fragmentarios para identificarlo como un dinosaurio o sólo como un pariente cercano. [116] La referencia de la Formación Manda al Anisiano también es incierta. Independientemente, los dinosaurios existieron junto con los ornitodiranos no dinosaurios durante un período de tiempo, con estimaciones que oscilan entre 5 y 10 millones de años [117] a 21 millones de años. [113]
Cuando aparecieron los dinosaurios, no eran los animales terrestres dominantes. Los hábitats terrestres fueron ocupados por varios tipos de arcosauromorfos y terápsidos , como cinodontos y rincosaurios . Sus principales competidores fueron los pseudosuquios , como los aetosaurios , los ornitosúquidos y los rauisuquios, que tuvieron más éxito que los dinosaurios. [118] La mayoría de estos otros animales se extinguieron en el Triásico, en uno de dos eventos. En primer lugar, hace unos 215 millones de años, se extinguieron una variedad de arcosauromorfos basales , incluidos los protorosaurios . A esto le siguió el evento de extinción Triásico-Jurásico (hace unos 201 millones de años), que supuso el fin de la mayoría de los otros grupos de arcosaurios primitivos, como los aetosaurios, los ornitosúquidos, los fitosaurios y los rauisuquios. Los rincosaurios y los dicinodontos sobrevivieron (al menos en algunas áreas) al menos en etapas tempranas del noriano medio y tardío o temprano rético , respectivamente, [119] [120] y la fecha exacta de su extinción es incierta. Estas pérdidas dejaron tras de sí una fauna terrestre de crocodilomorfos , dinosaurios, mamíferos, pterosaurios y tortugas . [10] Las primeras líneas de dinosaurios primitivos se diversificaron a lo largo de las etapas Carniense y Noriana del Triásico, posiblemente ocupando los nichos de los grupos que se extinguieron. [12] También es de destacar que hubo una mayor tasa de extinción durante el evento pluvial del Carniano . [121]
Evolución y paleobiogeografía.
El supercontinente Pangea a principios del Mesozoico (hace unos 200 millones de años)
La evolución de los dinosaurios después del Triásico siguió cambios en la vegetación y la ubicación de los continentes. En el Triásico Tardío y el Jurásico Temprano, los continentes estaban conectados como una única masa terrestre Pangea , y había una fauna de dinosaurios en todo el mundo compuesta principalmente por carnívoros celofisoides y herbívoros sauropodomorfos tempranos. [122] Las plantas gimnospermas (particularmente coníferas ), una fuente potencial de alimento, irradiaron en el Triásico Tardío. Los primeros sauropodomorfos no tenían mecanismos sofisticados para procesar los alimentos en la boca, por lo que debieron emplear otros medios para descomponer los alimentos a lo largo del tracto digestivo. [123] La homogeneidad general de las faunas de los dinosaurios continuó hasta el Jurásico medio y tardío, donde la mayoría de las localidades tenían depredadores formados por ceratosaurios , megalosauroides y alosauroides , y herbívoros formados por ornitisquios estegosaurios y grandes saurópodos. Ejemplos de esto incluyen la Formación Morrison de América del Norte y los lechos Tendaguru de Tanzania. Los dinosaurios en China muestran algunas diferencias, con terópodos metriacantosáuridos especializados y saurópodos inusuales de cuello largo como Mamenchisaurus . [122] Los anquilosaurios y los ornitópodos también se estaban volviendo más comunes, pero los sauropodomorfos primitivos se habían extinguido. Las coníferas y las pteridofitas eran las plantas más comunes. Los saurópodos, al igual que los sauropodomorfos anteriores, no eran procesadores orales, pero los ornitisquios estaban desarrollando diversos medios para manejar los alimentos en la boca, incluidos posibles órganos parecidos a mejillas para mantener los alimentos en la boca y movimientos de la mandíbula para moler los alimentos. [123] Otro evento evolutivo notable del Jurásico fue la aparición de aves verdaderas, descendientes de celurosaurios maniraptoranos. [14]
En el Cretácico Inferior y la actual desintegración de Pangea, los dinosaurios se diferenciaban fuertemente según la masa terrestre. La primera parte de esta época vio la expansión de anquilosaurios, iguanodontes y braquiosáuridos por Europa, América del Norte y el norte de África . Estos fueron posteriormente complementados o reemplazados en África por grandes terópodos espinosáuridos y carcarodontosáuridos , y saurópodos rebaquisáuridos y titanosaurios , que también se encuentran en América del Sur . En Asia , los celurosaurios maniraptoranos como los dromeosáuridos, troodóntidos y oviraptorosaurios se convirtieron en terópodos comunes, y los anquilosáuridos y los primeros ceratopsianos como Psittacosaurus se convirtieron en importantes herbívoros. Mientras tanto, Australia albergaba una fauna de anquilosaurios basales, hipsilofodontos e iguanodontes. [122] Los estegosaurios parecen haberse extinguido en algún momento a finales del Cretácico Inferior o principios del Cretácico Superior . Un cambio importante en el Cretácico Inferior, que se amplificaría en el Cretácico Superior, fue la evolución de las plantas con flores . Al mismo tiempo, varios grupos de dinosaurios herbívoros desarrollaron formas más sofisticadas de procesar los alimentos por vía oral. Los ceratopsianos desarrollaron un método de corte con dientes apilados unos sobre otros en baterías, y los iguanodoncianos perfeccionaron un método de trituración con baterías dentales , llevado al extremo en los hadrosáuridos. [123] Algunos saurópodos también desarrollaron baterías de dientes, mejor ejemplificadas por el rebaquisáurido Nigersaurus . [124]
Había tres faunas generales de dinosaurios en el Cretácico Superior. En los continentes del norte de América del Norte y Asia, los principales terópodos eran tiranosáuridos y varios tipos de terópodos maniraptoranos más pequeños, con un conjunto de herbívoros predominantemente ornitisquios de hadrosáuridos, ceratopsianos, anquilosáuridos y paquicefalosaurios. En los continentes del sur que formaban el ahora dividido supercontinente Gondwana , los abelisáuridos eran los terópodos comunes y los saurópodos titanosaurios los herbívoros comunes. Finalmente, en Europa prevalecieron los dromeosáuridos, los iguanodóntidos rabdodóntidos , los anquilosaurios nodosáuridos y los saurópodos titanosáuridos. [122] Las plantas con flores irradiaban mucho, [123] y los primeros pastos aparecieron al final del Cretácico. [125] Los hadrosáuridos trituradores y los ceratopsianos esquiladores se volvieron muy diversos en América del Norte y Asia. Los terópodos también se irradiaron como herbívoros u omnívoros , y los terizinosaurios y ornitomimosaurios se volvieron comunes. [123]
El evento de extinción Cretácico-Paleógeno, que ocurrió hace aproximadamente 66 millones de años al final del Cretácico, provocó la extinción de todos los grupos de dinosaurios excepto las aves neornitinas. Algunos otros grupos de diápsidos, incluidos los cocodrilos , los dirosaurios , los sebecosuquios , las tortugas, los lagartos , las serpientes , los esfenodontos y los coristoderanos , también sobrevivieron al evento. [126]
Los linajes supervivientes de aves neornitinas, incluidos los antepasados de las ratites , patos y gallinas modernos , y una variedad de aves acuáticas , se diversificaron rápidamente al comienzo del período Paleógeno , ingresando a nichos ecológicos que quedaron vacantes por la extinción de grupos de dinosaurios mesozoicos como los arbóreos. enantiornitinas , hesperornitinas acuáticas e incluso los terópodos terrestres más grandes (en forma de gastornis , eogruidos , batornitidos , ratites, geranoides , mihirungs y " pájaros del terror "). A menudo se afirma que los mamíferos superaron a los neornitinos por el dominio de la mayoría de los nichos terrestres, pero muchos de estos grupos coexistieron con ricas faunas de mamíferos durante la mayor parte de la Era Cenozoica . [127] Las aves terroríficas y los batornítidos ocuparon gremios carnívoros junto con los mamíferos depredadores, [128] [129] y las ratites todavía tienen bastante éxito como herbívoros de tamaño mediano; Los eogruidos existieron de manera similar desde el Eoceno hasta el Plioceno , y se extinguieron muy recientemente, después de más de 20 millones de años de coexistencia con muchos grupos de mamíferos. [130]
Clasificación
Los dinosaurios pertenecen a un grupo conocido como arcosaurios, al que también pertenecen los cocodrilos modernos. Dentro del grupo de los arcosaurios, los dinosaurios se diferencian más notablemente por su forma de andar. Las patas de los dinosaurios se extienden directamente debajo del cuerpo, mientras que las patas de los lagartos y los cocodrilos se extienden hacia ambos lados. [30]
Colectivamente, los dinosaurios como clado se dividen en dos ramas principales, Saurischia y Ornithischia. Saurischia incluye aquellos taxones que comparten un ancestro común más reciente con las aves que con Ornithischia, mientras que Ornithischia incluye todos los taxones que comparten un ancestro común más reciente con Triceratops que con Saurischia. Anatómicamente, estos dos grupos se pueden distinguir más notablemente por su estructura pélvica . Los primeros saurisquios, "cadera de lagarto", del griego sauros ( σαῦρος ), que significa "lagarto" e ischion ( ἰσχίον ), que significa "articulación de la cadera", conservaban la estructura de la cadera de sus antepasados, con un hueso púbico dirigido cranealmente o hacia adelante. [37] Esta forma básica se modificó girando el pubis hacia atrás en diversos grados en varios grupos ( Herrerasaurus , [131] terizinosáuridos, [132] dromeosáuridos, [133] y aves [14] ). Los saurisquios incluyen a los terópodos (exclusivamente bípedos y con una amplia variedad de dietas) y a los sauropodomorfos (herbívoros de cuello largo que incluyen grupos cuadrúpedos avanzados). [29] [134]
Por el contrario, los ornitisquios ("cadera de pájaro", del griego ornitheios (ὀρνίθειος), que significa "de un pájaro" e ischion (ἰσχίον), que significa "articulación de la cadera", tenían una pelvis que superficialmente se parecía a la pelvis de un pájaro: el hueso púbico estaba orientado caudalmente (apuntando hacia atrás). A diferencia de las aves, el pubis ornitisquio también solía tener un proceso adicional de orientación hacia adelante. Ornithischia incluye una variedad de especies que eran principalmente herbívoras.
A pesar de los términos "cadera de pájaro" (Ornithischia) y "cadera de lagarto" (Saurischia), las aves no forman parte de Ornithischia. En cambio, las aves pertenecen a Saurischia, los dinosaurios con "caderas de lagarto": las aves evolucionaron a partir de dinosaurios anteriores con "caderas de lagarto". [30]
Taxonomía
La siguiente es una clasificación simplificada de los grupos de dinosaurios basada en sus relaciones evolutivas, y las de los principales grupos de dinosaurios Theropoda, Sauropodomorpha y Ornithischia, compilada por Justin Tweet. [135] Se pueden encontrar más detalles y otras hipótesis de clasificación en artículos individuales.
† Ceratosauria (carnívoros generalmente con cuernos elaborados que existieron desde el período Jurásico al Cretácico, originalmente incluían Coelophysoidea)
Cronología de los principales grupos de dinosaurios según Holtz (2007).
Paleobiología
El conocimiento sobre los dinosaurios se deriva de una variedad de registros fósiles y no fósiles, incluidos huesos fosilizados, heces , huellas , gastrolitos , plumas , impresiones de piel, órganos internos y otros tejidos blandos . [90] [94] Muchos campos de estudio contribuyen a nuestra comprensión de los dinosaurios, incluida la física (especialmente la biomecánica ), la química , la biología y las ciencias de la Tierra (de las cuales la paleontología es una subdisciplina). [136] [137] Dos temas de particular interés y estudio han sido el tamaño y el comportamiento de los dinosaurios. [138]
Tamaño
Diagrama de escala que compara al humano promedio con los dinosaurios más largos conocidos en cinco clados principales :
La evidencia actual sugiere que el tamaño promedio de los dinosaurios varió a lo largo del Triásico, Jurásico Temprano, Jurásico Tardío y Cretácico. [111] Los dinosaurios terópodos depredadores, que ocuparon la mayoría de los nichos de carnívoros terrestres durante el Mesozoico, suelen caer en la categoría de 100 a 1 000 kg (220 a 2 200 lb) cuando se clasifican por peso estimado en categorías basadas en orden de magnitud , mientras que los dinosaurios recientes Los mamíferos carnívoros depredadores alcanzan su punto máximo en la categoría de 10 a 100 kg (22 a 220 lb). [139] La moda de las masas corporales de los dinosaurios mesozoicos es de entre 1 y 10 toneladas métricas (1,1 y 11,0 toneladas cortas). [140] Esto contrasta marcadamente con el tamaño promedio de los mamíferos cenozoicos, estimado por el Museo Nacional de Historia Natural en alrededor de 2 a 5 kg (4,4 a 11,0 libras). [141]
Los saurópodos eran los dinosaurios más grandes y pesados. Durante gran parte de la era de los dinosaurios, los saurópodos más pequeños eran más grandes que cualquier otra cosa en su hábitat, y el más grande era un orden de magnitud más masivo que cualquier otra cosa que haya caminado sobre la Tierra desde entonces. Los mamíferos prehistóricos gigantes como Paraceratherium (el mamífero terrestre más grande jamás visto) quedaron eclipsados por los saurópodos gigantes, y sólo las ballenas modernas se acercan o superan en tamaño. [142] Hay varias ventajas propuestas para el gran tamaño de los saurópodos, incluida la protección contra la depredación, la reducción del uso de energía y la longevidad, pero puede ser que la ventaja más importante fuera la dietética. Los animales grandes son más eficientes en la digestión que los animales pequeños, porque la comida pasa más tiempo en sus sistemas digestivos. Esto también les permite subsistir con alimentos con menor valor nutritivo que los animales más pequeños. Los restos de saurópodos se encuentran principalmente en formaciones rocosas interpretadas como secas o estacionalmente secas, y la capacidad de comer grandes cantidades de ramoneo bajo en nutrientes habría sido ventajosa en tales ambientes. [143]
más grande y más pequeño
Probablemente los científicos nunca estarán seguros de cuáles son los dinosaurios más grandes y más pequeños que jamás hayan existido. Esto se debe a que sólo un pequeño porcentaje de animales fueron fosilizados y la mayoría de ellos permanecen enterrados en la tierra. Pocos de los especímenes recuperados son esqueletos completos y las impresiones de piel y otros tejidos blandos son raras. Reconstruir un esqueleto completo comparando el tamaño y la morfología de los huesos con los de especies similares y más conocidas es un arte inexacto, y reconstruir los músculos y otros órganos del animal vivo es, en el mejor de los casos, un proceso de conjeturas fundamentadas. [144]
Tamaño comparativo del Argentinosaurus con el del humano promedio
El dinosaurio más alto y pesado conocido a partir de buenos esqueletos es Giraffatitan brancai (anteriormente clasificado como una especie de Brachiosaurus ). Sus restos fueron descubiertos en Tanzania entre 1907 y 1912. Se incorporaron huesos de varios individuos de tamaño similar al esqueleto ahora montado y expuesto en el Museo für Naturkunde de Berlín ; [145] esta montura mide 12 metros (39 pies) de alto y 21,8 a 22,5 metros (72 a 74 pies) de largo, [146] [147] y habría pertenecido a un animal que pesaba entre 30 000 y 60 000 kilogramos ( 70 000 y 130 000 libras). El dinosaurio completo más largo es el Diplodocus de 27 metros (89 pies) de largo , que fue descubierto en Wyoming en los Estados Unidos y exhibido en el Museo Carnegie de Historia Natural de Pittsburgh en 1907. [148] El dinosaurio más largo conocido a partir de buen material fósil es Patagotitan : el esqueleto que se encuentra en el Museo Americano de Historia Natural de Nueva York mide 37 metros (121 pies) de largo. El Museo Municipal Carmen Funes en Plaza Huincul , Argentina, tiene un esqueleto de Argentinosaurus reconstruido que mide 39,7 metros (130 pies) de largo. [149]
Maraapunisaurus , uno de los animales más grandes que ha caminado sobre la tierra.Bruhathkayosaurus , potencialmente el animal terrestre más grande que jamás haya existido.
Existieron dinosaurios más grandes, pero su conocimiento se basa enteramente en un pequeño número de fósiles fragmentarios. La mayoría de los especímenes herbívoros más grandes registrados fueron descubiertos en la década de 1970 o después, e incluyen al enorme Argentinosaurus , que pudo haber pesado entre 80.000 y 100.000 kilogramos (88 a 110 toneladas cortas) y alcanzar longitudes de 30 a 40 metros (98 a 110 toneladas). 131 pies); algunos de los más largos fueron el Diplodocus hallorum [143] (antes Seismosaurus ), de 33,5 metros (110 pies) de largo, el Supersaurus de 33 a 34 metros (108 a 112 pies) de largo , [150] y el de 37 metros (121 ft) largo Patagotitan ; y el más alto, el Sauroposeidon de 18 metros (59 pies) , que podría haber llegado a una ventana del sexto piso. Hubo algunos dinosaurios que se consideraban los más pesados y los más largos. El más famoso incluye Amphicoelias fragillimus , conocido sólo por un arco neural vertebral parcial ahora perdido descrito en 1878. Extrapolando a partir de la ilustración de este hueso, el animal pudo haber medido 58 metros (190 pies) de largo y pesado 122 400 kg ( 269 800 libras). [143] Sin embargo, investigaciones recientes han colocado a Amphicoelias desde el largo y grácil diplodócido hasta el más corto pero mucho más robusto rebaquisáurido. Ahora rebautizado como Maraapunisaurus , este saurópodo mide hasta 40 metros (130 pies) de largo y pesa hasta 120 000 kg ( 260 000 lb). [151] [152] Otro contendiente por este título incluye Bruhathkayosaurus matleyi , un taxón increíblemente controvertido cuya existencia se confirmó recientemente después de que se descubrieron fotografías archivadas. [153] Bruhathkayosaurus era un titanosaurio y probablemente habría pesado más que incluso Marrapunisaurus . Estimaciones de tamaño recientes en 2023 han colocado a este saurópodo alcanzando longitudes de hasta 44 m (144 pies) de largo y un rango de peso colosal de alrededor de 110 000 – 170 000 kg ( 240 000 – 370 000 lb), si estas estimaciones superiores son ciertas, Bruhathkayosaurus habría rivalizado con la ballena azul y el coloso Perucetus como uno de los animales más grandes que jamás haya existido. [154]
El dinosaurio carnívoro más grande fue el Spinosaurus , que alcanzaba una longitud de 12,6 a 18 metros (41 a 59 pies) y un peso de 7 a 20,9 toneladas métricas (7,7 a 23,0 toneladas cortas). [155] [156] Otros terópodos carnívoros grandes incluyeron Giganotosaurus , Carcharodontosaurus y Tyrannosaurus . [156] Therizinosaurus y Deinocheirus estaban entre los terópodos más altos. El dinosaurio ornitisquio más grande fue probablemente el hadrosáurido Shantungosaurus giganteus , que medía 16,6 metros (54 pies). [157] Los individuos más grandes pueden haber pesado hasta 16 toneladas métricas (18 toneladas cortas). [158]
El dinosaurio más pequeño conocido es el colibrí abeja , [159] con una longitud de sólo 5 centímetros (2,0 pulgadas) y una masa de alrededor de 1,8 g (0,063 oz). [160] Los dinosaurios no aviales más pequeños conocidos eran aproximadamente del tamaño de palomas y eran los terópodos más estrechamente relacionados con las aves. [161] Por ejemplo, Anchiornis huxleyi es actualmente el dinosaurio no aviano más pequeño descrito a partir de un espécimen adulto, con un peso estimado de 110 g (3,9 oz) [162] y una longitud esquelética total de 34 centímetros (1,12 pies). [161] [162] Los dinosaurios herbívoros no aviares más pequeños incluían a Microceratus y Wannanosaurus , de aproximadamente 60 centímetros (2,0 pies) de largo cada uno. [163] [164]
Muchas aves modernas son muy sociables y a menudo se las encuentra viviendo en bandadas. Existe un acuerdo general en que algunos comportamientos que son comunes en las aves, así como en los cocodrilos (parientes vivos más cercanos de las aves), también eran comunes entre los grupos de dinosaurios extintos. Las interpretaciones del comportamiento de las especies fósiles se basan generalmente en la pose de los esqueletos y su hábitat , simulaciones por computadora de su biomecánica y comparaciones con animales modernos en nichos ecológicos similares. [136]
La primera evidencia potencial de pastoreo o bandadas como un comportamiento generalizado común a muchos grupos de dinosaurios además de las aves fue el descubrimiento en 1878 de 31 Iguanodon , ornitisquios que entonces se pensaba que habían perecido juntos en Bernissart , Bélgica , después de caer en un profundo, Dolina inundada y ahogada. [165] Posteriormente se descubrieron otros lugares de muerte masiva. Estos, junto con múltiples huellas, sugieren que el comportamiento gregario era común en muchas especies de dinosaurios primitivos. Las huellas de cientos o incluso miles de herbívoros indican que los hadrosáuridos pueden haberse movido en grandes manadas, como el bisonte americano o la gacela africana . Las huellas de saurópodos documentan que estos animales viajaban en grupos compuestos de varias especies diferentes, al menos en Oxfordshire , Inglaterra, [166] aunque no hay evidencia de estructuras de rebaño específicas. [167] La congregación en rebaños puede haber evolucionado con fines de defensa, con fines migratorios o para brindar protección a las crías. Hay evidencia de que muchos tipos de dinosaurios de crecimiento lento, incluidos varios terópodos, saurópodos, anquilosaurios, ornitópodos y ceratopsianos, formaron agregaciones de individuos inmaduros. Un ejemplo es un sitio en Mongolia Interior que ha arrojado restos de más de 20 Sinornithomimus , de uno a siete años de edad. Este ensamblaje se interpreta como un grupo social que quedó atrapado en el barro. [168] La interpretación de los dinosaurios como gregarios también se ha extendido a representar a los terópodos carnívoros como cazadores en manada que trabajan juntos para derribar presas grandes. [169] [170] Sin embargo, este estilo de vida es poco común entre las aves, los cocodrilos y otros reptiles modernos, y la evidencia tafonómica que sugiere la caza en manada similar a la de los mamíferos en terópodos como Deinonychus y Allosaurus también puede interpretarse como el resultado de disputas fatales entre alimentar a los animales, como se ve en muchos depredadores diápsidos modernos. [171]
Desde un punto de vista conductual, uno de los fósiles de dinosaurios más valiosos fue descubierto en el desierto de Gobi en 1971. Incluía un Velociraptor atacando a un Protoceratops , [173] proporcionando evidencia de que los dinosaurios efectivamente se atacaban entre sí. [174] Evidencia adicional de atacar presas vivas es la cola parcialmente curada de un Edmontosaurus , un dinosaurio hadrosáurido; la cola está dañada de tal manera que muestra que el animal fue mordido por un tiranosaurio pero sobrevivió. [174] El canibalismo entre algunas especies de dinosaurios fue confirmado por marcas de dientes encontradas en Madagascar en 2003, que involucraban al terópodo Majungasaurus . [175]
Se han utilizado comparaciones entre los anillos esclerales de los dinosaurios y las aves y reptiles modernos para inferir los patrones de actividad diaria de los dinosaurios. Aunque se ha sugerido que la mayoría de los dinosaurios estaban activos durante el día, estas comparaciones han demostrado que los pequeños dinosaurios depredadores como los dromeosáuridos, Juravenator y Megapnosaurus probablemente eran nocturnos . Los dinosaurios herbívoros y omnívoros de tamaño grande y mediano, como los ceratopsianos, sauropodomorfos, hadrosáuridos y ornitomimosaurios, pueden haber sido catémerales , activos durante intervalos cortos a lo largo del día, aunque se infirió que el pequeño ornitisquio Agilisaurus era diurno . [176]
Según la evidencia fósil de dinosaurios como Oryctodromeus , algunas especies de ornitisquios parecen haber llevado un estilo de vida parcialmente fosorial (excavador). [177] Muchas aves modernas son arbóreas (trepan árboles), y esto también era cierto para muchas aves mesozoicas, especialmente las enantiornitinas. [178] Si bien algunas especies primitivas parecidas a aves pueden haber sido también arbóreas (incluidos los dromeosáuridos) como Microraptor [179] ), la mayoría de los dinosaurios no aviales parecen haber dependido de la locomoción terrestre. Una buena comprensión de cómo se movían los dinosaurios sobre la Tierra es clave para los modelos de comportamiento de los dinosaurios; La ciencia de la biomecánica, iniciada por Robert McNeill Alexander , ha proporcionado conocimientos importantes en esta área. Por ejemplo, los estudios de las fuerzas ejercidas por los músculos y la gravedad sobre la estructura esquelética de los dinosaurios han investigado qué tan rápido podían correr los dinosaurios, [136] si los diplodócidos podían crear explosiones sónicas mediante el chasquido de la cola en forma de látigo , [180] y si los saurópodos podían flotar. [181]
Comunicación
Se sabe que las aves modernas se comunican mediante señales visuales y auditivas, y la amplia diversidad de estructuras de visualización visual entre los grupos de dinosaurios fósiles, como cuernos, volantes, crestas, velas y plumas, sugiere que la comunicación visual siempre ha sido importante en la biología de los dinosaurios. [182] La reconstrucción del color del plumaje de Anchiornis sugiere la importancia del color en la comunicación visual en dinosaurios no aviares. [183] La vocalización en dinosaurios no aviares es menos segura. En las aves, la laringe no desempeña ningún papel en la producción de sonidos. En lugar de eso, vocalizan con un órgano novedoso llamado siringe , ubicado más abajo en la tráquea. [184] Los restos más antiguos de una siringe se encontraron en un espécimen de Vegavis iaai , parecido a un pato, que data de hace 69 a 66 millones de años, y es poco probable que este órgano haya existido en dinosaurios no aviares. [185]
Restauración de una exhibición visual llamativa e inusual en un Lambeosaurus magnicristatus . La cresta también podría haber actuado como cámara de resonancia de sonidos.
El paleontólogo Phil Senter ha sugerido que dado que los dinosaurios no aviares no tenían siringe, y sus parientes vivos más cercanos, los cocodrilos, usaban la laringe, no podían vocalizar ya que el ancestro común habría sido mudo. Afirma que principalmente en exhibiciones visuales y posiblemente en sonidos acústicos no vocales como silbidos, rechinar la mandíbula o aplaudir, chapotear y batir de alas (posible en los dinosaurios maniraptoranos alados). [182] Otros investigadores han respondido que las vocalizaciones también existen en las tortugas, los parientes más cercanos de los arcosaurios, lo que sugiere que el rasgo es ancestral de su linaje. Además, la comunicación vocal en los dinosaurios está indicada por el desarrollo de una audición avanzada en casi todos los grupos principales. Por lo tanto, la siringe pudo haber complementado y luego reemplazado a la laringe como órgano vocal en lugar de haber habido un "período de silencio" en la evolución de las aves. [186]
En 2023, se describió una laringe fosilizada de un espécimen del anquilosáurido Pinacosaurus . La estructura estaba compuesta por cartílagos cricoides y aritenoides , similares a los de los reptiles no aviares. Sin embargo, la articulación cricoides-aritenoidea móvil y los cartílagos aritenoides largos habrían permitido un control del flujo de aire similar al de las aves y, por lo tanto, podrían haber emitido vocalizaciones similares a las de las aves. Además, los cartílagos estaban osificados , lo que implica que la osificación laríngea es una característica de algunos dinosaurios no aviares. [187] Un estudio de 2016 concluye que algunos dinosaurios pueden haber producido vocalizaciones con la boca cerrada como arrullos, ululares y retumbos. Estos ocurren tanto en reptiles como en aves e implican inflar el esófago o las bolsas traqueales. Tales vocalizaciones evolucionaron de forma independiente en los arcosaurios existentes en numerosas ocasiones, tras aumentar el tamaño corporal. [188] Se ha sugerido que las crestas de algunos hadrosáuridos y las cámaras nasales de los anquilosáuridos han funcionado en resonancia acústica . [189] [190]
Todos los dinosaurios pusieron huevos amnióticos . Los huevos de dinosaurio generalmente se ponían en un nido. La mayoría de las especies crean nidos algo elaborados que pueden ser tazas, cúpulas, platos, lechos, montículos o madrigueras. [191] Algunas especies de aves modernas no tienen nidos; el arao común que anida en los acantilados pone sus huevos sobre rocas desnudas, y los pingüinos emperador machos mantienen los huevos entre el cuerpo y las patas. Las aves primitivas y muchos dinosaurios no aviales a menudo ponen huevos en nidos comunales, y los machos son principalmente los que incuban los huevos. Mientras que las aves modernas sólo tienen un oviducto funcional y ponen un huevo a la vez, las aves y los dinosaurios más primitivos tenían dos oviductos, como los cocodrilos. Algunos dinosaurios no aviales, como Troodon , exhibían una puesta iterativa, donde el adulto podía poner un par de huevos cada uno o dos días, y luego aseguraba la eclosión simultánea retrasando la crianza hasta que se hubieran puesto todos los huevos. [192]
Al poner huevos, a las hembras les crece un tipo especial de hueso entre el hueso exterior duro y la médula de sus extremidades. Este hueso medular, rico en calcio , se utiliza para fabricar cáscaras de huevo. Un descubrimiento de características en el esqueleto de un tiranosaurio proporcionó evidencia de hueso medular en dinosaurios extintos y, por primera vez, permitió a los paleontólogos establecer el sexo de un espécimen de dinosaurio fósil. Investigaciones adicionales han encontrado hueso medular en el carnosaurio Allosaurus y el ornitópodo Tenontosaurus . Debido a que la línea de dinosaurios que incluye Allosaurus y Tyrannosaurus divergió de la línea que condujo a Tenontosaurus muy temprano en la evolución de los dinosaurios, esto sugiere que la producción de tejido medular es una característica general de todos los dinosaurios. [193]
Otro rasgo muy extendido entre las aves modernas (pero véase más adelante en lo que respecta a los grupos fósiles y los megapodos existentes ) es el cuidado parental de las crías después de la eclosión. El descubrimiento de Jack Horner en 1978 de un lugar de anidación de Maiasaura ("lagarto buena madre") en Montana demostró que el cuidado de los padres continuaba mucho después del nacimiento entre los ornitópodos. [194] Un espécimen del oviraptórido Citipati osmolskae fue descubierto en una posición melancólica similar a la de un pollo en 1993, [195] lo que puede indicar que habían comenzado a usar una capa aislante de plumas para mantener los huevos calientes. [196] Se encontró un embrión del sauropodomorfo basal Massospondylus sin dientes, lo que indica que se requería cierto cuidado de los padres para alimentar a los dinosaurios jóvenes. [197] Los rastros también han confirmado el comportamiento parental entre los ornitópodos de la isla de Skye en el noroeste de Escocia . [198]
Sin embargo, existe amplia evidencia de precocialidad o superprecocialidad entre muchas especies de dinosaurios, particularmente los terópodos. Por ejemplo, se ha demostrado abundantemente que las aves no ornituromorfas tenían tasas de crecimiento lentas, un comportamiento de enterramiento de huevos similar al de los megapodos y la capacidad de volar poco después del nacimiento. [199] [200] [201] [202] Tanto Tyrannosaurus como Troodon tenían juveniles con clara superprecocialidad y probablemente ocupaban nichos ecológicos diferentes a los de los adultos. [192] Se ha inferido la superprecocialidad de los saurópodos. [203]
Es poco probable que las estructuras genitales se fosilicen ya que carecen de escamas que puedan permitir la conservación mediante pigmentación o sales residuales de fosfato de calcio. En 2021, se describió el espécimen mejor conservado del exterior de la cloaca de un dinosaurio para Psittacosaurus , que demuestra hinchazones laterales similares a las glándulas almizcleras de los cocodrilos utilizadas en exhibiciones sociales por ambos sexos y regiones pigmentadas que también podrían reflejar una función de señalización. Sin embargo, este ejemplar por sí solo no ofrece suficiente información para determinar si este dinosaurio tenía funciones de señalización sexual; sólo apoya la posibilidad. La señalización visual cloacal puede ocurrir tanto en machos como en hembras en aves vivas, por lo que es poco probable que sea útil para determinar el sexo de los dinosaurios extintos. [204]
Fisiología
Debido a que tanto los cocodrilos como las aves modernas tienen corazones de cuatro cámaras (aunque modificados en los cocodrilos), es probable que este sea un rasgo compartido por todos los arcosaurios, incluidos todos los dinosaurios. [205] Si bien todas las aves modernas tienen metabolismos elevados y son endotérmicas ("de sangre caliente"), desde la década de 1960 ha habido un vigoroso debate sobre hasta qué punto se remontaba este rasgo en el linaje de los dinosaurios. Varios investigadores han sostenido que los dinosaurios eran endotérmicos, ectotérmicos ("de sangre fría") o algo intermedio. [206] Un consenso emergente entre los investigadores es que, si bien diferentes linajes de dinosaurios habrían tenido metabolismos diferentes, la mayoría de ellos tenían tasas metabólicas más altas que otros reptiles pero más bajas que las aves y mamíferos vivos, [207] lo que algunos denominan mesotermia . [208] La evidencia de los cocodrilos y sus parientes extintos sugiere que metabolismos tan elevados podrían haberse desarrollado en los primeros arcosaurios, que eran los ancestros comunes de los dinosaurios y los cocodrilos. [209] [210]
Esta restauración de 1897 del Brontosaurio como un animal acuático que arrastra la cola, realizada por Charles R. Knight , tipifica las primeras opiniones sobre los estilos de vida de los dinosaurios.
Después de que se descubrieron los dinosaurios no aviares, los paleontólogos postularon por primera vez que eran ectotérmicos. Esto se utilizó para dar a entender que los dinosaurios antiguos eran organismos relativamente lentos y perezosos, a pesar de que muchos reptiles modernos son rápidos y ligeros a pesar de depender de fuentes externas de calor para regular su temperatura corporal. La idea de que los dinosaurios eran ectotérmicos siguió siendo una visión predominante hasta que Robert T. Bakker , uno de los primeros defensores de la endotermia de los dinosaurios, publicó un influyente artículo sobre el tema en 1968. Bakker utilizó específicamente evidencia anatómica y ecológica para argumentar que los saurópodos, que hasta entonces habían sido representados. como animales acuáticos extendidos con la cola arrastrando por el suelo, eran endotermos que vivían vidas terrestres vigorosas. En 1972, Bakker amplió sus argumentos basados en los requisitos energéticos y la relación depredador-presa. Este fue uno de los resultados fundamentales que condujeron al renacimiento de los dinosaurios. [63] [64] [60] [211]
Una de las mayores contribuciones a la comprensión moderna de la fisiología de los dinosaurios ha sido la paleohistología , el estudio de la estructura microscópica de los tejidos de los dinosaurios. [212] [213] A partir de la década de 1960, Armand de Ricqlès sugirió que la presencia de hueso fibrolamelar (tejido óseo con una textura fibrosa irregular y lleno de vasos sanguíneos) era indicativa de un crecimiento consistentemente rápido y, por lo tanto, de endotermia. El hueso fibrolamelar era común tanto en dinosaurios como en pterosaurios, [214] [215] aunque no estaba presente universalmente. [216] [217] Esto ha llevado a una importante cantidad de trabajo en la reconstrucción de curvas de crecimiento y modelado de la evolución de las tasas de crecimiento en varios linajes de dinosaurios, [218] lo que ha sugerido en general que los dinosaurios crecieron más rápido que los reptiles vivos. [213] Otras líneas de evidencia que sugieren endotermia incluyen la presencia de plumas y otros tipos de cubiertas corporales en muchos linajes (ver § Plumas); proporciones más consistentes del isótopo oxígeno-18 en el tejido óseo en comparación con los ectotermos, particularmente a medida que variaba la latitud y, por lo tanto, la temperatura del aire, lo que sugiere temperaturas internas estables [219] [220] (aunque estas proporciones pueden alterarse durante la fosilización [221] ); y el descubrimiento de los dinosaurios polares , que vivieron en Australia, la Antártida y Alaska cuando estos lugares habrían tenido climas fríos y templados. [222] [223] [224] [225]
En los dinosaurios saurisquios, los metabolismos superiores estuvieron respaldados por la evolución del sistema respiratorio aviar, caracterizado por un extenso sistema de sacos de aire que extendieron los pulmones e invadieron muchos de los huesos del esqueleto, haciéndolos huecos. [226] Dichos sistemas respiratorios, que pueden haber aparecido en los primeros saurisquios, [227] les habrían proporcionado más oxígeno en comparación con un mamífero de tamaño similar, al mismo tiempo que tenían un volumen corriente en reposo mayor y requerían una frecuencia respiratoria más baja, lo que les habría permitido mantener mayores niveles de actividad. [142] El rápido flujo de aire también habría sido un mecanismo de enfriamiento eficaz, que junto con una tasa metabólica más baja [228] habría evitado que los saurópodos grandes se sobrecalentaran. Estos rasgos pueden haber permitido a los saurópodos crecer rápidamente hasta alcanzar tamaños gigantescos. [229] [230] Los saurópodos también pueden haberse beneficiado de su tamaño: su pequeña relación entre superficie y volumen significaba que habrían podido regular la termorregulación más fácilmente, un fenómeno denominado gigantotermia . [142] [231]
Al igual que otros reptiles, los dinosaurios son principalmente uricotélicos , es decir, sus riñones extraen desechos nitrogenados del torrente sanguíneo y los excretan como ácido úrico en lugar de urea o amoníaco a través de los uréteres hacia el intestino. Esto les habría ayudado a conservar el agua. [207] En la mayoría de las especies vivas, el ácido úrico se excreta junto con las heces como un desecho semisólido. [232] [233] Sin embargo, al menos algunas aves modernas (como los colibríes ) pueden ser facultativamente amonotélicas y excretan la mayoría de los desechos nitrogenados como amoníaco. [234] Este material, así como la salida de los intestinos, emerge de la cloaca . [235] [236] Además, muchas especies regurgitan bolitas , [237] y las bolitas fósiles se conocen ya en el Jurásico de Anchiornis . [238]
El tamaño y la forma del cerebro se pueden reconstruir parcialmente basándose en los huesos circundantes. En 1896, Marsh calculó relaciones entre el peso del cerebro y el peso corporal de siete especies de dinosaurios, mostrando que el cerebro de los dinosaurios era proporcionalmente más pequeño que el de los cocodrilos actuales, y que el cerebro del Stegosaurus era más pequeño que el de cualquier vertebrado terrestre vivo. Esto contribuyó a la noción pública generalizada de que los dinosaurios eran lentos y extraordinariamente estúpidos. Harry Jerison, en 1973, demostró que se esperan cerebros proporcionalmente más pequeños en cuerpos de mayor tamaño, y que el tamaño del cerebro en los dinosaurios no era menor de lo esperado en comparación con el de los reptiles vivos. [239] Investigaciones posteriores demostraron que el tamaño relativo del cerebro aumentó progresivamente durante la evolución de los terópodos, con la inteligencia más alta, comparable a la de las aves modernas, calculada para el troodóntido Troodon . [240]
origen de las aves
La posibilidad de que los dinosaurios fueran los antepasados de las aves fue sugerida por primera vez en 1868 por Thomas Henry Huxley . [241] Después del trabajo de Gerhard Heilmann a principios del siglo XX, la teoría de las aves como descendientes de dinosaurios fue abandonada en favor de la idea de que eran descendientes de tecodontos generalizados , siendo la evidencia clave la supuesta falta de clavículas en dinosaurios. [242] Sin embargo, como lo demostraron descubrimientos posteriores, las clavículas (o una única espoleta fusionada , que derivaba de clavículas separadas) en realidad no estaban ausentes; [14] habían sido encontrados ya en 1924 en Oviraptor , pero identificados erróneamente como una interclavícula . [243] En la década de 1970, Ostrom revivió la teoría dinosaurio-pájaro, [244] que ganó impulso en las próximas décadas con la llegada del análisis cladístico, [245] y un gran aumento en el descubrimiento de pequeños terópodos y pájaros primitivos. [32] De particular interés han sido los fósiles de Jehol Biota, donde se ha encontrado una variedad de terópodos y pájaros primitivos, a menudo con plumas de algún tipo. [70] [14] Las aves comparten más de cien características anatómicas distintas con los dinosaurios terópodos, que ahora se acepta generalmente que fueron sus parientes antiguos más cercanos. [246] Están más estrechamente aliados con los celurosaurios maniraptoranos. [14] Una minoría de científicos, en particular Alan Feduccia y Larry Martin , han propuesto otros caminos evolutivos, incluidas versiones revisadas de la propuesta del arcosaurio basal de Heilmann, [247] o que los terópodos maniraptoranos son los ancestros de las aves pero que ellos mismos no son dinosaurios, sólo convergente con los dinosaurios. [248]
Las plumas son una de las características más reconocibles de las aves modernas y un rasgo que también compartían varios dinosaurios no aviares. Según la distribución actual de la evidencia fósil, parece que las plumas eran un rasgo ancestral de los dinosaurios, aunque puede haberse perdido selectivamente en algunas especies. [249] Se ha descubierto evidencia fósil directa de plumas o estructuras similares a plumas en una amplia gama de especies en muchos grupos de dinosaurios no aviares, [70] tanto entre saurisquios como entre ornitisquios. Se conocen estructuras simples, ramificadas y parecidas a plumas de heterodontosáuridos , neornitisquios primitivos , [250] y terópodos, [251] y ceratopsianos primitivos. Sólo se ha encontrado evidencia de plumas verdaderas con vetas similares a las plumas de vuelo de las aves modernas en el subgrupo de terópodos Maniraptora, que incluye oviraptorosaurios, troodóntidos, dromeosáuridos y aves. [14] [252] También se han encontrado en pterosaurios estructuras parecidas a plumas conocidas como picnofibras . [253]
Sin embargo, los investigadores no se ponen de acuerdo sobre si estas estructuras comparten un origen común entre linajes (es decir, son homólogas ), [254] [255] o si fueron el resultado de una experimentación generalizada con cubiertas de piel entre ornitodiranos. [256] Si el primero es el caso, los filamentos pueden haber sido comunes en el linaje de los ornitodiranos y haber evolucionado antes de la aparición de los dinosaurios. [249] La investigación sobre la genética de los caimanes americanos ha revelado que los escudos de los cocodrilos poseen queratinas de plumas durante el desarrollo embrionario, pero los animales no expresan estas queratinas antes de la eclosión. [257] La descripción de los dinosaurios emplumados no ha estado exenta de controversia en general; Quizás los críticos más acérrimos hayan sido Alan Feduccia y Theagarten Lingham-Soliar, quienes han propuesto que algunos supuestos fósiles con forma de plumas son el resultado de la descomposición de la fibra colágena que subyacía a la piel de los dinosaurios, [ 258] [259] [260] y que los dinosaurios maniraptoranos con plumas avellanadas no eran en realidad dinosaurios, sino convergentes con los dinosaurios. [248] [259] Sin embargo, sus puntos de vista en su mayor parte no han sido aceptados por otros investigadores, hasta el punto de que la naturaleza científica de las propuestas de Feduccia ha sido cuestionada. [261]
Archaeopteryx fue el primer fósil encontrado que reveló una posible conexión entre los dinosaurios y las aves. Se considera un fósil de transición , ya que muestra características de ambos grupos. Salido a la luz sólo dos años despuésdel influyente libro de Charles Darwin Sobre el origen de las especies (1859), su descubrimiento estimuló el incipiente debate entre los defensores de la biología evolutiva y el creacionismo . Este madrugador se parece tanto a un dinosaurio que, al no tener una impresión clara de las plumas en la roca circundante, al menos un ejemplar fue confundido con el pequeño terópodo Compsognathus . [262] Desde la década de 1990, se han encontrado varios dinosaurios emplumados adicionales, lo que proporciona evidencia aún más sólida de la estrecha relación entre los dinosaurios y las aves modernas. Muchos de estos ejemplares fueron desenterrados en los lagerstätten de Jehol Biota. [255] Si las estructuras parecidas a plumas realmente estaban ampliamente presentes entre los dinosaurios no aviares, la falta de evidencia fósil abundante de ellas puede deberse al hecho de que las características delicadas como la piel y las plumas rara vez se conservan mediante fosilización y, por lo tanto, a menudo están ausentes del registro fósil. [263]
Esqueleto
Debido a que las plumas a menudo se asocian con las aves, los dinosaurios emplumados a menudo se promocionan como el eslabón perdido entre las aves y los dinosaurios. Sin embargo, las múltiples características esqueléticas que también comparten los dos grupos representan otra línea importante de evidencia para los paleontólogos. Las áreas del esqueleto con similitudes importantes incluyen el cuello, el pubis, la muñeca ( carpiano semilunar ), el brazo y la cintura pectoral , la fúrcula (horquilla) y el esternón . La comparación de esqueletos de aves y dinosaurios mediante análisis cladístico refuerza los argumentos a favor del vínculo. [264]
Los grandes dinosaurios carnívoros tenían un complejo sistema de sacos aéreos similar a los que se encuentran en las aves modernas, según una investigación de 2005 dirigida por Patrick M. O'Connor. Los pulmones de los dinosaurios terópodos (carnívoros que caminaban sobre dos patas y tenían patas parecidas a las de los pájaros) probablemente bombeaban aire hacia sacos huecos en sus esqueletos, como es el caso de las aves. "Lo que alguna vez se consideró formalmente exclusivo de las aves estaba presente de alguna forma en los antepasados de las aves", dijo O'Connor. [265] [266] En 2008, los científicos describieron Aerosteon riocoloradensis , cuyo esqueleto proporciona la evidencia más sólida hasta la fecha de un dinosaurio con un sistema respiratorio similar al de un pájaro. La tomografía computarizada de los huesos fósiles de Aerosteon reveló evidencia de la existencia de sacos de aire dentro de la cavidad corporal del animal. [226] [267]
Evidencia conductual
Los fósiles de los troodontos Mei y Sinornithoides demuestran que algunos dinosaurios dormían con la cabeza bajo el brazo. [268] Este comportamiento, que puede haber ayudado a mantener la cabeza caliente, también es característico de las aves modernas. También se han encontrado varios especímenes de deinonicosaurio y oviraptorosaurio conservados encima de sus nidos, probablemente meditando como si fueran pájaros. [269] La relación entre el volumen de huevos y la masa corporal de los adultos entre estos dinosaurios sugiere que los huevos fueron incubados principalmente por el macho, y que las crías eran muy precoces, similares a muchas aves terrestres modernas. [270]
Se sabe que algunos dinosaurios usaban piedras de molleja como las aves modernas. Los animales tragan estos cálculos para ayudar a la digestión y descomponer los alimentos y las fibras duras una vez que ingresan al estómago. Cuando se encuentran asociados con fósiles, los cálculos de molleja se denominan gastrolitos. [271]
Extinción de grupos principales.
Todos los dinosaurios no aviares y la mayoría de los linajes de aves [272] se extinguieron en un evento de extinción masiva , llamado evento de extinción Cretácico-Paleógeno (K-Pg) , al final del período Cretácico. Por encima del límite Cretácico-Paleógeno , que data de hace 66,038 ± 0,025 millones de años, [273] los fósiles de dinosaurios no aviares desaparecen abruptamente; La ausencia de fósiles de dinosaurios se utilizó históricamente para asignar rocas al Cenozoico posterior. La naturaleza del evento que causó esta extinción masiva ha sido ampliamente estudiada desde la década de 1970, lo que llevó al desarrollo de dos mecanismos que se cree que desempeñaron papeles importantes: un evento de impacto extraterrestre en la Península de Yucatán , junto con un vulcanismo de inundación de basalto en la India. . Sin embargo, los mecanismos específicos del evento de extinción y el alcance de sus efectos sobre los dinosaurios aún son áreas de investigación en curso. [274] Junto a los dinosaurios, muchos otros grupos de animales se extinguieron: pterosaurios, reptiles marinos como mosasaurios y plesiosaurios, varios grupos de mamíferos, amonitas ( moluscos parecidos a nautilos ), rudistas ( bivalvos formadores de arrecifes ) y varios grupos de animales marinos. plancton. [275] [276] En total, aproximadamente el 47% de los géneros y el 76% de las especies de la Tierra se extinguieron durante el evento de extinción K-Pg. [277] El tamaño relativamente grande de la mayoría de los dinosaurios y la baja diversidad de especies de dinosaurios de cuerpo pequeño al final del Cretácico pueden haber contribuido a su extinción; [278] la extinción de los linajes de aves que no sobrevivieron también puede haber sido causada por una dependencia de los hábitats forestales o una falta de adaptaciones para comer semillas para sobrevivir. [279] [280]
Diversidad previa a la extinción
Justo antes del evento de extinción K-Pg, el número de especies de dinosaurios no aviares que existían a nivel mundial se estimó entre 628 y 1078. [281] Sigue siendo incierto si la diversidad de dinosaurios estaba en disminución gradual antes de la extinción K-Pg. evento, o si los dinosaurios realmente prosperaban antes de la extinción. Se ha descubierto que las formaciones rocosas de la época del Maastrichtiano , que precedieron directamente a la extinción, tenían una diversidad menor que la de la época del Campaniano anterior , lo que llevó a la opinión predominante de una disminución a largo plazo de la diversidad. [275] [276] [282] Sin embargo, estas comparaciones no tuvieron en cuenta el potencial de conservación variable entre unidades de roca ni los diferentes grados de exploración y excavación. [274] En 1984, Dale Russell llevó a cabo un análisis para tener en cuenta estos sesgos y no encontró evidencia de una disminución; [283] otro análisis realizado por David Fastovsky y sus colegas en 2004 incluso mostró que la diversidad de dinosaurios aumentó continuamente hasta la extinción, [284] pero este análisis ha sido refutado. [285] Desde entonces, diferentes enfoques basados en estadísticas y modelos matemáticos han apoyado de diversas formas una extinción repentina [274] [281] [286] o un declive gradual. [287] [288] Las tendencias en diversidad del final del Cretácico pueden haber variado entre los linajes de dinosaurios: se ha sugerido que los saurópodos no estaban en declive, mientras que los ornitisquios y los terópodos estaban en declive. [289] [290]
La hipótesis del impacto del bólido , que llamó la atención por primera vez en 1980 por Walter Álvarez , Luis Álvarez y sus colegas, atribuye el evento de extinción del K-Pg al impacto de un bólido (proyectil extraterrestre). [291] Álvarez y sus colegas propusieron que un aumento repentino en los niveles de iridio , registrados en todo el mundo en depósitos de rocas en el límite Cretácico-Paleógeno, era evidencia directa del impacto. [292] También se encontró en todo el mundo cuarzo impactado , indicativo de una fuerte onda de choque que emana de un impacto. [293] El lugar real del impacto siguió siendo difícil de alcanzar hasta que se descubrió un cráter que medía 180 km (110 millas) de ancho en la península de Yucatán, en el sureste de México , y fue publicado en un artículo de 1991 por Alan Hildebrand y sus colegas. [294] Ahora, la mayor parte de la evidencia sugiere que un bólido de 5 a 15 kilómetros (3 a 9+1 ⁄ milla) de ancho impactó la Península de Yucatán hace 66 millones de años, formando este cráter [295] y creando un "mecanismo de muerte" que desencadenó el evento de extinción. [296] [297] [298]
En cuestión de horas, el impacto de Chicxulub habría creado efectos inmediatos como terremotos, [299] tsunamis, [300] y una tormenta de fuego global que probablemente mató a animales desprotegidos y provocó incendios forestales. [301] [302] Sin embargo, también habría tenido consecuencias a más largo plazo para el medio ambiente. En cuestión de días, los aerosoles de sulfato liberados por las rocas en el lugar del impacto habrían contribuido a la lluvia ácida y la acidificación de los océanos . [303] [304] Se cree que los aerosoles de hollín se extendieron por todo el mundo durante los meses y años siguientes; habrían enfriado la superficie de la Tierra al reflejar la radiación térmica y ralentizado enormemente la fotosíntesis al bloquear la luz solar, creando así un impacto invernal . [274] [305] [306] (Esta función se atribuyó a los aerosoles de sulfato hasta que los experimentos demostraron lo contrario. [304] ) El cese de la fotosíntesis habría llevado al colapso de las redes alimentarias que dependían de las plantas frondosas, que incluían a todos los dinosaurios excepto a aves que se alimentan de cereales. [280]
Trampas del Decán
En el momento de la extinción de K-Pg, los basaltos de inundación de Deccan Traps de la India estaban en erupción activa. Las erupciones se pueden dividir en tres fases alrededor del límite K-Pg, dos antes del límite y una después. La segunda fase, que ocurrió muy cerca del límite, habría extruido entre el 70 y el 80% del volumen de estas erupciones en pulsos intermitentes que ocurrieron con unos 100.000 años de diferencia. [307] [308] Esta actividad volcánica habría liberado gases de efecto invernadero como el dióxido de carbono y el dióxido de azufre , [309] [310] , lo que provocó un cambio climático a través de perturbaciones de temperatura de aproximadamente 3 °C (5,4 °F), pero posiblemente como hasta 7 °C (13 °F). [311] Al igual que el impacto de Chicxulub, las erupciones también pueden haber liberado aerosoles de sulfato, lo que habría causado lluvia ácida y enfriamiento global. [312] Sin embargo, debido a los grandes márgenes de error en la datación de las erupciones, el papel de las trampas del Deccan en la extinción de K-Pg sigue sin estar claro. [273] [274] [313]
Antes de 2000, los argumentos de que las erupciones de las trampas del Decán (a diferencia del impacto de Chicxulub) causaron la extinción generalmente estaban vinculados a la opinión de que la extinción era gradual. Antes del descubrimiento del cráter Chicxulub, las trampas del Deccan se utilizaban para explicar la capa global de iridio; [309] [314] Incluso después del descubrimiento del cráter, todavía se pensaba que el impacto solo había tenido un efecto regional, no global, en el evento de extinción. [315] En respuesta, Luis Álvarez rechazó la actividad volcánica como explicación de la capa de iridio y la extinción en su conjunto. [316] Desde entonces, sin embargo, la mayoría de los investigadores han adoptado una posición más moderada, que identifica el impacto de Chicxulub como el principal progenitor de la extinción y al mismo tiempo reconoce que las trampas del Deccan también pueden haber desempeñado un papel. El propio Walter Álvarez ha reconocido que las trampas del Deccan y otros factores ecológicos pueden haber contribuido a las extinciones además del impacto de Chicxulub. [317] Algunas estimaciones han situado el inicio de la segunda fase de las erupciones de las Trampas del Deccan dentro de los 50.000 años posteriores al impacto de Chicxulub. [318] Combinado con el modelado matemático de las ondas sísmicas que habrían sido generadas por el impacto, esto ha llevado a la sugerencia de que el impacto de Chicxulub puede haber desencadenado estas erupciones al aumentar la permeabilidad de la pluma del manto subyacente a las trampas del Deccan. [319] [320]
Aún no se sabe si las trampas del Deccan fueron una causa importante de la extinción, a la par del impacto de Chicxulub. Los defensores consideran que el impacto climático del dióxido de azufre liberado estuvo a la par con el impacto de Chicxulub, y también señalan el papel del vulcanismo de inundación de basalto en otras extinciones masivas como el evento de extinción del Pérmico-Triásico . [321] [322] Consideran que el impacto de Chicxulub ha empeorado el cambio climático en curso causado por las erupciones. [323] Mientras tanto, los detractores señalan la naturaleza repentina de la extinción y que otros pulsos en la actividad de las Trampas del Deccan de magnitud comparable no parecían haber causado extinciones. También sostienen que las causas de las diferentes extinciones masivas deberían evaluarse por separado. [324] En 2020, Alfio Chiarenza y sus colegas sugirieron que las trampas del Deccan pueden incluso haber tenido el efecto contrario: sugirieron que el calentamiento a largo plazo causado por sus emisiones de dióxido de carbono puede haber amortiguado el impacto invernal del impacto de Chicxulub. [298]
Posibles supervivientes del Paleoceno
Ocasionalmente se han encontrado restos de dinosaurios no aviares por encima del límite K-Pg. En 2000, Spencer Lucas y sus colegas informaron del descubrimiento de un único fémur derecho de hadrosaurio en la cuenca de San Juan de Nuevo México , y lo describieron como evidencia de dinosaurios del Paleoceno. La unidad de roca en la que se descubrió el hueso data del Paleoceno temprano , hace aproximadamente 64,8 millones de años. [325] Si el hueso no se volvió a depositar por acción de la erosión, proporcionaría evidencia de que algunas poblaciones de dinosaurios sobrevivieron al menos medio millón de años en el Cenozoico. [326] Otra evidencia incluye la presencia de restos de dinosaurios en la Formación Hell Creek hasta 1,3 m (4,3 pies) por encima del límite Cretácico-Paleógeno, lo que representa 40.000 años de tiempo transcurrido. Esto se ha utilizado para respaldar la opinión de que la extinción de K-Pg fue gradual. [327] Sin embargo, muchos otros investigadores consideran que estos supuestos dinosaurios del Paleoceno fueron reelaborados , es decir, arrastrados fuera de sus ubicaciones originales y luego vueltos a enterrar en sedimentos más jóvenes. [328] [329] [330] Las estimaciones de edad también se han considerado poco confiables. [331]
Según los estándares humanos, los dinosaurios eran criaturas de apariencia fantástica y, a menudo, de tamaño enorme. Como tales, han capturado la imaginación popular y se han convertido en una parte duradera de la cultura humana. La entrada de la palabra "dinosaurio" en la lengua vernácula común refleja la importancia cultural de los animales: en inglés, "dinosaurio" se usa comúnmente para describir cualquier cosa que sea imprácticamente grande, obsoleta o a punto de extinción. [332]
El entusiasmo público por los dinosaurios se desarrolló por primera vez en la Inglaterra victoriana , donde en 1854, tres décadas después de las primeras descripciones científicas de restos de dinosaurios, se inauguró una colección de esculturas de dinosaurios realistas en el Crystal Palace Park de Londres . Los dinosaurios de Crystal Palace resultaron tan populares que pronto se desarrolló un fuerte mercado de réplicas más pequeñas. En las décadas siguientes, se abrieron exhibiciones de dinosaurios en parques y museos de todo el mundo, lo que garantizó que las generaciones sucesivas conocieran a los animales de una manera inmersiva y emocionante. [333] La duradera popularidad de los dinosaurios, a su vez, ha resultado en una importante financiación pública para la ciencia de los dinosaurios y con frecuencia ha estimulado nuevos descubrimientos. En Estados Unidos, por ejemplo, la competencia entre museos por la atención del público condujo directamente a las Guerras de los Huesos de las décadas de 1880 y 1890, durante las cuales un par de paleontólogos enfrentados hicieron enormes contribuciones científicas. [334]
La preocupación popular por los dinosaurios ha asegurado su aparición en la literatura , el cine y otros medios . A partir de 1852, con una mención pasajera en La casa desolada de Charles Dickens , [335] los dinosaurios han aparecido en un gran número de obras de ficción . La novela de Julio Verne de 1864 Viaje al centro de la Tierra , el libro de Sir Arthur Conan Doyle de 1912 El mundo perdido , la película animada de 1914 Gertie el dinosaurio (que presenta el primer dinosaurio animado), la icónica película de 1933 King Kong , la película de 1954 Godzilla y sus numerosas secuelas, la novela más vendida de 1990 Jurassic Park de Michael Crichton y su adaptación cinematográfica de 1993 son sólo algunos ejemplos notables de apariciones de dinosaurios en la ficción. Los autores de obras de no ficción de interés general sobre dinosaurios, incluidos algunos paleontólogos destacados, a menudo han tratado de utilizar a los animales como una forma de educar a los lectores sobre la ciencia en general. Los dinosaurios están omnipresentes en la publicidad ; Numerosas empresas han hecho referencia a los dinosaurios en anuncios impresos o televisados, ya sea para vender sus propios productos o para caracterizar a sus rivales como lentos, tontos u obsoletos. [336] [337]
Universidad de Southampton (29 de septiembre de 2021). "Dos nuevas especies de dinosaurios depredadores grandes con cráneos parecidos a los de un cocodrilo descubiertas en la Isla de Wight". SciTechDaily .
Zhou, Zhonghe (octubre de 2004). "El origen y evolución temprana de las aves: descubrimientos, disputas y perspectivas a partir de la evidencia fósil" (PDF) . Naturwissenschaften . Berlín: Springer Science+Business Media. 91 (10): 455–471. Código Bib : 2004NW..... 91.. 455Z. doi :10.1007/s00114-004-0570-4. ISSN 0028-1042. PMID 15365634. S2CID 3329625. Archivado desde el original (PDF) el 21 de julio de 2011 . Consultado el 6 de noviembre de 2019 .
Sternberg, Charles Mortram (1966) [Edición original publicada por E. Cloutier, impresor del Rey, 1946]. Dinosaurios canadienses . Serie Geológica. vol. 54 (2ª ed.). Ottawa : Museo Nacional de Canadá . LCCN gs46000214. OCLC 1032865683.
Notas
^ Los dinosaurios (incluidas las aves) son miembros del grupo natural Reptilia . Su biología no se corresponde precisamente con la anticuada clase Reptilia de la taxonomía linneana , formada por amniotas de sangre fría sin pelaje ni plumas. Como la taxonomía linneana se formuló para los animales modernos antes del estudio de la evolución y la paleontología, no tiene en cuenta los animales extintos con rasgos intermedios entre las clases tradicionales.
Chiappe, Luis M .; Witmer, Lawrence M. , eds. (2002). Aves mesozoicas: sobre las cabezas de los dinosaurios . Berkeley: Prensa de la Universidad de California. ISBN 978-0-520-20094-4. LCCN 2001044600. OCLC 901747962.
Cowen, Richard (2005). Historia de la vida (4ª ed.). Malden, MA: Blackwell Publishing. ISBN 978-1-4051-1756-2. LCCN 2003027993. OCLC 53970577.La quinta edición del libro está disponible en Internet Archive. Consultado el 19 de octubre de 2019.
Currie, Philip J.; Koppelhus, Eva B.; Shugar, Martín A.; Wright, Joanna L., eds. (2004). Dragones emplumados: estudios sobre la transición de dinosaurios a aves . Vida del pasado. Bloomington, IN : Prensa de la Universidad de Indiana . ISBN 978-0-253-34373-4. LCCN 2003019035. OCLC 52942941.
Desmond, Adrián J. (1975). Los dinosaurios de sangre caliente: una revolución en paleontología . Londres: Blond & Briggs. ISBN 978-0-8037-3755-6. LCCN 76359907. OL 4933052M . Consultado el 30 de octubre de 2019 .
Dique, Gareth; Kaiser, Gary, eds. (2011). Dinosaurios vivos: la historia evolutiva de las aves modernas . Chichester ; Hoboken, Nueva Jersey: Wiley-Blackwell. ISBN 978-0-470-65666-2. LCCN 2010043277. OCLC 729724640.
Farlow, James O.; Brett-Surman, MK, eds. (1997). El dinosaurio completo . Bloomington, IN: Prensa de la Universidad de Indiana. ISBN 978-0-253-33349-0. LCCN 97-23698. OCLC 924985811 . Consultado el 14 de octubre de 2019 .
Foster, John R .; Lucas, Spencer G. , eds. (2006). "Paleontología y Geología de la Formación Morrison del Jurásico Superior". Boletín del Museo de Historia Natural y Ciencia de Nuevo México . Boletín del Museo de Historia Natural y Ciencias de Nuevo México. Albuquerque, NM : Museo de Historia Natural y Ciencia de Nuevo México . 36 . ISSN 1524-4156. OCLC 77520577 . Consultado el 21 de octubre de 2019 .
Gunther, Robert Theodore , ed. (1968) [Impreso por primera vez en Oxford 1945]. Vida y cartas de Edward Lhwyd. Ciencia temprana en Oxford. vol. XIV. Prefacio de Albert Everard Gunther (Reimpresión ed.). Londres: Dawsons de Pall Mall. ISBN 978-0-7129-0292-2. LCCN 22005926. OCLC 43529321 . Consultado el 4 de noviembre de 2019 .
Hansell, Mike (2000). Nidos de pájaros y comportamiento constructivo . Ilustración con pluma y tinta de Raith Overhill. Cambridge : Prensa de la Universidad de Cambridge. ISBN 978-0-521-46038-5. LCCN 99087681. OCLC 876286627 . Consultado el 30 de octubre de 2019 .
Holtz, Thomas R. Jr. (2007). Dinosaurios: la enciclopedia más completa y actualizada para los amantes de los dinosaurios de todas las edades . Ilustrado por Luis V. Rey . Nueva York: Random House . ISBN 978-0-375-82419-7. LCCN 2006102491. OCLC 77486015 . Consultado el 22 de octubre de 2019 .
Lamberto, David; El grupo de diagramas (1990). El libro de datos de dinosaurios: la enciclopedia definitiva y completamente ilustrada de dinosaurios . Nueva York: Libros Avon . ISBN 978-0-380-75896-8. LCCN 89092487. OCLC 21833417 . Consultado el 14 de octubre de 2019 .
Lessem, Don ; Exceso, Donald F. (1993). Enciclopedia de dinosaurios de la Dinosaur Society . Ilustraciones de Tracy Lee Ford; asesores científicos, Peter Dodson, et al. Nueva York: Casa aleatoria. ISBN 978-0-679-41770-5. LCCN 94117716. OCLC 30361459 . Consultado el 30 de octubre de 2019 .
Lhuyd, Eduardo (1699). Lithophylacii Britannici ichnographia [ piedras labradas británicas ]. Londres: Ex Officina MC Archivado desde el original el 14 de agosto de 2020 . Consultado el 4 de noviembre de 2019 .
Mayr, Gerald (2009). Aves fósiles paleógenas. Berlín : Springer-Verlag. doi :10.1007/978-3-540-89628-9. ISBN 978-3-540-89627-2. LCCN 2008940962. OCLC 916182693. S2CID 88941254 . Consultado el 30 de octubre de 2019 .
Norell, Marcos ; Gaffney, Eugene S .; Dingus, Lowell (2000) [Publicado originalmente como Descubriendo dinosaurios en el Museo Americano de Historia Natural . Nueva York: Knopf , 1995]. Descubrimiento de dinosaurios: evolución, extinción y lecciones de la prehistoria (edición revisada). Berkeley: Prensa de la Universidad de California. ISBN 978-0-520-22501-5. LCCN 99053335. OCLC 977125867 . Consultado el 30 de octubre de 2019 .
Olshevsky, George (2000). Una lista de verificación comentada de especies de dinosaurios por continente . Meandros mesozoicos. vol. 3. Ilustrado por Tracy Lee Ford. San Diego, CA : Publicaciones que requieren investigación. ISSN 0271-9428. LCCN 00708700. OCLC 44433611.
Owen, Ricardo (1842). "Informe sobre reptiles fósiles británicos. Parte II". Informe de la Undécima Reunión de la Asociación Británica para el Avance de la Ciencia; Celebrada en Plymouth en julio de 1841. Londres : John Murray . págs. 60-204. ISBN 978-0-8201-1526-9. LCCN 99030427. OCLC 1015526268 . Consultado el 13 de octubre de 2019 .
Padian, Kevin, ed. (1986). El origen de las aves y la evolución del vuelo . Memorias de la Academia de Ciencias de California. vol. 8. San Francisco, CA: Academia de Ciencias de California . ISBN 978-0-940228-14-6. OCLC 946083441. OL 9826926M.
Parsons, Keith M. (2001). Dibujando Leviatán: los dinosaurios y las guerras científicas . La vida en el pasado. Bloomington, IN: Prensa de la Universidad de Indiana. ISBN 978-0-253-33937-9. LCCN 2001016803. OCLC 50174737 . Consultado el 30 de octubre de 2019 .
Pablo, Gregory S. (1988). Dinosaurios depredadores del mundo: una guía ilustrada completa. Nueva York: Simon & Schuster . ISBN 978-0-671-61946-6. LCCN 88023052. OCLC 859819093 . Consultado el 30 de octubre de 2019 .
Pablo, Gregory S., ed. (2000). El libro científico americano de los dinosaurios (1ª ed.). Nueva York: St. Martin's Press . ISBN 978-0-312-26226-6. LCCN 2001269051. OCLC 45256074.
Trama, Robert (1677). La historia natural de Oxford-shire: un ensayo sobre la historia natural de Inglaterra. Oxford; Londres: S. Millers. LCCN 11004267. OCLC 933062622 . Consultado el 13 de noviembre de 2019 .
Sarjeant, William AS , ed. (1995). Fósiles de vertebrados y la evolución de los conceptos científicos: escritos en homenaje a Beverly Halstead, de algunos de sus muchos amigos . Geología moderna, vol. 18. Ámsterdam: Gordon and Breach Publishers. ISBN 978-2-88124-996-9. ISSN 0026-7775. LCCN 00500382. OCLC 34672546."Reimpresión de artículos publicados en un volumen especial de Geología moderna [v. 18 (volumen conmemorativo de Halstead), 1993], con cinco contribuciones adicionales.--Pref."
Tanner, Lawrence H.; Spielmann, Justin A.; Lucas, Spencer G., eds. (2013). "El sistema Triásico: nuevos avances en estratigrafía y paleontología". Boletín del Museo de Historia Natural y Ciencia de Nuevo México . Boletín del Museo de Historia Natural y Ciencias de Nuevo México. Albuquerque, NM: Museo de Historia Natural y Ciencia de Nuevo México. 61 . ISSN 1524-4156. OCLC 852432407 . Consultado el 21 de octubre de 2019 .
^ Mateo G. Barón; Megan E. Williams (2018). "Una reevaluación del enigmático dinosaurio dinosaurio Caseosaurus crosbyensis del Triásico Tardío de Texas, EE. UU. y sus implicaciones para la evolución temprana de los dinosaurios". Acta Paleontológica Polonica. 63. doi : 10.4202/app.00372.2017.
^ Andrea Cau (2018). "El montaje del plan corporal aviar: un proceso de 160 millones de años" (PDF). Bollettino della Società Paleontologica Italiana. 57 (1): 1–25. doi :10.4435/BSPI.2018.01.
^ Ferigolo, Jorge; Langer, Max C. (2007). "Un dinosaurio del Triásico Tardío del sur de Brasil y el origen del hueso predentario ornitisquio". Biología histórica . 19 (1): 23–33. Código Bib : 2007HBio...19...23F. doi :10.1080/08912960600845767. ISSN 0891-2963. S2CID 85819339.
^ Langer, Max C.; Ferigolo, Jorge (2013). "El dinosauriomorfo del Triásico Tardío Sacisaurus agudoensis (Formación Caturrita; Rio Grande do Sul, Brasil): anatomía y afinidades". Sociedad Geológica, Londres, Publicaciones especiales . 379 (1): 353–392. Código Bib : 2013GSLSP.379..353L. doi :10.1144/SP379.16. ISSN 0305-8719. S2CID 131414332.
^ Cabreira, SF; Kellner, AWA; Dias-da-Silva, S.; da Silva, LR; Bronzati, M.; de Almeida Marsola, JC; Müller, RT; de Souza Bittencourt, J.; Batista, BJ; Raugust, T.; Carrilho, R.; Brodt, A.; Langer, MC (2016). "Un conjunto único de dinosauriomorfos del Triásico tardío revela la anatomía y la dieta ancestral de los dinosaurios". Biología actual . 26 (22): 3090–3095. doi : 10.1016/j.cub.2016.09.040 . PMID 27839975.
^ Müller, Rodrigo Temp; García, Maurício Silva (26 de agosto de 2020). "Un 'Silesauridae' parafilético como hipótesis alternativa para la radiación inicial de los dinosaurios ornitisquios". Cartas de biología . 16 (8): 20200417. doi : 10.1098/rsbl.2020.0417 . PMC 7480155 . PMID 32842895.
^ abc "El 'nacimiento' de los dinosaurios". Más que un dodo . 28 de abril de 2017 . Consultado el 15 de marzo de 2023 .
^ abc "El nacimiento de los dinosaurios: Richard Owen y Dinosauria". Biblioteca del Patrimonio de la Biodiversidad . 16 de octubre de 2015 . Consultado el 15 de marzo de 2023 .
^ ab Brett-Surman, MK; Holtz, Thomas R.; Farlow, James O. (27 de junio de 2012). El dinosaurio completo. Prensa de la Universidad de Indiana. pag. 25.ISBN _978-0-253-00849-7.
^ abcd Weishampel, Dodson y Osmólska 2004, págs. 7-19, cap. 1: "Origen y relaciones de Dinosauria" de Michael J. Benton .
^ Olshevsky 2000
^ ab Langer, Max C.; Ezcurra, Martín D .; Bittencourt, Jonathas S.; Novas, Fernando E. (febrero de 2010). "El origen y evolución temprana de los dinosaurios". Reseñas biológicas . Cambridge : Sociedad Filosófica de Cambridge . 85 (1): 65–66, 82. doi :10.1111/j.1469-185x.2009.00094.x. hdl : 11336/103412 . ISSN 1464-7931. PMID 19895605. S2CID 34530296.
^ "Usar el árbol para clasificar". Comprender la evolución . Berkeley : Universidad de California . Archivado desde el original el 31 de agosto de 2019 . Consultado el 14 de octubre de 2019 .
^ abcdefg Weishampel, Dodson y Osmólska 2004, págs. 210-231, cap. 11: "Basal Avialae" de Kevin Padian .
^ Wade, Nicholas (22 de marzo de 2017). "Reorganizando el árbol genealógico de los dinosaurios" . Los New York Times . Nueva York . ISSN 0362-4331. Archivado desde el original el 7 de abril de 2018 . Consultado el 30 de octubre de 2019 ."Una versión de este artículo aparece impresa el 28 de marzo de 2017, en la página D6 de la edición de Nueva York con el titular: Shaking Up the Dinosaur Family Tree".
^ Barón, Matthew G.; Norman, David B .; Barrett, Paul M. (2017). "Una nueva hipótesis sobre las relaciones entre los dinosaurios y la evolución temprana de los dinosaurios". Naturaleza . Londres: Investigación de la naturaleza . 543 (7646): 501–506. Código Bib :2017Natur.543..501B. doi : 10.1038/naturaleza21700. ISSN 0028-0836. PMID 28332513. S2CID 205254710."Este archivo contiene texto y datos complementarios, tablas complementarias 1 a 3 y referencias adicionales.": Información complementaria [ enlace muerto permanente ]
^ Exceso 1997, pag. 40
^ Chamary, JV (30 de septiembre de 2014). "Dinosaurios, pterosaurios y otros saurios: grandes diferencias". Forbes . Jersey City, Nueva Jersey . ISSN 0015-6914. Archivado desde el original el 10 de noviembre de 2014 . Consultado el 2 de octubre de 2018 .
^ Lambert y The Diagram Group 1990, pág. 288
^ Farlow y Brett-Surman 1997, págs. 607–624, cap. 39: "Principales grupos de vertebrados no dinosaurios de la era mesozoica" por Michael Morales.
^ ab Tennant, Jonathan P.; Chiarenza, Alfio Alessandro; Barón, Matthew (19 de febrero de 2018). "¿Cómo ha cambiado nuestro conocimiento sobre la diversidad de dinosaurios a lo largo del tiempo geológico a lo largo de la historia de la investigación?". PeerJ . 6 : e4417. doi : 10.7717/peerj.4417 . PMC 5822849 . PMID 29479504.
^ Starrfelt, Jostein; Liow, Lee Hsiang (2016). "¿Cuántas especies de dinosaurios había? Sesgo fósil y riqueza real estimada utilizando un modelo de muestreo de Poisson". Transacciones Filosóficas de la Royal Society B: Ciencias Biológicas . 371 (1691): 20150219. doi :10.1098/rstb.2015.0219. PMC 4810813 . PMID 26977060.
^ Starrfelt, Jostein; Liow, Lee Hsiang (2016). "¿Cuántas especies de dinosaurios había? Sesgo fósil y riqueza real estimada utilizando un modelo de muestreo de Poisson". Transacciones filosóficas de la Royal Society B. Londres: Sociedad de la Realeza . 371 (1691): 20150219. doi :10.1098/rstb.2015.0219. ISSN 0962-8436. PMC 4810813 . PMID 26977060.
^ Negro, Riley (23 de marzo de 2016). "La mayoría de las especies de dinosaurios aún no se han descubierto". Noticias de National Geographic . Archivado desde el original el 6 de marzo de 2021 . Consultado el 6 de junio de 2021 .
^ Gill, F.; Donsker, D.; Rasmussen, P. (2021). "Bienvenido". Lista Mundial de Aves del COI 11.1 .
^ MacLeod, normando; Rawson, Peter F.; Forey, Peter L.; et al. (1997). "La transición biótica Cretácico-Terciario". Revista de la Sociedad Geológica . Londres: Sociedad Geológica de Londres . 154 (2): 265–292. Código Bib : 1997JGSoc.154..265M. doi :10.1144/gsjgs.154.2.0265. ISSN 0016-7649. S2CID 129654916.
^ ab Amiot, Romain; Buffetaut, Éric ; Lécuyer, Christophe; et al. (2010). "Evidencia de isótopos de oxígeno de hábitos semiacuáticos entre terópodos espinosáuridos". Geología . Boulder, CO : Sociedad Geológica de América . 38 (2): 139-142. Código Bib : 2010Geo....38..139A. doi :10.1130/G30402.1. ISSN 0091-7613.
^ Nesbitt, Sterling J.; Irmis, Randall B.; Parker, William G. (2007). "Una reevaluación crítica de los taxones de dinosaurios del Triásico Tardío de América del Norte". Revista de Paleontología Sistemática . Milton Park, Oxfordshire: Taylor & Francis en nombre del Museo de Historia Natural de Londres . 5 (2): 209–243. Código Bib : 2007JSPal...5..209N. doi :10.1017/S1477201907002040. ISSN 1477-2019. S2CID 28782207.
^ Esto se reconoció a más tardar en 1909: Celeskey, Matt (2005). "Dr. WJ Holland y los saurópodos en expansión". El Museo Peludo de Historia Natural . Archivado desde el original el 12 de junio de 2011 . Consultado el 18 de octubre de 2019 .
Holanda, William J. (mayo de 1910). "Una revisión de algunas críticas recientes a las restauraciones de dinosaurios saurópodos existentes en los museos de los Estados Unidos, con especial referencia a la de Diplodocus Carnegiei en el Museo Carnegie". El naturalista americano . Sociedad Estadounidense de Naturalistas . 44 (521): 259–283. doi :10.1086/279138. ISSN 0003-0147. S2CID 84424110 . Consultado el 18 de octubre de 2019 .
Los argumentos y muchas de las imágenes también se presentan en Desmond 1975.
^ ab Kubo, Tai; Benton, Michael J. (noviembre de 2007). "Evolución de la postura de las extremidades posteriores en arcosaurios: tensiones en las extremidades en vertebrados extintos" (PDF) . Paleontología . Hoboken, Nueva Jersey : Wiley-Blackwell . 50 (6): 1519-1529. Código Bib : 2007Palgy..50.1519K. doi :10.1111/j.1475-4983.2007.00723.x. ISSN 0031-0239. S2CID 140698705.
^ Dong 1992
^ "Huesos de dinosaurio 'utilizados como medicina'". Noticias de la BBC . Londres: BBC. 6 de julio de 2007. Archivado desde el original el 27 de agosto de 2019 . Consultado el 4 de noviembre de 2019 .
^ Paul 2000, págs. 10–44, cap. 1: "Una breve historia de la paleontología de los dinosaurios" por Michael J. Benton.
^ ab Farlow y Brett-Surman 1997, págs. 3-11, cap. 1: "Los primeros descubrimientos" de William AS Sarjeant .
^ "Trama de Robert" (PDF) . Aprendiendo más . Oxford: Museo de Historia Natural de la Universidad de Oxford . 2006. Archivado desde el original (PDF) el 1 de octubre de 2006 . Consultado el 14 de noviembre de 2019 .
^ Lhuyd 1699, pag. 67
^ Delair, Justin B.; Sarjeant, William AS (2002). "Los primeros descubrimientos de dinosaurios: los registros reexaminados". Actas de la Asociación de Geólogos . Ámsterdam: Elsevier en nombre de la Asociación de Geólogos . 113 (3): 185-197. Código Bib : 2002PrGA..113..185D. doi :10.1016/S0016-7878(02)80022-0. ISSN 0016-7878.
^ Gunther 1968
^ Buckland, William (1824). "Aviso sobre el Megalosaurus o gran lagarto fósil de Stonesfield". Transacciones de la Sociedad Geológica de Londres . Londres: Sociedad Geológica de Londres. 1 (2): 390–396. doi :10.1144/transgslb.1.2.390. ISSN 2042-5295. S2CID 129920045. Archivado (PDF) desde el original el 21 de octubre de 2019 . Consultado el 5 de noviembre de 2019 .
^ Farlow y Brett-Surman 1997, págs.14, cap. 2: "Cazadores de dinosaurios europeos" de Hans-Dieter Sues.
^ ab Owen 1842, p.103: "Se presume que la combinación de tales caracteres ... se considerará motivo suficiente para establecer una tribu o suborden distinto de reptiles saurios, para lo cual propondría el nombre de Dinosauria *. (*Gr. δεινός , terriblemente grande; σαύρος , un lagarto. ...)
^ Farlow y Brett-Surman 1997, págs. ix-xi, Prefacio, "Dinosaurios: el superlativo terrestre" de James O. Farlow y MK Brett-Surman.
^ Rupke 1994
^ Prieto-Márquez, Albert; Weishampel, David B.; Horner, John R. (marzo de 2006). «El dinosaurio Hadrosaurus foulkii, del Campaniano de la Costa Este de Norteamérica, con una reevaluación del género» (PDF) . Acta Paleontológica Polonica . Varsovia: Instituto de Paleobiología, Academia Polaca de Ciencias. 51 (1): 77–98. ISSN 0567-7920. Archivado (PDF) desde el original el 22 de junio de 2019 . Consultado el 5 de noviembre de 2019 .
^ Holmes 1998
^ abc Taylor, diputado (2010). "Investigación de dinosaurios saurópodos: una revisión histórica". Sociedad Geológica, Londres, Publicaciones especiales . 343 (1): 361–386. Código Bib : 2010GSLSP.343..361T. doi :10.1144/SP343.22. S2CID 910635.
^ Naish, D. (2009). Los grandes descubrimientos de los dinosaurios . Londres, Reino Unido: A & C Black Publishers Ltd. págs. 89–93. ISBN978-1-4081-1906-8.
^ Cenador, V. (2018). "Los resultados llegan desde el renacimiento de los dinosaurios". Ciencia . 360 (6389): 611. Bibcode : 2018Sci...360..611A. doi : 10.1126/ciencia.aat0451. S2CID 46887409.
^ ab Bakker, RT (1968). "La superioridad de los dinosaurios". Descubrimiento: Revista del Museo Peabody de Historia Natural . 3 (2): 11–22. ISSN 0012-3625. OCLC 297237777.
^ ab Bakker, RT (1972). "Evidencia anatómica y ecológica de endotermia en dinosaurios". Naturaleza . 238 (5359): 81–85. Código Bib :1972Natur.238...81B. doi :10.1038/238081a0. S2CID 4176132.
^ Bakker 1986
^ ab Benton, MJ (2008). "Calidad de los fósiles y denominación de dinosaurios". Cartas de biología . 4 (6): 729–732. doi :10.1098/rsbl.2008.0402. PMC 2614166 . PMID 18796391.
^ ab Cashmore, DD; Mannion, PD; Upchurch, P.; Mayordomo, RJ (2020). "Diez años más de descubrimiento: revisando la calidad del registro fósil de dinosaurio sauropodomorfo". Paleontología . 63 (6): 951–978. Código Bib : 2020Palgy..63..951C. doi : 10.1111/pala.12496 . S2CID 219090716.
^ Cashmore, DD; Mayordomo, RJ (2019). "Completitud esquelética del registro fósil de dinosaurios terópodos no aviares". Paleontología . 62 (6): 951–981. Código Bib : 2019Palgy..62..951C. doi : 10.1111/pala.12436 . S2CID 197571209.
^ Holtz, TR Jr.; Brett-Surman, MK (1997). "La taxonomía y sistemática de los dinosaurios". El dinosaurio completo . Bloomington: Prensa de la Universidad de Indiana. págs. 209–223. ISBN978-0-253-33349-0.
^ abc St. Fleur, Nicholas (8 de diciembre de 2016). "¿Esa cosa con plumas atrapada en ámbar? Era la cola de un dinosaurio" . Trilobites. Los New York Times . Nueva York. ISSN 0362-4331. Archivado desde el original el 31 de agosto de 2017 . Consultado el 8 de diciembre de 2016 .
^ Lockley, MG; Wright, JL (2000). "Lectura sobre dinosaurios: una bibliografía comentada de libros". Revista de Educación en Geociencias . 48 (2): 167-178. Código bibliográfico : 2000JGeEd..48..167L. doi :10.5408/1089-9995-48.2.167. S2CID 151426669.
^ Lloyd, GT; Davis, KE; Pisani, D.; Tarver, JE; Ruta, R.; Sakamoto, M.; Perfecto, DWE; Jennings, R.; Benton, MJ (2008). "Los dinosaurios y la revolución terrestre del Cretácico". Actas de la Royal Society B. 275 (1650): 2483–2490. doi :10.1098/rspb.2008.0715. PMC 2603200 . PMID 18647715.
^ ab Schweitzer, MH (2011). "Preservación de tejidos blandos en vertebrados mesozoicos terrestres". Revista Anual de Ciencias de la Tierra y Planetarias . 39 : 187–216. Código Bib : 2011AREPS..39..187S. doi : 10.1146/annurev-earth-040610-133502.
^ ab Hooley, RW (1917). "II—Sobre el tegumento de Iguanodon bernissartensis, Boulenger, y de Morosaurus becklesii, Mantell". Revista Geológica . 4 (4): 148-150. Código Bib : 1917GeoM....4..148H. doi :10.1017/s0016756800192386. S2CID 129640665.
^ Osborn, HF (1912). "Tegumento del dinosaurio iguanodonte Trachodon ". Memorias del Museo Americano de Historia Natural . 1 : 33–54.
^ Bell, PR (2014). "Una revisión de las impresiones de la piel de hadrosaurios". En Eberth, D.; Evans, D. (eds.). Los hadrosaurios: actas del Simposio internacional sobre hadrosaurios . Bloomington: Prensa de la Universidad de Princeton. págs. 572–590.
^ Eliason, CM; Hudson, L.; vatios, T.; Garza, H.; Clarke, JA (2017). "Preservación excepcional y registro fósil del tegumento de tetrápodos". Actas de la Royal Society B. 284 (1862): 1–10. doi :10.1098/rspb.2017.0556. PMC 5597822 . PMID 28878057.
^ Benton, MJ (1998). «Fósiles de dinosaurios con partes blandas» (PDF) . Tendencias en ecología y evolución . 13 (8): 303–304. doi :10.1016/s0169-5347(98)01420-7. PMID 21238317.
^ Zhou, Z.-H.; Wang, Y. (2017). "Conjuntos de vertebrados de la biota Yanliao del Jurásico y la biota Jehol del Cretácico temprano: comparaciones e implicaciones". Paleomundo . 26 (2): 241–252. doi :10.1016/j.palwor.2017.01.002.
^ Norell, MA; Xu, X. (2005). "Dinosaurios emplumados". Revista Anual de Ciencias de la Tierra y Planetarias . 33 : 277–299. Código Bib : 2005AREPS..33..277N. doi : 10.1146/annurev.earth.33.092203.122511.
^ ab Roy, A.; Pittman, M.; Saitta, et al; Kaye, TG; Xu, X. (2020). "Avances recientes en la reconstrucción del paleocolor de amniotas y un marco para futuras investigaciones". Reseñas biológicas . 95 (1): 22–50. doi :10.1111/brv.12552. PMC 7004074 . PMID 31538399.
^ Vinther, J. (2020). "Reconstrucción de vertebrados paleocolor". Revista Anual de Ciencias de la Tierra y Planetarias . 48 : 345–375. Código Bib : 2020AREPS..48..345V. doi : 10.1146/annurev-earth-073019-045641 . S2CID 219768255.
^ Zhang, F.; Kearns, SL; Orr, PJ; Benton, MJ; Zhou, Z.; Johnson, D.; Xu, X.; Wang, X. (2010). "Melanosomas fosilizados y el color de los dinosaurios y aves del Cretácico" (PDF) . Naturaleza . 463 (7284): 1075–1078. Código Bib : 2010Natur.463.1075Z. doi : 10.1038/naturaleza08740. PMID 20107440. S2CID 205219587.
^ Smithwick, FM; Nicholls, R.; Cuthill, IC; Vinther, J. (2017). "El contrasombreado y las rayas en el dinosaurio terópodo Sinosauropteryx revelan hábitats heterogéneos en la biota Jehol del Cretácico temprano". Biología actual . 27 (21): 3337–3343.e2. doi : 10.1016/j.cub.2017.09.032 . hdl : 1983/8ee95b15-5793-42ad-8e57-da6524635349 . PMID 29107548.
^ Vinther, J.; Nicholls, R.; Lautenschlager, S.; Pittman, M.; Kaye, TG; Rayfield, E.; Mayr, G.; Cuthill, IC (2016). "Camuflaje 3D en un dinosaurio ornitisquio". Biología actual . 26 (18): 2456–2462. doi :10.1016/j.cub.2016.06.065. PMC 5049543 . PMID 27641767.
^ Lindgren, J.; Moyer, A.; Schweitzer, MH; Sjövall, P.; Uvdal, P.; Nilsson, DE; Heimdal, J.; Engdahl, A.; Gren, JA; Schultz, BP; Kear, BP (2015). "Interpretación de la coloración basada en melanina a través del tiempo profundo: una revisión crítica". Actas de la Royal Society B. 282 (1813): 20150614. doi :10.1098/rspb.2015.0614. PMC 4632609 . PMID 26290071.
^ Schweitzer, MH; Lindgren, J.; Moyer, AE (2015). "Reexamen de los melanosomas y la coloración antigua: una respuesta a Vinther 2015 (DOI 10.1002/bies.201500018)". Bioensayos . 37 (11): 1174-1183. doi :10.1002/bies.201500061. PMID 26434749. S2CID 45178498.
^ Zhou, Z. (2014). "La Jehol Biota, una Lagerstätte terrestre del Cretácico Inferior: nuevos descubrimientos e implicaciones". Revista Nacional de Ciencias . 1 (4): 543–559. doi : 10.1093/nsr/nwu055 .
^ O'Connor, JK; Zhou, Z. (2019). "La evolución del sistema digestivo aviar moderno: conocimientos de los fósiles paravianos de las biotas de Yanliao y Jehol". Paleontología . 63 (1): 13–27. doi :10.1111/pala.12453. S2CID 210265348.
^ ab Dal Sasso, Cristiano ; Signore, Marco (26 de marzo de 1998). "Preservación excepcional de tejidos blandos en un dinosaurio terópodo de Italia" (PDF) . Naturaleza . Londres: Investigación de la naturaleza. 392 (6674): 383–387. Código Bib :1998Natur.392..383D. doi :10.1038/32884. ISSN 0028-0836. S2CID 4325093. Archivado (PDF) desde el original el 20 de septiembre de 2016.
^ Morell, V. (1993). "Dino DNA: la caza y el bombo". Ciencia . 261 (5118): 160–162. Código bibliográfico : 1993 Ciencia... 261.. 160 M. doi : 10.1126/ciencia.8327889. PMID 8327889.
^ Pawlicki, R.; Korbel, A.; Kubiak, H. (1996). "Células, fibrillas de colágeno y vasos en huesos de dinosaurio". Naturaleza . 211 (5049): 655–657. doi :10.1038/211655a0. PMID 5968744. S2CID 4181847.
^ Pawlicki, R.; Nowogrodzka-Zagórska, M. (1998). "Vasos sanguíneos y glóbulos rojos conservados en huesos de dinosaurio". Anales de anatomía - Anatomischer Anzeiger . 180 (1): 73–77. doi :10.1016/S0940-9602(98)80140-4. PMID 9488909.
^ ab Schweitzer, María H .; Wittmeyer, Jennifer L.; Horner, John R .; Toporski, Jan K. (2005). "Vasos de tejidos blandos y preservación celular en Tyrannosaurus rex ". Ciencia . Washington, DC: Asociación Estadounidense para el Avance de la Ciencia. 307 (5717): 1952-1955. Código Bib : 2005 Ciencia... 307.1952S. doi : 10.1126/ciencia.1108397. ISSN 0036-8075. PMID 15790853. S2CID 30456613.
^ Anderson, Luisiana (2023). "Un marco químico para la preservación de células y tejidos blandos de vertebrados fósiles". Reseñas de ciencias de la tierra . 240 : 104367. Código bibliográfico : 2023ESRv..24004367A. doi : 10.1016/j.earscirev.2023.104367 . S2CID 257326012.
^ ab Schweitzer, MH; Zheng, W.; Órgano, CL; Avci, R.; Suo, Z.; Freimark, LM; LeBleu, VS; Duncan, MB; van der Heiden, MG; Neveu, JM; Carril, WS; Cottrell, JS; Horner, JR; Cantley, LC; Kalluri, R.; Asara, JM (2009). "Caracterización biomolecular y secuencias de proteínas del hadrosaurio B. canadensis del Campaniano ". Ciencia . 324 (5927): 626–631. Código Bib : 2009 Ciencia... 324..626S. doi : 10.1126/ciencia.1165069. PMID 19407199. S2CID 5358680.
^ Órgano, CL; Schweitzer, MH; Zheng, W.; Freimark, LM; Cantley, LC; Asara, JM (2008). "Filogenética molecular de mastodonte y tiranosaurio rex ". Ciencia . 320 (5875): 499. Código bibliográfico : 2008Sci...320..499O. doi : 10.1126/ciencia.1154284. PMID 18436782. S2CID 24971064.
^ Schweitzer, MH; Zheng, W.; Cleland, TP; Berna, M. (2013). "Los análisis moleculares de osteocitos de dinosaurios respaldan la presencia de moléculas endógenas". Hueso . Ámsterdam: Elsevier. 52 (1): 414–423. doi :10.1016/j.bone.2012.10.010. ISSN 8756-3282. PMID 23085295.
^ Bailleul, AM; Zheng, W.; Horner, JR; Salón, BK; Vacaciones, CM; Schweitzer, MH (2020). "Evidencia de proteínas, cromosomas y marcadores químicos de ADN en cartílago de dinosaurio excepcionalmente conservado". Revista Nacional de Ciencias . 7 (4): 815–822. doi :10.1093/nsr/nwz206. PMC 8289162 . PMID 34692099.
^ Bertazzo, S.; Sirvienta, SCR ; Kallepitis, C.; et al. (2015). "Fibras y estructuras celulares conservadas en especímenes de dinosaurios de 75 millones de años". Comunicaciones de la naturaleza . 6 : 7352. Código Bib : 2015NatCo...6.7352B. doi : 10.1038/ncomms8352. ISSN 2041-1723. PMC 4468865 . PMID 26056764.
^ Kaye, TG; Gaugler, G.; Sawlowicz, Z. (2008). "Los tejidos blandos de los dinosaurios interpretados como biopelículas bacterianas". MÁS UNO . 3 (7): e2808. Código Bib : 2008PLoSO...3.2808K. doi : 10.1371/journal.pone.0002808 . PMC 2483347 . PMID 18665236.
^ Peterson, JE; Lenczewski, ME; Scherer, RP (2010). "Influencia de las biopelículas microbianas en la preservación del tejido blando primario en arcosaurios fósiles y existentes". MÁS UNO . 5 (10): e13334. Código Bib : 2010PLoSO...513334P. doi : 10.1371/journal.pone.0013334 . ISSN 1932-6203. PMC 2953520 . PMID 20967227.
^ Buckley, M.; Warwood, S.; van Dongen, B.; Cocina, aire acondicionado; Manning, PL (2017). "Una quimera de proteína fósil; dificultades para discriminar secuencias de péptidos de dinosaurios de la contaminación cruzada moderna". Actas de la Royal Society B. 284 (1855). doi :10.1098/rspb.2017.0544. PMC 5454271 . PMID 28566488.
^ Kump, Lee R .; Pávlov, Alejandro; Arturo, Michael A. (2005). "Liberación masiva de sulfuro de hidrógeno a la superficie del océano y la atmósfera durante intervalos de anoxia oceánica". Geología . Boulder, CO: Sociedad Geológica de América. 33 (5): 397–400. Código Bib : 2005Geo....33..397K. doi :10.1130/G21295.1. ISSN 0091-7613. S2CID 34821866.
^ Tanner, Lawrence H.; Lucas, Spencer G .; Chapman, Mary G. (marzo de 2004). "Evaluación del registro y las causas de las extinciones del Triásico Tardío" (PDF) . Reseñas de ciencias de la tierra . Ámsterdam: Elsevier. 65 (1–2): 103–139. Código Bib : 2004ESRv...65..103T. doi :10.1016/S0012-8252(03)00082-5. ISSN 0012-8252. Archivado desde el original (PDF) el 25 de octubre de 2007 . Consultado el 22 de octubre de 2007 .
^ ab Griffin, CT; Wynd, BM; Munyikwa, D.; Broderick, TJ; Zondo, M.; Tolan, S.; Langer, MC; Nesbitt, SJ; Taruvinga, Recursos Humanos (2022). "Los dinosaurios más antiguos de África revelan una supresión temprana de la distribución de los dinosaurios". Naturaleza . 609 (7926): 313–319. Código Bib :2022Natur.609..313G. doi :10.1038/s41586-022-05133-x. ISSN 0028-0836. PMID 36045297. S2CID 251977824.
^ Desojo, JB; Fiorelli, LE; Ezcurra, MD; Martinelli, AG; Ramezani, J.; Da Rosa, AAS; Belén von Baczko, M.; Jimena Trotteyn, M.; Montefeltro, FC; Ezpeleta, M.; Langer, MC (2020). "La Formación Ischigualasto del Triásico Tardío en Cerro Las Lajas (La Rioja, Argentina): tetrápodos fósiles, cronoestratigrafía de alta resolución y correlaciones faunísticas". Informes científicos . 10 (1): 12782. Código bibliográfico : 2020NatSR..1012782D. doi :10.1038/s41598-020-67854-1. PMC 7391656 . PMID 32728077.
^ Alcober, Óscar A.; Martínez, Ricardo N. (2010). "Un nuevo herrerasáurido (Dinosauria, Saurischia) de la Formación Ischigualasto del Triásico Superior del noroeste de Argentina". Llaves del zoológico . Sofía : Pensoft Publishers (63): 55–81. Código Bib : 2010ZooK...63...55A. doi : 10.3897/zookeys.63.550 . ISSN 1313-2989. PMC 3088398 . PMID 21594020.
^ abc Novas, FE; Agnolín, Florida; Ezcurra, MD; Müller, RT; Martinelli, A.; Langer, M. (2021). "Revisión del registro fósil de los primeros dinosaurios de América del Sur y sus implicaciones filogenéticas". Revista de Ciencias de la Tierra Sudamericana . 110 : 103341. Código bibliográfico : 2021JSAES.11003341N. doi : 10.1016/j.jsames.2021.103341. ISSN 0895-9811.
^ Nesbitt, Sterling J; Demandas, Hans-Dieter (2021). "La osteología del dinosaurio Daemonosaurus chauliodus (Archosauria: Dinosauria) de la cantera Coelophysis (Triásico: Rético) de Nuevo México y sus relaciones con otros dinosaurios primitivos". Revista zoológica de la Sociedad Linneana . 191 (1): 150-179. doi : 10.1093/zoolinnean/zlaa080 .
^ ab Sereno, Paul C. (1999). "La evolución de los dinosaurios". Ciencia . Washington, DC: Asociación Estadounidense para el Avance de la Ciencia . 284 (5423): 2137–2147. doi : 10.1126/ciencia.284.5423.2137. ISSN 0036-8075. PMID 10381873. Archivado (PDF) desde el original el 5 de enero de 2018 . Consultado el 8 de noviembre de 2019 .
^ Sereno, Paul C.; Forster, Catherine A .; Rogers, Raymond R .; Monetta, Alfredo M. (1993). "Esqueleto de dinosaurio primitivo de Argentina y la evolución temprana de Dinosauria". Naturaleza . Londres: Investigación de la naturaleza. 361 (6407): 64–66. Código Bib :1993Natur.361...64S. doi :10.1038/361064a0. ISSN 0028-0836. S2CID 4270484.
^ ab Langer, Max C.; Ramezani, Jahandar; Da Rosa, Átila AS (mayo de 2018). "Limitaciones de edad U-Pb sobre el surgimiento de dinosaurios en el sur de Brasil". Investigación de Gondwana . Ámsterdam: Elsevier. 57 : 133-140. Código Bib : 2018GondR..57..133L. doi :10.1016/j.gr.2018.01.005. ISSN 1342-937X.
^ Novas, FE; Ezcurra, MD; Chatterjee, S.; Kutty, TS (2011). "Nuevas especies de dinosaurios del Triásico Superior, las formaciones Alto Maleri y Bajo Dharmaram del centro de la India". Transacciones de ciencias ambientales y de la tierra de la Real Sociedad de Edimburgo . 101 (3–4): 333–349. doi :10.1017/S1755691011020093. S2CID 128620874.
^ Marsicano, California; Irmis, RB; Mancuso, AC; Mundil, R.; Chemale, F. (2016). "La calibración temporal precisa de los orígenes de los dinosaurios". Procedimientos de la Academia Nacional de Ciencias . 113 (3): 509–513. Código Bib : 2016PNAS..113..509M. doi : 10.1073/pnas.1512541112 . PMC 4725541 . PMID 26644579.
^ Nesbitt, Sterling J.; Barrett, Paul M.; Werning, Sara; et al. (2012). "¿El dinosaurio más antiguo? Un dinosaurio del Triásico Medio de Tanzania". Cartas de biología . Londres: Sociedad de la Realeza. 9 (1): 20120949. doi :10.1098/rsbl.2012.0949. ISSN 1744-9561. PMC 3565515 . PMID 23221875.
^ Marsicano, California; Irmis, RB; Mancuso, AC; Mundil, R.; Chemale, F. (2015). "La calibración temporal precisa de los orígenes de los dinosaurios". Procedimientos de la Academia Nacional de Ciencias . 113 (3): 509–513. Código Bib : 2016PNAS..113..509M. doi : 10.1073/pnas.1512541112 . ISSN 0027-8424. PMC 4725541 . PMID 26644579.
^ Brusatte, Stephen L .; Benton, Michael J.; Ruta, Marcello; Lloyd, Graeme T. (2008). "Superioridad, competencia y oportunismo en la radiación evolutiva de los dinosaurios" (PDF) . Ciencia . Washington, DC: Asociación Estadounidense para el Avance de la Ciencia. 321 (5895): 1485-1488. Código Bib : 2008 Ciencia... 321.1485B. doi : 10.1126/ciencia.1161833. hdl :20.500.11820/00556baf-6575-44d9-af39-bdd0b072ad2b. ISSN 0036-8075. PMID 18787166. S2CID 13393888. Archivado (PDF) desde el original el 19 de julio de 2018 . Consultado el 22 de octubre de 2019 .
^ Tanner, Spielmann & Lucas 2013, págs. 562–566, "El primer rincosaurio noriano (revueltiano): formación Bull Canyon, Nuevo México, EE. UU." por Justin A. Spielmann, Spencer G. Lucas y Adrian P. Hunt.
^ Sulej, Tomasz; Niedźwiedzki, Grzegorz (2019). "Un sinápsido del Triásico Tardío del tamaño de un elefante con extremidades erectas". Ciencia . Washington, DC: Asociación Estadounidense para el Avance de la Ciencia. 363 (6422): 78–80. Código Bib : 2019 Ciencia... 363... 78S. doi : 10.1126/science.aal4853 . ISSN 0036-8075. PMID 30467179. S2CID 53716186.
^ "Las huellas de fósiles en los Alpes ayudan a explicar la evolución de los dinosaurios" . Ciencia y Tecnología. El economista . Londres. 19 de abril de 2018. ISSN 0013-0613 . Consultado el 24 de mayo de 2018 .
^ abcd Weishampel, Dodson y Osmólska 2004, págs. 627–642, cap. 27: "Biogeografía mesozoica de Dinosauria" por Thomas R. Holtz Jr., Ralph E. Chapman y Matthew C. Lamanna .
^ abcde Weishampel, Dodson y Osmólska 2004, págs. 614–626, cap. 26: "Paleoecología de dinosaurios" de David E. Fastovsky y Joshua B. Smith.
^ Sereno, Paul C.; Wilson, Jeffrey A .; Witmer, Lawrence M .; et al. (2007). Kemp, Tom (ed.). "Extremos estructurales en un dinosaurio del Cretácico". MÁS UNO . San Francisco, CA : MÁS . 2 (11): e1230. Código Bib : 2007PLoSO...2.1230S. doi : 10.1371/journal.pone.0001230 . ISSN 1932-6203. PMC 2077925 . PMID 18030355.
^ Prasad, Vandana; Strömberg, Caroline AE; Alimohammadian, Habib; et al. (2005). "Coprolitos de dinosaurios y la evolución temprana de pastos y herbívoros". Ciencia . Washington, DC: Asociación Estadounidense para el Avance de la Ciencia. 310 (5751): 1170–1180. Código Bib : 2005 Ciencia... 310.1177P. doi : 10.1126/ciencia.1118806. ISSN 0036-8075. PMID 16293759. S2CID 1816461.
^ Weishampel, Dodson y Osmólska 2004, págs. 672–684, cap. 30: "Extinción de dinosaurios" de J. David Archibald y David E. Fastovsky.
^ Dyke y Kaiser 2011, cap. 14: "Evolución de las aves a través del límite K-Pg y la diversificación basal neornitina" por Bent EK Lindow. doi :10.1002/9781119990475.ch14
^ Cracraft, Joel (1968). "Una revisión de Bathornithidae (Aves, Gruiformes), con comentarios sobre las relaciones del suborden Cariamae" (PDF) . Novitates del Museo Americano . Nueva York: Museo Americano de Historia Natural (2326): 1–46. hdl :2246/2536. ISSN 0003-0082 . Consultado el 22 de octubre de 2019 .
^ Alvarenga, Herculano ; Jones, Washington W.; Rinderknecht, Andrés (mayo de 2010). "El registro más joven de aves forusrácidas (Aves, Phorusrhacidae) del Pleistoceno tardío del Uruguay". Neues Jahrbuch für Geologie und Paläontologie . Stuttgart : E. Schweizerbart . 256 (2): 229–234. doi :10.1127/0077-7749/2010/0052. ISSN 0077-7749 . Consultado el 22 de octubre de 2019 .
^ Mayo 2009
^ Pablo 1988, págs. 248-250
^ Weishampel, Dodson y Osmólska 2004, págs. 151-164, cap. 7: "Therizinosauroidea" de James M. Clark, Teresa Maryańska y Rinchen Barsbold .
^ Weishampel, Dodson y Osmólska 2004, págs. 196-210, cap. 10: "Dromaeosauridae" de Peter J. Makovicky y Mark A. Norell .
^ Taylor, Michael P .; Wedel, Mathew J. (2013). "Por qué los saurópodos tenían el cuello largo y por qué las jirafas tenían el cuello corto". PeerJ . Corte Madera, California ; Londres. 1 : e36. doi : 10.7717/peerj.36 . ISSN 2167-8359. PMC 3628838 . PMID 23638372.
^ Justin tuiteó. "Diagramas de clasificación". Minnesota ecuatorial . Consultado el 6 de septiembre de 2022 .
^ abc Alexander, R. McNeill (2006). "Biomecánica de los dinosaurios". Actas de la Royal Society B. Londres: Sociedad de la Realeza. 273 (1596): 1849–1855. doi :10.1098/rspb.2006.3532. ISSN 0962-8452. PMC 1634776 . PMID 16822743.
^ Dodson y Gingerich 1993, págs. 167-199, "Sobre la rareza de los animales grandes y feroces: especulaciones sobre el tamaño del cuerpo, las densidades de población y la distribución geográfica de los mamíferos depredadores y los grandes dinosaurios carnívoros" por James O. Farlow.
^ "Evolución de los dinosaurios". Departamento de Paleobiología . Dinosaurios. Washington, DC: Museo Nacional de Historia Natural . 2007. Archivado desde el original el 11 de noviembre de 2007 . Consultado el 21 de noviembre de 2007 .
^ abc Sander, P. Martín; Cristiano, Andrés; Clauss, Marco; et al. (febrero de 2011). "Biología de los dinosaurios saurópodos: la evolución del gigantismo". Reseñas biológicas . Cambridge: Sociedad Filosófica de Cambridge. 86 (1): 117-155. doi :10.1111/j.1469-185X.2010.00137.x. ISSN 1464-7931. PMC 3045712 . PMID 21251189.
^ abc Foster & Lucas 2006, págs. 131-138, "El más grande de los grandes: una reevaluación crítica del megasaurópodo Amphicoelias fragillimus Cope, 1878" por Kenneth Carpenter .
^ Pablo 2010
^ Colberto 1971
^ Mazzetta, Gerardo V.; Christiansenb, Per; Fariñaa, Richard A. (2004). "Gigantes y extraños: tamaño corporal de algunos dinosaurios del Cretácico del sur de América del Sur" (PDF) . Biología histórica . Milton Park, Oxfordshire: Taylor y Francis. 16 (2–4): 71–83. Código Bib : 2004HBio...16...71M. CiteSeerX 10.1.1.694.1650 . doi :10.1080/08912960410001715132. ISSN 0891-2963. S2CID 56028251. Archivado (PDF) desde el original el 25 de febrero de 2009.
^ Janensch, Werner (1950). Traducción de Gerhard Maier. "Die Skelettrekonstruktion von Brachiosaurus brancai" [La reconstrucción del esqueleto de Brachiosaurus brancai] (PDF) . Paleontográfica . Stuttgart: E. Schweizerbart . Suplemento VII (1. Reihe, Teil 3, Lieferung 2): 97–103. OCLC 45923346. Archivado (PDF) desde el original el 11 de julio de 2017 . Consultado el 24 de octubre de 2019 .
^ Lucas, Spencer G.; Herne, Mateo C.; Hecket, Andrew B.; et al. (2004). Reevaluación del Seismosaurus, un dinosaurio saurópodo del Jurásico tardío de Nuevo México. Reunión anual de Denver de 2004 (7 al 10 de noviembre de 2004). vol. 36. Boulder, CO: Sociedad Geológica de América. pag. 422. OCLC 62334058. Documento No. 181-4. Archivado desde el original el 8 de octubre de 2019 . Consultado el 25 de octubre de 2019 .
^ Vendedores, William Irvin.; Margetts, Lee; Coria, Rodolfo Aníbal ; Manning, Phillip Lars (2013). Portador, David (ed.). "Marcha de los titanes: las capacidades locomotoras de los dinosaurios saurópodos". MÁS UNO . San Francisco, CA: MÁS. 8 (10): e78733. Código Bib : 2013PLoSO...878733S. doi : 10.1371/journal.pone.0078733 . ISSN 1932-6203. PMC 3864407 . PMID 24348896.
^ Lovelace, David M.; Hartman, Scott A.; Wahl, William R. (octubre-diciembre de 2007). "Morfología de un espécimen de Supersaurus (Dinosauria, Sauropoda) de la Formación Morrison de Wyoming y una reevaluación de la filogenia de los diplodócidos". Archivos del Museo Nacional . Río de Janeiro : Museo Nacional de Brasil ; Universidad Federal de Río de Janeiro . 65 (4): 527–544. CiteSeerX 10.1.1.603.7472 . ISSN 0365-4508 . Consultado el 26 de octubre de 2019 .
^ Carpintero, Kenneth (2018). "Maraapunisaurus fragillimus, NG (anteriormente Amphicoelias fragillimus), un Rebbachisáurido basal de la Formación Morrison (Jurásico superior) de Colorado". Geología del Oeste Intermontañoso . 5 : 227–244. doi : 10.31711/giw.v5i0.28 .
^ Pablo, Gregory S. (2019). "Determinación del animal terrestre más grande conocido: una comparación crítica de diferentes métodos para restaurar el volumen y la masa de animales extintos" (PDF) . Anales del Museo Carnegie . 85 (4): 335–358. doi :10.2992/007.085.0403. S2CID 210840060.
^ Pal, Saurabh; Ayyasami, Krishnan (27 de junio de 2022). "El titán perdido de Cauvery". Geología hoy . 38 (3): 112-116. Código Bib : 2022GeolT..38..112P. doi :10.1111/gto.12390. ISSN 0266-6979. S2CID 250056201.
^ Pablo, Gregorio S.; Larramendi, Asier (11 de abril de 2023). "La estimación de la masa corporal del Bruhathkayosaurus y otros restos fragmentarios de saurópodos sugiere que los animales terrestres más grandes eran aproximadamente tan grandes como las ballenas más grandes". Lethaia . 56 (2): 1–11. Código Bib : 2023 Letha..56..2.5P. doi :10.18261/let.56.2.5. ISSN 0024-1164. S2CID 259782734.
^ Dal Sasso, Cristiano; Magánuco, Simone; Buffetaut, Éric; et al. (2005). "Nueva información sobre el cráneo del enigmático terópodo Spinosaurus, con comentarios sobre sus tamaños y afinidades" (PDF) . Revista de Paleontología de Vertebrados . Milton Park, Oxfordshire: Taylor & Francis para la Sociedad de Paleontología de Vertebrados. 25 (4): 888–896. doi :10.1671/0272-4634(2005)025[0888:NIOTSO]2.0.CO;2. ISSN 0272-4634. S2CID 85702490. Archivado desde el original (PDF) el 29 de abril de 2011 . Consultado el 5 de mayo de 2011 .
^ ab Therrien, François; Henderson, Donald M. (2007). "Mi terópodo es más grande que el tuyo... o no: estimar el tamaño corporal a partir de la longitud del cráneo en los terópodos". Revista de Paleontología de Vertebrados . Milton Park, Oxfordshire: Taylor & Francis para la Sociedad de Paleontología de Vertebrados. 27 (1): 108-115. doi :10.1671/0272-4634(2007)27[108:MTIBTY]2.0.CO;2. ISSN 0272-4634. S2CID 86025320.
^ Zhao, Xijin ; Li, Dunjing; Han, pandilla; et al. (2007). "Zhuchengosaurus maximus de la provincia de Shandong". Acta Geoscientia Sínica . Beijing : Academia China de Ciencias Geológicas . 28 (2): 111-122. ISSN 1006-3021.
^ Weishampel, Dodson y Osmólska 2004, págs. 438–463, cap. 20: "Hadrosauridae" de John R. Horner David B. Weishampel y Catherine A. Forster.
^ Norell, Gaffney y Dingus 2000
^ "Abeja Colibrí (Mellisuga helenae)". Aves.com . Medios Paley. Archivado desde el original el 3 de abril de 2015 . Consultado el 27 de octubre de 2019 .
^ ab Zhang, Fucheng; Zhou, Zhonghe ; Xu, Xing ; et al. (2008). "Un extraño maniraptoran del Jurásico de China con plumas alargadas en forma de cinta". Naturaleza . Londres: Investigación de la naturaleza. 455 (7216): 1105–1108. Código Bib : 2008Natur.455.1105Z. doi : 10.1038/naturaleza07447. ISSN 0028-0836. PMID 18948955. S2CID 4362560.
^ ab Xu, Xing; Zhao, Qi; Norell, Marcos; et al. (febrero de 2008). "Un nuevo fósil de dinosaurio maniraptorano emplumado que llena un vacío morfológico en el origen aviar". Boletín de ciencia chino . Ámsterdam: Elsevier en nombre de Science in China Press. 54 (3): 430–435. doi : 10.1007/s11434-009-0009-6 . ISSN 1001-6538. S2CID 53445386.
^ Holtz 2007
^ Mayordomo, Richard J.; Zhao, Qi (febrero de 2009). "Los dinosaurios ornitisquios de cuerpo pequeño Micropachycephalosaurus hongtuyanensis y Wannanosaurus yansiensis del Cretácico Superior de China". Investigación del Cretácico . Ámsterdam: Elsevier. 30 (1): 63–77. Código Bib : 2009CrRes..30...63B. doi :10.1016/j.cretres.2008.03.002. ISSN 0195-6671.
^ Yans, Johan; Dejax, Jean; Pons, Denise; et al. (enero-febrero de 2005). "Implications paléontologiques et géodynamiques de la datation palynologique des sédiments à faciès wealdien de Bernissart (bassin de Mons, Belgique)" [Implicaciones paleontológicas y geodinámicas de la datación palinológica de los sedimentos de facies wealden de Bernissart (Cuenca de Mons, Bélgica)]. Comptes Rendus Palevol (en francés). Amsterdam: Elsevier en nombre de la Academia de Ciencias de Francia. 4 (1–2): 135–150. Código Bib : 2005CRPal...4..135Y. doi :10.1016/j.crpv.2004.12.003. ISSN 1631-0683.
^ Día, Julia J.; Upchurch, Pablo; Norman, David B.; et al. (2002). "Huellas, evolución y comportamiento de los saurópodos" (PDF) . Ciencia . Washington, DC: Asociación Estadounidense para el Avance de la Ciencia. 296 (5573): 1659. doi :10.1126/science.1070167. ISSN 0036-8075. PMID 12040187. S2CID 36530770.
^ Curry Rogers y Wilson 2005, págs. 252–284, cap. 9: "Pasos para comprender la biología de los saurópodos: la importancia de las huellas de los saurópodos" por Joanna L. Wright.
^ Varricchio, David J.; Sereno, Paul C.; Zhao, Xijin; et al. (2008). "La manada atrapada en el barro captura evidencia de la sociabilidad distintiva de los dinosaurios" (PDF) . Acta Paleontológica Polonica . Varsovia : Instituto de Paleobiología, Academia Polaca de Ciencias . 53 (4): 567–578. doi : 10.4202/app.2008.0402. ISSN 0567-7920. S2CID 21736244. Archivado (PDF) desde el original el 30 de marzo de 2019 . Consultado el 6 de mayo de 2011 .
^ Lessem & Glut 1993, págs. 19-20, "Alosaurio"
^ Maxwell, W. Desmond; Ostrom, John H. (1995). "Tafonomía e implicaciones paleobiológicas de las asociaciones Tenontosaurus - Deinonychus ". Revista de Paleontología de Vertebrados . Milton Park, Oxfordshire: Taylor & Francis para la Sociedad de Paleontología de Vertebrados. 15 (4): 707–712. Código Bib : 1995JVPal..15..707M. doi :10.1080/02724634.1995.10011256. ISSN 0272-4634.
^ Cucaracha, Brian T.; Brinkman, Daniel L. (abril de 2007). "Una reevaluación de la caza cooperativa en manada y el gregarismo en Deinonychus antirrhopus y otros dinosaurios terópodos no aviares". Boletín del Museo Peabody de Historia Natural . New Haven, CT : Museo Peabody de Historia Natural . 48 (1): 103-138. doi :10.3374/0079-032X(2007)48[103:AROCPH]2.0.CO;2. ISSN 0079-032X. S2CID 84175628.
^ Tanke, Darren H. (1998). "Comportamiento de morderse la cabeza en dinosaurios terópodos: evidencia paleopatológica" (PDF) . Gaia: Revista de Geociencias . Lisboa : Museo Nacional de Historia Natural y Ciencia (15): 167–184. doi :10.7939/R34T6FJ1P. ISSN 0871-5424. S2CID 90552600. Archivado desde el original (PDF) el 27 de febrero de 2008.
^ "Los dinosaurios que luchan". Nueva York: Museo Americano de Historia Natural. Archivado desde el original el 18 de enero de 2012 . Consultado el 5 de diciembre de 2007 .
^ ab Carpenter, Kenneth (1998). "Evidencia de comportamiento depredador por parte de dinosaurios terópodos" (PDF) . Gaia: Revista de Geociencias . Lisboa: Museo Nacional de Historia Natural y Ciencia. 15 : 135-144. ISSN 0871-5424. Archivado (PDF) desde el original el 26 de septiembre de 2013.
^ Rogers, Raymond R.; Krause, David W .; Curry Rogers, Kristina (2007). "Canibalismo en el dinosaurio malgache Majungatholus atopus ". Naturaleza . Londres: Investigación de la naturaleza. 422 (6931): 515–518. Código Bib :2003Natur.422..515R. doi : 10.1038/naturaleza01532. ISSN 0028-0836. PMID 12673249. S2CID 4389583.
^ Schmitz, Lars; Motani, Ryosuke (2011). "La nocturnidad en los dinosaurios se infiere de la morfología de la órbita y el anillo escleral". Ciencia . Washington, DC: Asociación Estadounidense para el Avance de la Ciencia. 332 (6030): 705–708. Código Bib : 2011 Ciencia... 332..705S. doi :10.1126/ciencia.1200043. ISSN 0036-8075. PMID 21493820. S2CID 33253407.
^ Varricchio, David J.; Martín, Antonio J .; Katsura, Yoshihiro (2007). "Primer rastro y evidencia fósil corporal de un dinosaurio excavador y madriguera". Actas de la Royal Society B. Londres: Sociedad de la Realeza. 274 (1616): 1361-1368. doi :10.1098/rspb.2006.0443. ISSN 0962-8452. PMC 2176205 . PMID 17374596.
^ Chiappe y Witmer 2002
^ Chatterjee, Sankar ; Templin, R. Jack (2007). "Forma en planta del ala de biplano y rendimiento de vuelo del dinosaurio emplumado Microraptor gui" (PDF) . Proc. Nacional. Acad. Ciencia. EE.UU . Washington, DC: Academia Nacional de Ciencias. 104 (5): 1576-1580. Código bibliográfico : 2007PNAS..104.1576C. doi : 10.1073/pnas.0609975104 . ISSN 0027-8424. PMC 1780066 . PMID 17242354. Archivado (PDF) desde el original el 18 de agosto de 2019 . Consultado el 29 de octubre de 2019 .
^ Henderson, Donald M. (2003). "Efectos de los cálculos estomacales sobre la flotabilidad y el equilibrio de un cocodrilo flotante: un análisis computacional". Revista Canadiense de Zoología . Ottawa : Prensa de investigación de la NRC . 81 (8): 1346-1357. doi :10.1139/z03-122. ISSN 0008-4301.
^ a b Senter, Phil (2008). "Voces del pasado: una revisión de los sonidos de animales del Paleozoico y Mesozoico". Biología histórica . Milton Park, Oxfordshire: Taylor y Francis. 20 (4): 255–287. Código Bib : 2008HBio...20..255S. doi : 10.1080/08912960903033327 . ISSN 0891-2963. S2CID 84473967.
^ Li, Quanguo; Gao, Ke-Qin; Vinther, Jakob; et al. (2010). "Patrones de color del plumaje de un dinosaurio extinto" (PDF) . Ciencia . Washington, DC: Asociación Estadounidense para el Avance de la Ciencia. 327 (5971): 1369-1372. Código Bib : 2010 Ciencia... 327.1369L. doi : 10.1126/ciencia.1186290. ISSN 0036-8075. PMID 20133521. S2CID 206525132. Archivado (PDF) desde el original el 30 de marzo de 2019 . Consultado el 7 de noviembre de 2019 .
^ Riede, T. (2019). "La evolución de la siringe: una teoría acústica". MÁS UNO . 17 (2): e2006507. doi : 10.1371/journal.pbio.2006507 . PMC 6366696 . PMID 30730882.
^ Clarke, Julia A .; Chatterjee, Sankar; Zhiheng, Li; et al. (2016). "Evidencia fósil del órgano vocal aviar del Mesozoico". Naturaleza . Londres: Investigación de la naturaleza. 538 (7626): 502–505. Código Bib :2016Natur.538..502C. doi : 10.1038/naturaleza19852. ISSN 0028-0836. PMID 27732575. S2CID 4389926.
^ Kingsley, EP; et al. (2018). "Identidad y novedad en la siringe aviar". Actas de la Academia Nacional de Ciencias de los Estados Unidos de América . 115 (41): 10109–10217. Código Bib : 2018PNAS..11510209K. doi : 10.1073/pnas.1804586115 . PMC 6187200 . PMID 30249637.
^ Yoshida, adicto; Kobayashi, Yoshitsugu; Norell, Mark A. (15 de febrero de 2023). "La laringe de un anquilosaurio proporciona información sobre la vocalización similar a la de un pájaro en dinosaurios no aviares". Biología de las Comunicaciones . 6 (1): 152. doi :10.1038/s42003-023-04513-x. ISSN 2399-3642. PMC 9932143 . PMID 36792659.
^ Riede, Tobías; Eliason, Chad M.; Molinero, Edward H.; et al. (2016). "Arrullos, booms y ululares: la evolución del comportamiento vocal con la boca cerrada en las aves". Evolución . Hoboken, Nueva Jersey: John Wiley & Sons para la Sociedad para el Estudio de la Evolución . 70 (8): 1734-1746. doi : 10.1111/evo.12988 . ISSN 0014-3820. PMID 27345722. S2CID 11986423.
^ Weishampel, David B. (primavera de 1981). "Análisis acústico de la vocalización de dinosaurios lambeosaurinos (Reptilia: Ornithischia)" (PDF) . Paleobiología . Bethesda, MD : Sociedad Paleontológica . 7 (2): 252–261. doi :10.1017/S0094837300004036. ISSN 0094-8373. JSTOR 2400478. S2CID 89109302. Archivado desde el original (PDF) el 6 de octubre de 2014 . Consultado el 30 de octubre de 2019 .
^ Miyashita, Tetsuto; Cenador, Victoria M .; Witmer, Lawrence M.; et al. (Diciembre de 2011). "La morfología craneal interna de un dinosaurio blindado Euoplocephalus corroborada por reconstrucción por tomografía computarizada con rayos X" (PDF) . Revista de Anatomía . Hoboken, Nueva Jersey: John Wiley & Sons. 219 (6): 661–675. doi :10.1111/j.1469-7580.2011.01427.x. ISSN 1469-7580. PMC 3237876 . PMID 21954840. Archivado desde el original (PDF) el 24 de septiembre de 2015 . Consultado el 30 de octubre de 2019 .
^ Hansell 2000
^ ab Varricchio, David J.; Horner, John R.; Jackson, Frankie D. (2002). "Embriones y huevos del dinosaurio terópodo del Cretácico Troodon formosus ". Revista de Paleontología de Vertebrados . Milton Park, Oxfordshire: Taylor & Francis para la Sociedad de Paleontología de Vertebrados. 22 (3): 564–576. doi :10.1671/0272-4634(2002)022[0564:EAEFTC]2.0.CO;2. ISSN 0272-4634. S2CID 85728452.
^ Lee, Andrew H.; Werning, Sarah (2008). "La madurez sexual en los dinosaurios en crecimiento no se ajusta a los modelos de crecimiento de los reptiles". Proc. Nacional. Acad. Ciencia. EE.UU . Washington, DC: Academia Nacional de Ciencias. 105 (2): 582–587. Código Bib : 2008PNAS..105..582L. doi : 10.1073/pnas.0708903105 . ISSN 0027-8424. PMC 2206579 . PMID 18195356.
^ Horner, John R.; Makela, Robert (1979). "El nido de juveniles proporciona evidencia de la estructura familiar entre los dinosaurios". Naturaleza . Londres: Investigación de la naturaleza. 282 (5736): 296–298. Código Bib :1979Natur.282..296H. doi :10.1038/282296a0. ISSN 0028-0836. S2CID 4370793.
^ "Descubriendo el comportamiento de los dinosaurios: vista desde 1960 hasta el presente". Enciclopedia Británica . Chicago, IL : Encyclopædia Britannica, Inc. Archivado desde el original el 13 de diciembre de 2013 . Consultado el 30 de octubre de 2019 .
^ Currie y col. 2004, págs. 234–250, cap. 11: "El comportamiento de incubación de los dinosaurios y el origen de las plumas de vuelo" por Thomas P. Hopp y Mark J. Orsen.
^ Reisz, Robert R .; Scott, Diana; Sues, Hans-Dieter ; et al. (2005). "Embriones de un dinosaurio prosaurópodo del Jurásico temprano y su importancia evolutiva" (PDF) . Ciencia . Washington, DC: Asociación Estadounidense para el Avance de la Ciencia. 309 (5735): 761–764. Código Bib : 2005 Ciencia... 309..761R. doi : 10.1126/ciencia.1114942. ISSN 0036-8075. PMID 16051793. S2CID 37548361. Archivado (PDF) desde el original el 22 de julio de 2018.
^ Clark, Neil DL; Cabina, Pablo; Stand, Claire L.; et al. (2004). "Huellas de dinosaurios de la Formación Duntulm (Bathoniano, Jurásico) de la Isla de Skye" (PDF) . Revista Escocesa de Geología . Londres: Sociedad Geológica de Londres . 40 (1): 13-21. Código Bib : 2004ScJG...40...13C. doi :10.1144/sjg40010013. ISSN 0036-9276. S2CID 128544813. Archivado (PDF) desde el original el 22 de julio de 2013 . Consultado el 12 de diciembre de 2019 .
^ Zhou, Zhonghe; Zhang, Fucheng (2004). "Un embrión de ave precocial del Cretácico Inferior de China". Ciencia . Washington, DC: Asociación Estadounidense para el Avance de la Ciencia. 306 (5696): 653. doi :10.1126/ciencia.1100000. ISSN 0036-8075. PMID 15499011. S2CID 34504916.
^ Naish, Darren (15 de mayo de 2012). "Una colonia de anidación ahogada de aves del Cretácico Superior". Ciencia . Científico americano . 306 (5696): 653. doi :10.1126/ciencia.1100000. PMID 15499011. S2CID 34504916. Archivado desde el original el 25 de septiembre de 2018 . Consultado el 16 de noviembre de 2019 .
^ Fernández, Mariela S.; García, Rodolfo A.; Fiorelli, Lucas; et al. (2013). "Una gran acumulación de huevos de aves del Cretácico Superior de la Patagonia (Argentina) revela una novedosa estrategia de anidación en aves mesozoicas". MÁS UNO . San Francisco, CA: MÁS. 8 (4): e61030. Código Bib : 2013PLoSO...861030F. doi : 10.1371/journal.pone.0061030 . ISSN 1932-6203. PMC 3629076 . PMID 23613776.
^ Considerando, Denis Charles; Mayr, Gerald (mayo de 2018). "La morfología de la pelvis sugiere que las aves del Mesozoico temprano eran demasiado pesadas para tener contacto e incubar sus huevos" (PDF) . Revista de biología evolutiva . Hoboken, Nueva Jersey: Wiley-Blackwell en nombre de la Sociedad Europea de Biología Evolutiva . 31 (5): 701–709. doi :10.1111/jeb.13256. ISSN 1010-061X. PMID 29485191. S2CID 3588317. Archivado (PDF) desde el original el 2 de junio de 2020.
^ Myers, Timothy S.; Fiorillo, Anthony R. (2009). "Evidencia de comportamiento gregario y segregación por edades en dinosaurios saurópodos" (PDF) . Paleogeografía, Paleoclimatología, Paleoecología . Ámsterdam: Elsevier. 274 (1–2): 96–104. Código Bib : 2009PPP...274...96M. doi :10.1016/j.palaeo.2009.01.002. ISSN 0031-0182. Archivado (PDF) desde el original el 29 de mayo de 2020.
^ Vinther, Jakob; Nicholls, Robert; Kelly, Diane A. (22 de febrero de 2021). "Una abertura cloacal en un dinosaurio no aviar". Biología actual . Elsevier. 31 (4): R1–R3. doi : 10.1016/j.cub.2020.12.039 . PMID 33472049. S2CID 231644183.
^ Weishampel, Dodson y Osmólska 2004, págs. 643–659, cap. 28: "Fisiología de los dinosaurios no avianos" de Anusuya Chinsamy y Willem J. Hillenius.
^ Pontzer, H.; Allen, V.; Hutchinson, JR (2009). "La biomecánica de la carrera indica endotermia en dinosaurios bípedos". MÁS UNO . 4 (11): e7783. Código Bib : 2009PLoSO...4.7783P. doi : 10.1371/journal.pone.0007783 . ISSN 1932-6203. PMC 2772121 . PMID 19911059.
^ ab Benson, RBJ (2018). "Macroevolución y macroecología de dinosaurios". Revisión anual de ecología, evolución y sistemática . 49 : 379–408. doi : 10.1146/annurev-ecolsys-110617-062231 . S2CID 92837486.
^ Grady, JM; Enquist, BJ; Dettweiler-Robinson, E.; Wright, NA; Smith, FA (2014). "Evidencia de mesotermia en dinosaurios". Ciencia . 344 (6189): 1268-1272. Código Bib : 2014 Ciencia... 344.1268G. doi : 10.1126/ciencia.1253143. PMID 24926017. S2CID 9806780.
^ Legendre, LJ; Guénard, G.; Botha-Brink, J.; Cubo, J. (2016). "Evidencia paleohistológica de una alta tasa metabólica ancestral en arcosaurios". Biología Sistemática . 65 (6): 989–996. doi : 10.1093/sysbio/syw033 . PMID 27073251.
^ Seymour, RS; Bennett-Stamper, CL; Johnston, SD; Transportista, República Dominicana; Grigg, GC (2004). "Evidencia de ancestros endotérmicos de los cocodrilos en el origen de la evolución de los arcosaurios". Zoología Fisiológica y Bioquímica . 77 (6): 1051–1067. doi : 10.1093/sysbio/syw033 . PMID 27073251.
^ Parsons 2001, págs. 22-48, "Las herejías del Dr. Bakker".
^ Erickson, gerente general (2014). "Sobre el crecimiento de los dinosaurios". Revista Anual de Ciencias de la Tierra y Planetarias . 42 (1): 675–697. Código Bib : 2014AREPS..42..675E. doi : 10.1146/annurev-earth-060313-054858.
^ abBailleul , AM; O'Connor, J.; Schweitzer, MH (2019). "Paleohistología de dinosaurios: revisión, tendencias y nuevas vías de investigación". PeerJ . 7 : e7764. doi : 10.7717/peerj.7764 . PMC 6768056 . PMID 31579624.
^ De Ricqlès, A. (1974). «Evolución de la endotermia: evidencia histológica» (PDF) . Teoría Evolutiva . 1 (2): 51–80. Archivado (PDF) desde el original el 17 de abril de 2021.
^ De Ricqlès, A. (1980). "Estructuras tisulares del hueso de dinosaurio, significado funcional y posible relación con la fisiología de los dinosaurios". En Thomas, RDK; Olson, CE (eds.). Una mirada fría a los dinosaurios de sangre caliente . Nueva York: Asociación Estadounidense para el Avance de la Ciencia. págs. 103-139.
^ Padian, K.; Horner, JR; de Ricqlès, A. (2004). "Crecimiento de pequeños dinosaurios y pterosaurios: la evolución de las estrategias de crecimiento de los arcosaurios" (PDF) . Revista de Paleontología de Vertebrados . 24 (3): 555–571. doi :10.1671/0272-4634(2004)024[0555:GISDAP]2.0.CO;2. S2CID 86019906.
^ de Souza, GA; Bento Soares, M.; Souza Brum, A.; Zucolotto, M.; Sayão, JM; Carlos Weinschütz, L.; Kellner, AWA (2020). "Osteohistología y dinámica de crecimiento del noasáurido brasileño Vespersaurus paranaensis Langer et al., 2019 (Theropoda: Abelisauroidea)". PeerJ . 8 : e9771. doi : 10.7717/peerj.9771 . PMC 7500327 . PMID 32983636. S2CID 221906765.
^ Para ver ejemplos de este trabajo realizado en diferentes linajes de dinosaurios, consulte
Erickson, gerente general; Tumanova, TA (2000). "Curva de crecimiento de Psittacosaurus mongoliensis Osborn (Ceratopsia: Psittacosauridae) inferida de la histología de huesos largos". Revista zoológica de la Sociedad Linneana . 130 (4): 551–566. doi :10.1111/j.1096-3642.2000.tb02201.x. S2CID 84241148.
Erickson, G.; Rogers, K.; Yerby, S. (2001). "Patrones de crecimiento de dinosaurios y rápidas tasas de crecimiento de aves". Naturaleza . 412 (429–433): 429–433. Código Bib :2001Natur.412..429E. doi :10.1038/35086558. PMID 11473315. S2CID 4319534.
Erickson, G.; Makovický, P.; Currie, P.; Norell, MA; Yerby, SA; Brochu, CA (2004). "Gigantismo y parámetros comparativos de la historia de vida de los dinosaurios tiranosáuridos" (PDF) . Naturaleza . 430 (7001): 772–775. Código Bib :2004Natur.430..772E. doi : 10.1038/naturaleza02699. PMID 15306807. S2CID 4404887. Archivado (PDF) desde el original el 14 de julio de 2020.
Lehman, TM; Woodward, HN (2008). "Modelado de tasas de crecimiento de dinosaurios saurópodos" (PDF) . Paleobiología . 34 (2): 264–281. doi :10.1666/0094-8373(2008)034[0264:MGRFSD]2.0.CO;2. S2CID 84163725.
Horner, JR; de Ricqles, A.; Padián, K.; Scheetz, RD (2009). "Histología comparativa de huesos largos y crecimiento de los dinosaurios" hipsilofdóntidos " Orodromeus makelai , Dryosaurus altus y Tenontosaurus illettii (Ornithischia: Euornithopoda)". Revista de Paleontología de Vertebrados . 29 (3): 734–747. Código Bib : 2009JVPal..29..734H. doi :10.1671/039.029.0312. S2CID 86277619.
Woodward, H.; Freedman Fowler, E.; Farlow, J.; Horner, J. (2015). " Maiasaura , un organismo modelo para la biología de poblaciones de vertebrados extintos: una evaluación estadística de muestra grande de la dinámica de crecimiento y la supervivencia". Paleobiología . 41 (4): 503–527. Código Bib : 2015Pbio...41..503W. doi :10.1017/pab.2015.19. S2CID 85902880.
^ Amiot, R.; Lecuyer, C.; Buffetaut, E.; Escarguel, G.; Fluteau, F.; Martineau, F. (2006). "Los isótopos de oxígeno de las apatitas biogénicas sugieren una endotermia generalizada en los dinosaurios del Cretácico" (PDF) . Cartas sobre ciencias planetarias y de la Tierra . 246 (1–2): 41–54. Código Bib : 2006E y PSL.246...41A. doi :10.1016/j.epsl.2006.04.018.
^ Amiot, R.; Wang, X.; Lecuyer, C.; Buffetaut, E.; Boudad, L.; Cavin, L.; Ding, Z.; Fluteau, F.; Kellner, AWA; Tong, H.; Zhang, F. (2010). "Composiciones de isótopos de oxígeno y carbono de vertebrados del Cretácico medio del norte de África y Brasil: importancia ecológica y ambiental". Paleogeografía, Paleoclimatología, Paleoecología . 297 (2): 439–451. Código Bib : 2010PPP...297..439A. doi :10.1016/j.palaeo.2010.08.027.
^ Kolodny, Y.; Luz, B.; Sander, M.; Clemens, WA (1996). "Huesos de dinosaurio: ¿fósiles o pseudomorfos? Los peligros de la reconstrucción fisiológica a partir de fósiles apatiticos" (PDF) . Paleogeografía, Paleoclimatología, Paleoecología . 126 (1–2): 161–171. Código Bib : 1996PPP...126..161K. doi :10.1016/S0031-0182(96)00112-5.
^ Pablo, GS (1988). "Implicaciones fisiológicas, migratorias, climatológicas, geofísicas, de supervivencia y evolutivas de los dinosaurios polares del Cretácico". Revista de Paleontología . 62 (4): 640–652. JSTOR 1305468.
^ Clemens, Washington; Nelms, LG (1993). "Implicaciones paleoecológicas de la fauna de vertebrados terrestres de Alaska en el Cretácico posterior en paleolatitudes altas". Geología . 21 (6): 503–506. Código Bib : 1993Geo....21..503C. doi :10.1130/0091-7613(1993)021<0503:PIOATV>2.3.CO;2.
^ Buffetaut, E. (2004). "Los dinosaurios polares y la cuestión de la extinción de los dinosaurios: una breve reseña" (PDF) . Paleogeografía, Paleoclimatología, Paleoecología . 214 (3): 225–231. doi :10.1016/j.palaeo.2004.02.050. Archivado (PDF) desde el original el 8 de junio de 2020.
^ ab Sereno, Paul C.; Martínez, Ricardo N.; Wilson, Jeffrey A.; et al. (Septiembre de 2008). Kemp, Tom (ed.). "Evidencia de sacos aéreos intratorácicos aviares en un nuevo dinosaurio depredador de Argentina". MÁS UNO . San Francisco, CA: MÁS. 3 (9): e3303. Código Bib : 2008PLoSO...3.3303S. doi : 10.1371/journal.pone.0003303 . ISSN 1932-6203. PMC 2553519 . PMID 18825273.
^ O'Connor, PM (2009). "Evolución de los planes corporales de los arcosaurios: adaptaciones esqueléticas de un aparato respiratorio basado en sacos de aire en aves y otros arcosaurios". Revista de Zoología Experimental Parte A: Genética y Fisiología Ecológica . 311 (8): 629–646. doi :10.1002/jez.548. PMID 19492308.
^ Águila, RA; Tütken, T.; Martín, TS; Tripati, Alaska; Fricke, HC; Connely, M.; Cifelli, RL; Eiler, JM (2011). "La temperatura corporal de los dinosaurios se determina a partir del ordenamiento isotópico ( 13 C - 18 O) en biominerales fósiles". Ciencia . 333 (6041): 443–445. Código Bib : 2011 Ciencia... 333.. 443E. doi : 10.1126/ciencia.1206196. PMID 21700837. S2CID 206534244.
^ Wedel, MJ (2003). "Neumática vertebral, sacos aéreos y fisiología de los dinosaurios saurópodos" (PDF) . Paleobiología . 29 (2): 243–255. Código Bib : 2003Pbio...29..243W. doi :10.1017/S0094837300018091.
^ Perry, SF; Cristiano, A.; Breuer, T.; Pajor, N.; Codd, JR (2009). "Implicaciones de un sistema respiratorio de estilo aviar para el gigantismo en dinosaurios saurópodos". Revista de Zoología Experimental Parte A: Genética y Fisiología Ecológica . 311 (8): 600–610. Código Bib : 2009JEZA..311..600P. doi :10.1002/jez.517. PMID 19189317.
^ Alejandro, RM (1998). "Gigantes de todos los tiempos: los animales más grandes y sus problemas". Paleontología . 41 : 1231-1245.
^ Tsahar, E.; Martínez del Río, C.; Izhaki, I.; Arad, Z. (2005). "¿Pueden las aves ser amonotélicas? Equilibrio y excreción de nitrógeno en dos frugívoros" (PDF) . La Revista de Biología Experimental . 208 (6): 1025-1034. doi :10.1242/jeb.01495. ISSN 0022-0949. PMID 15767304. S2CID 18540594. Archivado (PDF) desde el original el 17 de octubre de 2019 . Consultado el 31 de octubre de 2019 .
^ Skadhauge, E.; Erlwanger, KH; Ruziwa, SD; Dantzer, V.; Elbrønd, VS; Chamunorwa, JP (2003). "¿El coprodeum del avestruz ( Struthio camelus ) tiene las propiedades electrofisiológicas y la microestructura de otras aves?". Bioquímica y fisiología comparadas, Parte A: Fisiología molecular e integrativa . 134 (4): 749–755. doi :10.1016/S1095-6433(03)00006-0. ISSN 1095-6433. PMID 12814783.
^ Preest, señor; Beuchat, CA (1997). "Excreción de amoníaco por los colibríes". Naturaleza . 386 (6625): 561–562. Código Bib :1997Natur.386..561P. doi :10.1038/386561a0. ISSN 0028-0836. S2CID 4372695.
^ Mora, J.; Martuscelli, J.; Ortíz Pineda, J.; Soberón, G. (1965). "La regulación de las enzimas de biosíntesis de urea en vertebrados". Revista Bioquímica . 96 (1): 28–35. doi :10.1042/bj0960028. ISSN 0264-6021. PMC 1206904 . PMID 14343146.
^ Packard, GC (1966). "La influencia de la temperatura ambiente y la aridez en los modos de reproducción y excreción de los vertebrados amniotas". El naturalista americano . 100 (916): 667–682. doi :10.1086/282459. ISSN 0003-0147. JSTOR 2459303. S2CID 85424175.
^ Balgooyen, TG (1971). "Regurgitación de pellets por gavilán cautivo (Falco sparverius)" (PDF) . Cóndor . 73 (3): 382–385. doi :10.2307/1365774. JSTOR 1365774. Archivado desde el original (PDF) el 4 de abril de 2019 . Consultado el 30 de octubre de 2019 .
^ Xu, X.; Li, F.; Wang, Y.; Sullivan, C.; Zhang, F.; Zhang, X.; Sullivan, C.; Wang, X.; Zheng, X. (2018). "Excepcionales fósiles de dinosaurios revelan el origen temprano de la digestión al estilo aviar". Informes científicos . 8 (1): 14217. Código bibliográfico : 2018NatSR...814217Z. doi :10.1038/s41598-018-32202-x. ISSN 2045-2322. PMC 6155034 . PMID 30242170.
^ Russell, Dale A. (1997). "Inteligencia". En Kevin Padián; Philip J. Currie (eds.). Enciclopedia de dinosaurios . San Diego: Prensa académica. págs. 370–372. ISBN978-0-12-226810-6.
^ Osborn, Henry Fairfield (1924). "Tres nuevos Theropoda, zona de Protoceratops, Mongolia central" (PDF) . Novitates del Museo Americano . Nueva York: Museo Americano de Historia Natural (144): 1–12. ISSN 0003-0082. Archivado (PDF) desde el original el 12 de junio de 2007.
^ Ostrom, John H. (1973). "La ascendencia de las aves". Naturaleza . Londres: Investigación de la naturaleza. 242 (5393): 136. Código bibliográfico : 1973NPhS..242..136O. doi : 10.1038/242136a0 . ISSN 0028-0836. S2CID 29873831.
^ Padian 1986, págs. 1–55, "La monofilia saurisquiana y el origen de las aves" de Jacques Gauthier .
^ Mayr, Gerald; Pohl, Burkhard; Peters, D. Stefan (2005). "Un espécimen de Archaeopteryx bien conservado con características de terópodo" (PDF) . Ciencia . Washington, DC: Asociación Estadounidense para el Avance de la Ciencia. 310 (5753): 1483–1486. Código Bib : 2005 Ciencia... 310.1483M. doi : 10.1126/ciencia.1120331. ISSN 0036-8075. PMID 16322455. S2CID 28611454.
^ ab Feduccia, Alan (1 de octubre de 2002). "Los pájaros son dinosaurios: una respuesta simple a un problema complejo". El alca . Washington, DC: Unión Estadounidense de Ornitólogos . 119 (4): 1187-1201. doi :10.1642/0004-8038(2002)119[1187:BADSAT]2.0.CO;2. ISSN 0004-8038. JSTOR 4090252. S2CID 86096746 . Consultado el 3 de noviembre de 2019 .
^ ab Switek, Brian (2 de julio de 2012). "El ascenso de los dinosaurios peludos". Noticias. Naturaleza . Londres: Investigación de la naturaleza. doi :10.1038/naturaleza.2012.10933. ISSN 0028-0836. S2CID 123219913 . Consultado el 1 de enero de 2019 .
^ Godefroit, P.; Sinitsa, SM; Dhouailly, D.; Bolotsky, YL; Sizov, AV; McNamara, ME; Benton, MJ; España, P. (2014). "Un dinosaurio ornitisquio del Jurásico de Siberia con plumas y escamas" (PDF) . Ciencia . 345 (6195): 451–455. Código Bib : 2014 Ciencia... 345.. 451G. doi : 10.1126/ciencia.1253351. hdl :1983/a7ae6dfb-55bf-4ca4-bd8b-a5ea5f323103. PMID 25061209. S2CID 206556907. Archivado desde el original (PDF) el 9 de febrero de 2019 . Consultado el 27 de julio de 2016 .
^ Xu, Xing; Norell, Mark A.; Kuang, Xuewen; et al. (2004). "Tirannosáuridos basales de China y evidencia de protoplumas en tiranosáuridos" (PDF) . Naturaleza . Londres: Investigación de la naturaleza. 431 (7009): 680–684. Código Bib :2004Natur.431..680X. doi : 10.1038/naturaleza02855. ISSN 0028-0836. PMID 15470426. S2CID 4381777.
^ Göhlich, Úrsula B.; Chiappe, Luis M. (2006). "Un nuevo dinosaurio carnívoro del archipiélago de Solnhofen del Jurásico tardío" (PDF) . Naturaleza . Londres: Investigación de la naturaleza. 440 (7082): 329–332. Código Bib :2006Natur.440..329G. doi : 10.1038/naturaleza04579. ISSN 0028-0836. PMID 16541071. S2CID 4427002. Archivado desde el original (PDF) el 26 de abril de 2019 . Consultado el 1 de noviembre de 2019 .
^ Kellner, Alejandro WA ; Wang, Xiaolin; Tischlinger, Helmut; et al. (2010). "El tejido blando de Jeholopterus (Pterosauria, Anurognathidae, Batrachognathinae) y la estructura de la membrana del ala del pterosaurio". Actas de la Royal Society B. Londres: Sociedad de la Realeza. 277 (1679): 321–329. doi :10.1098/rspb.2009.0846. ISSN 0962-8452. PMC 2842671 . PMID 19656798.
^ Mayr, G.; Pittman, M.; Saitta, E.; Kaye, TG; Vinther, J. (2016). "Estructura y homología de las cerdas de la cola de Psittacosaurus". Paleontología . 59 (6): 793–802. Código Bib : 2016Palgy..59..793M. doi :10.1111/pala.12257. hdl : 1983/029c668f-08b9-45f6-a0c5-30ce9256e593 . S2CID 89156313.
^ ab Benton, MJ; Dhouailly, D.; Jiang, B.; McNamara, M. (2019). "El origen temprano de las plumas". Tendencias en ecología y evolución . 34 (9): 856–869. doi : 10.1016/j.tree.2019.04.018. hdl : 10468/8068 . PMID 31164250. S2CID 174811556.
^ Barrett, primer ministro; Evans, CC; Campione, NE (2015). "Evolución de las estructuras epidérmicas de los dinosaurios". Cartas de biología . 11 (6): 20150229. doi :10.1098/rsbl.2015.0229. PMC 4528472 . PMID 26041865.
^ Alibardi, Lorenzo; Knapp, Loren W.; Sawyer, Roger H. (2006). "Localización de beta-queratina en el desarrollo de escamas y plumas de caimán en relación con el desarrollo y evolución de las plumas". Revista de Citología y Patología Submicroscópica . Siena : Nuova Immagine Editrice. 38 (2–3): 175–192. ISSN 1122-9497. PMID 17784647.
^ Lingham-Soliar, Theagarten (diciembre de 2003). "El origen dinosaurio de las plumas: perspectivas de las fibras de colágeno de los delfines (Cetacea)". Naturwissenschaften . Berlín: Springer Science+Business Media . 90 (12): 563–567. Código Bib : 2003NW..... 90.. 563L. doi :10.1007/s00114-003-0483-7. ISSN 0028-1042. PMID 14676953. S2CID 43677545.
^ ab Feduccia, Alan; Lingham-Soliar, Theagarten; Hinchliffe, J. Richard (noviembre de 2005). "¿Existen los dinosaurios emplumados? Probando la hipótesis sobre evidencia neontológica y paleontológica". Revista de Morfología . Hoboken, Nueva Jersey: John Wiley & Sons. 266 (2): 125–166. doi :10.1002/jmor.10382. ISSN 0362-2525. PMID 16217748. S2CID 15079072.
^ Lingham-Soliar, Theagarten; Feduccia, Alan; Wang, Xiaolin (2007). "Un nuevo espécimen chino indica que las 'protoplumas' del dinosaurio terópodo del Cretácico Inferior Sinosauropteryx son fibras de colágeno degradadas". Actas de la Royal Society B. Londres: Sociedad de la Realeza. 274 (1620): 1823–1829. doi :10.1098/rspb.2007.0352. ISSN 0962-8452. PMC 2270928 . PMID 17521978.
^ Prum, Richard O. (2003). "¿Son científicas las críticas actuales al origen terópodo de las aves? Refutación a Feduccia 2002". El alca . Washington, DC: Unión Estadounidense de Ornitólogos. 120 (2): 550–561. doi : 10.1642/0004-8038(2003)120[0550:ACCOTT]2.0.CO;2 . ISSN 0004-8038. JSTOR 4090212.
^ Wellnhofer, Peter (1988). "Un nuevo ejemplar de Archaeopteryx ". Ciencia . Washington, DC: Asociación Estadounidense para el Avance de la Ciencia. 240 (4860): 1790–1792. Código Bib : 1988 Ciencia... 240.1790W. doi : 10.1126/ciencia.240.4860.1790. ISSN 0036-8075. JSTOR 1701652. PMID 17842432. S2CID 32015255.
—— (1988). "Un nuevo ejemplar de Archaeopteryx ". Arqueoptérix . 6 : 1–30.
^ Schweitzer, María H.; Watt, JA; Avci, R.; et al. (1999). "Reactividad inmunológica específica de beta-queratina en estructuras parecidas a plumas del Cretácico Alvarezsáurido, Shuvuuia deserti ". Revista de Zoología Experimental Parte B. Hoboken, Nueva Jersey: Wiley-Blackwell. 285 (2): 146-157. doi :10.1002/(SICI)1097-010X(19990815)285:2<146::AID-JEZ7>3.0.CO;2-A. ISSN 1552-5007. PMID 10440726.
^ "Archaeopteryx: un madrugador". Berkeley: Museo de Paleontología de la Universidad de California . Consultado el 30 de octubre de 2019 .
^ O'Connor, Patrick M.; Claessens, León PAM (2005). "Diseño pulmonar aviar básico y ventilación de flujo continuo en dinosaurios terópodos no aviares". Naturaleza . Londres: Investigación de la naturaleza. 436 (7048): 253–256. Código Bib :2005Natur.436..253O. doi : 10.1038/naturaleza03716. ISSN 0028-0836. PMID 16015329. S2CID 4390587.
^ Gibson, Andrea (13 de julio de 2005). "Estudio: los dinosaurios depredadores tenían un sistema pulmonar similar al de un pájaro". Comunicaciones de investigación . Atenas, OH : Universidad de Ohio . Consultado el 18 de noviembre de 2019 .[ enlace muerto permanente ]
^ "El dinosaurio carnívoro de Argentina tenía un sistema respiratorio parecido al de un pájaro". Noticias de la Universidad de Michigan . Ann Arbor, MI : Oficina del Vicepresidente de Comunicaciones; Regentes de la Universidad de Michigan . 2 de octubre de 2008 . Consultado el 2 de noviembre de 2019 .
^ Xu, Xing; Norell, Mark A. (2004). "Un nuevo dinosaurio troodóntido de China con una postura para dormir similar a la de un ave" (PDF) . Naturaleza . Londres: Investigación de la naturaleza. 431 (7010): 838–841. Código Bib :2004Natur.431..838X. doi : 10.1038/naturaleza02898. ISSN 0028-0836. PMID 15483610. S2CID 4362745.
^ Norell, Mark A.; Clark, James M.; Chiappe, Luis M.; et al. (1995). "Un dinosaurio anidando". Naturaleza . Londres: Investigación de la naturaleza. 378 (6559): 774–776. Código Bib :1995Natur.378..774N. doi :10.1038/378774a0. ISSN 0028-0836. S2CID 4245228.
^ Varricchio, David J.; Moore, Jason R.; Erickson, Gregorio M .; et al. (2008). "El cuidado paterno de las aves tuvo origen en los dinosaurios". Ciencia . Washington, DC: Asociación Estadounidense para el Avance de la Ciencia. 322 (5909): 1826–1828. Código Bib : 2008 Ciencia... 322.1826V. doi : 10.1126/ciencia.1163245 . ISSN 0036-8075. PMID 19095938. S2CID 8718747.
^ Alas, Oliver (2007). "Una revisión de la función de los gastrolitos con implicaciones para los vertebrados fósiles y una clasificación revisada" (PDF) . Paleontológica Polonica . Varsovia: Instituto de Paleobiología, Academia Polaca de Ciencias . 52 (1): 1–16. ISSN 0567-7920. Archivado (PDF) desde el original el 17 de diciembre de 2008 . Consultado el 2 de noviembre de 2019 .
^ Longrich, NR; Tokaryk, T.; Campo, DJ (2011). "Extinción masiva de aves en el límite Cretácico-Paleógeno (K-Pg)". Procedimientos de la Academia Nacional de Ciencias . 108 (37): 15253–15257. Código bibliográfico : 2011PNAS..10815253L. doi : 10.1073/pnas.1110395108 . PMC 3174646 . PMID 21914849.
^ ab Renne, PR; Deino, AL; Hilgen, FJ; Kuiper, KF; Marcos, DF; Mitchell, WS; Morgan, LE; Mundil, R.; Smit, J. (2013). "Escalas de tiempo de eventos críticos alrededor del límite Cretácico-Paleógeno". Ciencia . 339 (6120): 684–687. Código Bib : 2013 Ciencia... 339..684R. doi : 10.1126/ciencia.1230492. PMID 23393261. S2CID 6112274.
^ abcde Brusatte, SL; Mayordomo, RJ; Barrett, PM; Carrano, MT; Evans, CC; Lloyd, GT; Mannion, PD; Norell, MA; Peppe, DJ; Upchurch, P.; Williamson, TE (2015). "La extinción de los dinosaurios". Reseñas biológicas . 90 (2): 628–642. doi : 10.1111/brv.12128 . hdl :20.500.11820/176e5907-26ec-4959-867f-0f2e52335f88. PMID 25065505. S2CID 115134484.
^ ab MacLeod, N.; Rawson, PF; Forey, PL; et al. (1997). "La transición biótica Cretácico-Terciario". Revista de la Sociedad Geológica . 154 (2): 265–292. Código Bib : 1997JGSoc.154..265M. doi :10.1144/gsjgs.154.2.0265. ISSN 0016-7649. S2CID 129654916.
^ ab Archibald, JD; Clemens, WA (1982). "Extinciones del Cretácico Tardío". Científico americano . 70 (4): 377–385. Código bibliográfico : 1982AmSci..70..377A. JSTOR 27851545.
^ Jablonski, D. (1991). "Extinciones: una perspectiva paleontológica". Ciencia . 253 (5021): 754–757. Código bibliográfico : 1991 Ciencia... 253..754J. doi : 10.1126/ciencia.253.5021.754. PMID 17835491.
^ Longrich, NR; Bhullar, B.-AS; Gauthier, JA (2012). "Extinción masiva de lagartos y serpientes en el límite Cretácico-Paleógeno". Procedimientos de la Academia Nacional de Ciencias . 109 (52): 21396–21401. Código bibliográfico : 2012PNAS..10921396L. doi : 10.1073/pnas.1211526110 . ISSN 0027-8424. PMC 3535637 . PMID 23236177.
^ Campo, DJ; Bercovici, A.; Berv, JS; Dunn, R.; Fastovsky, DE; Lyson, TR; Vajda, V.; Gauthier, JA (2018). "Evolución temprana de las aves modernas estructurada por el colapso global de los bosques en la extinción masiva del final del Cretácico". Biología actual . 28 (11): 1825–1831. doi : 10.1016/j.cub.2018.04.062 . PMID 29804807. S2CID 44075214.
^ ab Larson, DW; Marrón, CM; Evans, DC (2016). "Disparidad dental y estabilidad ecológica en dinosaurios parecidos a aves antes de la extinción masiva del final del Cretácico". Biología actual . 26 (10): 1325-1333. doi : 10.1016/j.cub.2016.03.039 . PMID 27112293. S2CID 3937001.
^ ab Le Loeuff, J. (2012). "Paleobiogeografía y biodiversidad de los dinosaurios del Maastrichtiano tardío: ¿cuántas especies de dinosaurios se extinguieron en el límite Cretácico-Terciario?". Boletín de la Société Géologique de Francia . 183 (6): 547–559. doi :10.2113/gssgfbull.183.6.547. ISSN 0037-9409.
^ Carpintero, K. (1983). "Evidencia que sugiere la extinción gradual de los dinosaurios del Cretácico más reciente". Naturwissenschaften . 70 (12): 611–612. Código bibliográfico : 1983NW.....70..611C. doi :10.1007/BF00377404. S2CID 20078285.
^ Russell, DA (1984). "El declive gradual de los dinosaurios: ¿realidad o falacia?". Naturaleza . 307 (5949): 360–361. Código Bib :1984Natur.307..360R. doi :10.1038/307360a0. S2CID 4269426.
^ Fastovsky, DE; Huang, Y.; Hsu, J.; Martín-McNaughton, J.; Sheehan, primer ministro; Weishampel, DB (2004). "Forma de la riqueza de dinosaurios mesozoicos" (PDF) . Geología . 32 (10): 877–880. Código Bib : 2004Geo....32..877F. doi :10.1130/G20695.1.
^ Sullivan, RM (2006). "La forma de la riqueza de dinosaurios mesozoicos: una reevaluación". En Lucas, SG; Sullivan, RM (eds.). Vertebrados del Cretácico tardío del interior occidental . Boletín del Museo de Historia Natural y Ciencias de Nuevo México. vol. 35. págs. 403–405.
^ Chiarenza, AA; Mannion, PD; Lunt, DJ; Farnsworth, A.; Jones, Luisiana; Kelland, SJ; Allison, Pensilvania (2019). "El modelado de nichos ecológicos no respalda la disminución de la diversidad de dinosaurios impulsada por el clima antes de la extinción masiva del Cretácico/Paleógeno". Comunicaciones de la naturaleza . 10 (1): 1–14. Código Bib : 2019NatCo..10.1091C. doi :10.1038/s41467-019-08997-2. PMC 6403247 . PMID 30842410.
^ Lloyd, GT (2012). "Un enfoque de modelado refinado para evaluar la influencia del muestreo en las curvas de paleobiodiversidad: nuevo apoyo para la disminución de la riqueza de dinosaurios del Cretácico". Cartas de biología . 8 (1): 123–126. doi :10.1098/rsbl.2011.0210. PMC 3259943 . PMID 21508029. S2CID 1376734.
^ Sakamoto, M.; Benton, MJ; Venditti, C. (2016). "Dinosaurios en declive decenas de millones de años antes de su extinción final". Procedimientos de la Academia Nacional de Ciencias . 113 (18): 5036–5040. Código Bib : 2016PNAS..113.5036S. doi : 10.1073/pnas.1521478113 . PMC 4983840 . PMID 27092007.
^ Barrett, primer ministro; McGowan, AJ; Página, V. (2009). "La diversidad de dinosaurios y el registro del rock". Actas de la Royal Society B: Ciencias Biológicas . 276 (1667): 2667–2674. doi :10.1098/rspb.2009.0352. PMC 2686664 . PMID 19403535.
^ Upchurch, P.; Mannion, PD; Benson, RB; Mayordomo, RJ; Carrano, MT (2011). "Controles geológicos y antropogénicos en el muestreo del registro fósil terrestre: un estudio de caso de Dinosauria". Sociedad Geológica, Londres, Publicaciones especiales . 358 (1): 209–240. Código Bib : 2011GSLSP.358..209U. doi :10.1144/SP358.14. S2CID 130777837.
^ Randal 2015
^ Álvarez, LW ; Álvarez, W .; Asaro, F .; Michel, HV (1980). "Causa extraterrestre de la extinción del Cretácico-Terciario" (PDF) . Ciencia . 208 (4448): 1095-1108. Código Bib : 1980 Ciencia... 208.1095A. CiteSeerX 10.1.1.126.8496 . doi : 10.1126/ciencia.208.4448.1095. ISSN 0036-8075. PMID 17783054. S2CID 16017767. Archivado desde el original (PDF) el 8 de julio de 2010 . Consultado el 30 de octubre de 2019 .
^ Bohor, BF; Modreski, PJ; Foord, EE (1987). "Cuarzo impactado en las arcillas límite Cretácico-Terciario: evidencia de una distribución global". Ciencia . 236 (4802): 705–709. Código bibliográfico : 1987 Ciencia... 236..705B. doi : 10.1126/ciencia.236.4802.705. PMID 17748309. S2CID 31383614.
^ Hildebrand, AR; Penfield, GT; Kring, DA; Pilkington, M.; Camargo, ZA; Jacobsen, SB; Boynton, WV (1991). "Cráter Chicxulub: un posible cráter de impacto en el límite Cretácico / Terciario en la Península de Yucatán, México". Geología . 19 (9): 867–871. Código Bib :1991Geo....19..867H. doi :10.1130/0091-7613(1991)019<0867:CCAPCT>2.3.CO;2.
^ Papa, KO ; Ocampo, A.C .; Kinsland, GL; et al. (1996). "Expresión superficial del cráter Chicxulub". Geología . Boulder, CO: Sociedad Geológica de América. 24 (6): 527–530. Código Bib : 1996Geo....24..527P. doi :10.1130/0091-7613(1996)024<0527:SEOTCC>2.3.CO;2. ISSN 0091-7613. PMID 11539331.
^ Schulte, P.; Alegret, L.; Arenillas, I.; Arz, JA; Barton, PJ; Bown, PR; Bralower, TJ; Christeson, GL; Claeys, P.; Cockell, CS; Collins, GS; Deutsch, A.; Goldin, TJ; Ir a, K.; Grajales-Nishimura, JM; Afligirse, RAF; Gulick, SPS; Johnson, KR; Kiessling, W.; Koeberl, C.; Kring, DA; MacLeod, KG; Matsui, T.; Melosh, J.; Montanari, A.; Morgan, JV; Neal, CR; Nicolas, DJ; Norris, RD; Pierazzo, E.; Ravizza, G.; Rebolledo-Vieyra, M.; Uwe Reimold, W.; Robin, E.; Salge, T.; Speijer, RP; Dulce, AR; Urrutia-Fucugauchi, J.; Vajda, V.; Whalen, MT; Willumsen, PS (2010). "El impacto del asteroide Chicxulub y la extinción masiva en el límite Cretácico-Paleógeno". Ciencia . 327 (5970): 1214–1218. Código Bib : 2010 Ciencia... 327.1214S. doi : 10.1126/ciencia.1177265. PMID 20203042. S2CID 2659741.
^ Kring, DA (2007). "El evento de impacto de Chicxulub y sus consecuencias ambientales en el límite Cretácico-Terciario". Paleogeografía, Paleoclimatología, Paleoecología . 255 (1–2): 4–21. Código Bib : 2007PPP...255....4K. doi :10.1016/j.palaeo.2007.02.037.
^ ab Chiarenza, AA; Farnsworth, A.; Mannion, PD; Lunt, DJ; Valdés, PJ; Morgan, JV ; Allison, Pensilvania (2020). "El impacto de un asteroide, no el vulcanismo, causó la extinción de los dinosaurios del final del Cretácico". Procedimientos de la Academia Nacional de Ciencias . 117 (29): 17084–17093. Código Bib : 2020PNAS..11717084C. doi : 10.1073/pnas.2006087117 . PMC 7382232 . PMID 32601204.
^ Ivanov, Licenciado en Letras (2005). "Modelado numérico de los cráteres de meteoritos terrestres más grandes". Investigación del Sistema Solar . 39 (5): 381–409. Código Bib : 2005SoSyR..39..381I. doi :10.1007/s11208-005-0051-0. S2CID 120305483.
^ Matsui, T.; Imamura, F.; Tajika, E.; Nakano, Y.; Fujisawa, Y. (2002). "Generación y propagación de un tsunami a partir del evento de impacto Cretácico-Terciario". Documentos especiales de la Sociedad Geológica de América . 356 : 69–78. doi :10.1130/0-8137-2356-6.69. ISBN9780813723563.
^ Robertson, DS; McKenna, MC ; Dibujos animados, OB ; et al. (2004). «Supervivencia en las primeras horas del Cenozoico» (PDF) . Boletín de la Sociedad Geológica de América . 116 (5–6): 760–768. Código Bib : 2004GSAB..116..760R. doi :10.1130/B25402.1. ISSN 0016-7606. Archivado desde el original (PDF) el 18 de septiembre de 2012 . Consultado el 15 de junio de 2011 .
^ Robertson, DS; Lewis, WM; Sheehan, primer ministro; Dibujos animados, OB (2013). "Extinción K-Pg: reevaluación de la hipótesis del calor-fuego". Revista de Investigación Geofísica: Biogeociencias . 118 (1): 329–336. Código Bib : 2013JGRG..118..329R. doi : 10.1002/jgrg.20018 . S2CID 17015462.
^ Papa, KO; Baines, KH; Ocampo, A.C.; Ivanov, Licenciado en Letras (1997). "Energía, producción volátil y efectos climáticos del impacto Cretácico / Terciario de Chicxulub". Revista de investigación geofísica: planetas . 102 (E9): 21645–21664. Código bibliográfico : 1997JGR...10221645P. doi : 10.1029/97JE01743 . PMID 11541145. S2CID 8447773.
^ ab Ohno, S.; Kadono, T.; Kurosawa, K.; Hamura, T.; Sakaiya, T.; Shigemori, K.; Hironaka, Y.; Sanó, T.; Watari, T.; Otani, K.; Matsui, T.; Sugita, S. (2014). "Producción de vapor rico en sulfatos durante el impacto de Chicxulub e implicaciones para la acidificación de los océanos". Geociencia de la naturaleza . 7 (4): 279–282. Código Bib : 2014NatGe...7..279O. doi : 10.1038/ngeo2095.
^ Kaiho, K.; Oshima, N.; Adachi, K.; Adachi, Y.; Mizukami, T.; Fujibayashi, M.; Saito, R. (2016). "El cambio climático global impulsado por el hollín en el límite K-Pg como causa de la extinción masiva". Informes científicos . 6 (1): 1–13. Código Bib : 2016NatSR...628427K. doi : 10.1038/srep28427. PMC 4944614 . PMID 27414998.
^ Lyon, SL; Karp, AT; Bralower, TJ; Grice, K.; Schaefer, B.; Gulick, SP; Morgan, JV ; Freeman, KH (2020). "La materia orgánica del cráter Chicxulub exacerbó el invierno del impacto K-Pg". Procedimientos de la Academia Nacional de Ciencias . 117 (41): 25327–25334. Código Bib : 2020PNAS..11725327L. doi : 10.1073/pnas.2004596117 . PMC 7568312 . PMID 32989138.
^ Chenet, Alabama; Courtillot, V.; Fluteau, F.; Gerard, M.; Quidelleur, X.; Khadri, SFR; Subbarao, KV; Thordarson, T. (2009). "Determinación de erupciones rápidas del Deccan a lo largo del límite Cretácico-Terciario utilizando variación secular paleomagnética: 2. Restricciones del análisis de ocho nuevas secciones y síntesis para una sección compuesta de 3500 m de espesor" (PDF) . Revista de investigación geofísica: Tierra sólida . 114 (B6): B06103. Código Bib : 2009JGRB..114.6103C. doi :10.1029/2008JB005644. S2CID 140541003.
^ Schoene, B.; Eddy, diputado; Samperton, KM; Keller, CB; Keller, G.; Adatte, T.; Khadri, SF (2019). "Restricciones de U-Pb en la erupción pulsada de las trampas del Deccan durante la extinción masiva del final del Cretácico". Ciencia . 363 (6429): 862–866. Código Bib : 2019 Ciencia... 363..862S. doi : 10.1126/ciencia.aau2422 . OSTI 1497969. PMID 30792300. S2CID 67876950.
^ ab McLean, DM (1985). "Desgasificación del manto de Deccan Traps en las extinciones marinas terminales del Cretácico". Investigación del Cretácico . 6 (3): 235–259. Código Bib : 1985CrRes...6..235M. doi :10.1016/0195-6671(85)90048-5.
^ Yo, S.; Widowson, M.; Thordarson, T.; Jay, AE (2006). "Flujos volátiles durante las erupciones de inundación de basalto y efectos potenciales sobre el medio ambiente global: una perspectiva del Deccan". Cartas sobre ciencias planetarias y de la Tierra . 248 (1–2): 518–532. Código Bib : 2006E y PSL.248..518S. doi :10.1016/j.epsl.2006.05.041.
^ Tobin, TS; Bitz, CM; Arquero, D. (2017). "Modelado de los efectos climáticos de las emisiones de dióxido de carbono de las erupciones volcánicas de las trampas del Deccan alrededor del límite Cretácico-Paleógeno". Paleogeografía, Paleoclimatología, Paleoecología . 478 : 139-148. Código Bib : 2017PPP...478..139T. doi :10.1016/j.palaeo.2016.05.028.
^ Schmidt, A.; Skeffington, RA; Thordarson, T.; Yo, S.; Forster, PM; Rap, A.; Ridgwell, A.; Fowler, D.; Wilson, M.; Mann, GW; Wignall, PB; Carslaw, KS (2016). "Estrés ambiental selectivo por el azufre emitido por las erupciones de basalto de inundaciones continentales" (PDF) . Geociencia de la naturaleza . 9 (1): 77–82. Código Bib : 2016NatGe...9...77S. doi : 10.1038/ngeo2588. S2CID 59518452. Archivado (PDF) desde el original el 22 de septiembre de 2017.
^ Hofman, C.; Féraud, G.; Courtillot, V. (2000). " Datación 40 Ar / 39 Ar de separaciones minerales y rocas enteras de la pila de lava de los Ghats occidentales: mayores limitaciones en la duración y edad de las trampas del Deccan". Cartas sobre ciencias planetarias y de la Tierra . 180 (1–2): 13–27. Código Bib : 2000E y PSL.180...13H. doi :10.1016/S0012-821X(00)00159-X. ISSN 0012-821X.
^ Sahni, A. (1988). "Eventos en la frontera Cretácico-Terciario: extinciones masivas, enriquecimiento de iridio y vulcanismo del Deccan". Ciencia actual . 57 (10): 513–519. JSTOR 24090754.
^ Glasby, médico de cabecera; Kunzendorf, H. (1996). "Múltiples factores en el origen del límite Cretácico / Terciario: el papel del estrés ambiental y el vulcanismo de la Trampa del Deccan". Geologische Rundschau . 85 (2): 191–210. Código Bib : 1996IJEaS..85..191G. doi :10.1007/BF02422228. PMID 11543126. S2CID 19155384.
^ Álvarez, LW (1987). Extinciones masivas causadas por grandes impactos de bólidos (Informe). Laboratorio Lawrence Berkeley . pag. 39. LBL-22786 . Consultado el 27 de enero de 2021 .
^ Álvarez 1997, págs. 130-146, cap. 7: “El mundo después de Chicxulub”.
^ Renne, relaciones públicas; Esguince, CJ; Richards, MA; Yo, S.; Vanderkluysen, L.; Pandé, K. (2015). "Cambio de estado en el vulcanismo del Deccan en el límite Cretácico-Paleógeno, posiblemente inducido por el impacto". Ciencia . 350 (6256): 76–78. Código Bib : 2015 Ciencia... 350... 76R. doi : 10.1126/ciencia.aac7549 . PMID 26430116. S2CID 30612906.
^ Richards, MA; Álvarez, W.; Yo, S.; Karlstrom, L.; Renne, PR; Manga, M.; Esguince, CJ; Smith, J.; Vanderkluysen, L.; Gibson, SA (2015). "Desencadenamiento de las mayores erupciones del Deccan por el impacto de Chicxulub". Boletín de la Sociedad Geológica de América . 127 (11-12): 1507-1520. Código Bib : 2015GSAB..127.1507R. doi :10.1130/B31167.1. S2CID 3463018.
^ Khazins, V.; Shuvalov, V. (2019). "Impacto de Chicxulub como desencadenante de una de las fases del vulcanismo del Deccan: umbral de densidad de energía sísmica". En Kocharyan, G.; Lyakhov, A. (eds.). Efectos desencadenantes en geosistemas . Procedimientos Springer en Ciencias de la Tierra y el Medio Ambiente. Cham: Springer. págs. 523–530. doi :10.1007/978-3-030-31970-0_55. ISBN978-3-030-31969-4. S2CID 210277965.
^ Archibald, JD; Clemens, Washington; Padián, K.; Rowe, T.; Macleod, N.; Barrett, PM; Gale, A.; Holroyd, P.; Sues, H.-D.; Arens, Carolina del Norte; Horner, JR; Wilson, médico de cabecera; Goodwin, MB; Brochu, California; Lofgren, DL; Hurlbert, SH; Hartman, JH; Eberth, DA; Wignall, PB; Currie, PJ; Weil, A.; Prasad, GVR; Dingus, L.; Courtillot, V.; Milner, A.; Milner, A.; Bajpai, S.; Sala, DJ; Sahni, A. (2010). "Extinciones del Cretácico: múltiples causas". Ciencia . 328 (5981): 973, respuesta del autor 975–6. doi :10.1126/ciencia.328.5981.973-a. PMID 20489004.
^ Courtillot, V.; Fluteau, F. (2010). "Extinciones del Cretácico: la hipótesis volcánica". Ciencia . 328 (5981): 973–974. doi :10.1126/ciencia.328.5981.973-b. PMID 20489003.
^ Keller, G. (2014). "El vulcanismo del Deccan, el impacto de Chicxulub y la extinción masiva del final del Cretácico: ¿coincidencia? Causa y efecto". Documentos especiales de la Sociedad Geológica de América . 505 : 57–89. doi :10.1130/2014.2505(03). ISBN9780813725055.
^ Fassett, JE; Heaman, LM; Simonetti, A. (2011). "Datación directa U-Pb de huesos de dinosaurios del Cretácico y Paleoceno, Cuenca de San Juan, Nuevo México". Geología . 39 (2): 159-162. Código Bib : 2011Geo....39..159F. doi :10.1130/G31466.1. ISSN 0091-7613.
^ Fassett, JE; Heaman, LM; Simonetti, A. (2009). "Nueva evidencia geocronológica y estratigráfica confirma la edad del Paleoceno de la formación Animas y arenisca de Ojo Alamo, portadora de dinosaurios, en la cuenca de San Juan, Nuevo México y Colorado". Paleontología Electrónica . 12 (1): 3A.
^ Sloan, RE; Rigby, JK Jr.; Van Valen, LM ; et al. (1986). "Extinción gradual de dinosaurios y radiación ungulada simultánea en la formación Hell Creek". Ciencia . 232 (4750): 629–633. Código Bib : 1986 Ciencia... 232..629S. doi : 10.1126/ciencia.232.4750.629. ISSN 0036-8075. PMID 17781415. S2CID 31638639.
^ Lucas, SG; Sullivan, RM; Cather, SM; Jasinski, SE; Fowler, DW; Heckert, AB; Spielmann, JA; Cazar, AP (2009). "No hay evidencia definitiva de dinosaurios del Paleoceno en la Cuenca de San Juan". Paleontología Electrónica . 12 (2): 8A.
^ Renne, relaciones públicas; Goodwin, MB (2012). "Datación directa U-Pb de huesos de dinosaurios del Cretácico y Paleoceno, Cuenca de San Juan, Nuevo México: COMENTARIO". Geología . 40 (4): e259. Código Bib : 2012Geo....40E.259R. doi : 10.1130/G32521C.1 .
^ Lofgren, DL; Hotton, CL; Runkel, AC (1990). "Reelaboración de dinosaurios del Cretácico en canal del Paleoceno, depósitos, formación Hell Creek superior, Montana". Geología . 18 (9): 874–877. Código bibliográfico : 1990Geo....18..874L. doi :10.1130/0091-7613(1990)018<0874:ROCDIP>2.3.CO;2.
^ Koenig, AE; Lucas, SG; Neymark, Los Ángeles; Heckert, AB; Sullivan, RM; Jasinski, SE; Fowler, DW (2012). "Datación directa U-Pb de huesos de dinosaurios del Cretácico y Paleoceno, Cuenca de San Juan, Nuevo México: COMENTARIO". Geología . 40 (4): e262. Código Bib : 2012Geo....40E.262K. doi : 10.1130/G32154C.1 .
^ Sarjeant 1995, págs. 255–284, cap. 15: "Los dinosaurios y la dinomanía durante 150 años" de Hugh S. Torrens .
^ Currie y Padian 1997, págs. 347–350, "Historia de los descubrimientos de dinosaurios: primer período dorado" por Brent H. Breithaupt.
^ Dickens 1853, pag. 1, cap. I: " Londres. El trimestre de San Miguel acaba de terminar y el Lord Canciller está sentado en el Lincoln's Inn Hall. Un clima de noviembre implacable. Tanto barro en las calles, como si las aguas acabaran de retirarse de la faz de la tierra y no "Será maravilloso encontrar un Megalosaurus, de unos doce metros de largo, caminando como un lagarto elefante por Holborn Hill. "
^ Farlow y Brett-Surman 1997, págs. 675–697, cap. 43: "Los dinosaurios y los medios" de Donald F. Glut y MK Brett-Surman.
^ Lee, Newton; Madej, Krystina (2012). "Animación temprana: chistes y situaciones". Historias de Disney . págs. 17-24. doi :10.1007/978-1-4614-2101-6_3. ISBN978-1-4614-2100-9. S2CID 192335675.






_(19798844494).jpg/1280px-Bulletin_(1969)_(19798844494).jpg)









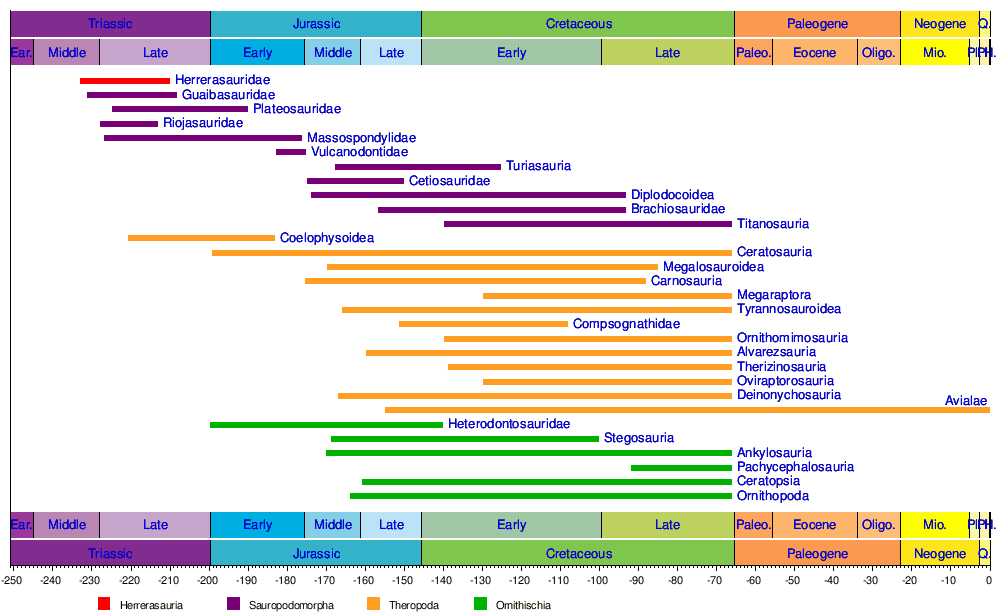




_adult_male_in_flight-cropped.jpg/1280px-Bee_hummingbird_(Mellisuga_helenae)_adult_male_in_flight-cropped.jpg)












{kind=link}