Un mamífero (del latín mamma 'pecho') [1] es un animal vertebrado de la clase Mammalia ( / m ə ˈ m eɪ l i . ə / ). Los mamíferos se caracterizan por la presencia de glándulas mamarias productoras de leche para alimentar a sus crías, una amplia región neocortical del cerebro, pelaje o pelo y tres huesos del oído medio . Estas características los distinguen de los reptiles y las aves , de los que sus ancestros divergieron en el Periodo Carbonífero hace más de 300 millones de años. Se han descrito alrededor de 6.400 especies existentes de mamíferos y se han dividido en 29 órdenes .
Los órdenes más grandes de mamíferos, por número de especies , son los roedores , los murciélagos y los eulipotiflos (que incluyen erizos , topos y musarañas ). Los tres siguientes son los primates (que incluyen humanos , monos y lémures ), los ungulados de dedos pares (que incluyen cerdos , camellos y ballenas ) y los carnívoros (que incluyen gatos , perros y focas ).
Los mamíferos son los únicos miembros vivos de Synapsida ; este clado , junto con Sauropsida (reptiles y aves), constituye el clado más grande Amniota . Los primeros sinápsidos se conocen como " pelicosaurios ". Los terápsidos más avanzados se volvieron dominantes durante el Guadalupiano . Los mamíferos se originaron a partir de los cinodontos , un grupo avanzado de terápsidos, durante el Triásico Tardío al Jurásico Temprano . Los mamíferos alcanzaron su diversidad moderna en los períodos Paleógeno y Neógeno de la era Cenozoica , después de la extinción de los dinosaurios no aviares , y han sido el grupo animal terrestre dominante desde hace 66 millones de años hasta el presente.
El tipo básico de cuerpo de los mamíferos es cuadrúpedo , y la mayoría de ellos utilizan cuatro extremidades para la locomoción terrestre ; pero en algunos, las extremidades están adaptadas para la vida en el mar , en el aire , en los árboles o bajo tierra . Los bípedos se han adaptado para moverse utilizando solo las dos extremidades inferiores, mientras que las extremidades traseras de los cetáceos y las vacas marinas son meros vestigios internos . Los mamíferos varían en tamaño desde el murciélago abejorro de 30 a 40 milímetros (1,2 a 1,6 pulgadas) hasta la ballena azul de 30 metros (98 pies), posiblemente el animal más grande que haya vivido jamás. La esperanza de vida máxima varía de dos años para la musaraña a 211 años para la ballena de Groenlandia . Todos los mamíferos modernos dan a luz crías vivas, excepto las cinco especies de monotremas , que ponen huevos. El grupo más rico en especies es el de los mamíferos placentarios vivíparos , llamados así por el órgano temporal ( placenta ) que utilizan las crías para obtener nutrición de la madre durante la gestación .
La mayoría de los mamíferos son inteligentes , algunos poseen cerebros grandes, conciencia de sí mismos y uso de herramientas . Los mamíferos pueden comunicarse y vocalizar de varias maneras, incluida la producción de ultrasonidos , marcado de olores , señales de alarma , canto , ecolocalización y, en el caso de los humanos, lenguaje complejo . Los mamíferos pueden organizarse en sociedades de fisión-fusión , harenes y jerarquías , pero también pueden ser solitarios y territoriales . La mayoría de los mamíferos son polígamos , pero algunos pueden ser monógamos o poliándricos .
La domesticación de muchos tipos de mamíferos por parte de los humanos jugó un papel importante en la Revolución Neolítica y dio como resultado que la agricultura reemplazara a la caza y la recolección como la principal fuente de alimento para los humanos. Esto condujo a una importante reestructuración de las sociedades humanas de nómadas a sedentarias, con una mayor cooperación entre grupos cada vez más grandes y, en última instancia, al desarrollo de las primeras civilizaciones . Los mamíferos domesticados proporcionaron, y continúan proporcionando, energía para el transporte y la agricultura, así como alimentos ( carne y productos lácteos ), pieles y cuero . Los mamíferos también son cazados y sometidos a carreras por deporte, se mantienen como mascotas y animales de trabajo de varios tipos, y se utilizan como organismos modelo en la ciencia. Los mamíferos han sido representados en el arte desde el Paleolítico y aparecen en la literatura, el cine, la mitología y la religión. La disminución en las cantidades y la extinción de muchos mamíferos se debe principalmente a la caza furtiva humana y la destrucción del hábitat , principalmente la deforestación .
Más del 70% de las especies de mamíferos pertenecen a los órdenes Rodentia , Chiroptera y Eulipotyphla .
La clasificación de los mamíferos ha pasado por varias revisiones desde que Carl Linnaeus definió inicialmente la clase, y en la actualidad [ ¿cuándo? ] , ningún sistema de clasificación es universalmente aceptado. McKenna y Bell (1997) y Wilson y Reeder (2005) proporcionan compendios recientes útiles. [2] Simpson (1945) [3] proporciona sistemáticas de los orígenes y relaciones de los mamíferos que se habían enseñado universalmente hasta finales del siglo XX. Sin embargo, desde 1945, se ha encontrado gradualmente una gran cantidad de información nueva y más detallada: el registro paleontológico ha sido recalibrado, y los años intermedios han visto mucho debate y progreso en relación con los fundamentos teóricos de la sistematización en sí, en parte a través del nuevo concepto de cladística . Aunque el trabajo de campo y de laboratorio desactualizó progresivamente la clasificación de Simpson, sigue siendo lo más cercano a una clasificación oficial de los mamíferos, a pesar de sus problemas conocidos. [4]
La mayoría de los mamíferos, incluidos los seis órdenes más ricos en especies , pertenecen al grupo placentario. Los tres órdenes más grandes en número de especies son Rodentia : ratones , ratas , puercoespines , castores , capibaras y otros mamíferos roedores; Chiroptera : murciélagos; y Soricomorpha : musarañas , topos y solenodontes . Los siguientes tres órdenes más grandes, dependiendo del esquema de clasificación biológica utilizado, son los primates : simios , monos y lémures ; los Cetartiodactyla : ballenas y ungulados de dedos pares ; y los Carnivora que incluyen gatos , perros , comadrejas , osos , focas y aliados. [5] Según Mammal Species of the World , en 2006 se identificaron 5416 especies. Estas se agruparon en 1229 géneros , 153 familias y 29 órdenes. [5] En 2008, la Unión Internacional para la Conservación de la Naturaleza (UICN) completó una Evaluación Global de Mamíferos de cinco años para su Lista Roja de la UICN , que contabilizó 5.488 especies. [6] Según una investigación publicada en el Journal of Mammalogy en 2018, el número de especies de mamíferos reconocidas es de 6.495, incluidas 96 recientemente extintas. [7]
La palabra "mamífero" es moderna, del nombre científico Mammalia acuñado por Carl Linnaeus en 1758, derivado del latín mamma ("teta, pap"). En un influyente artículo de 1988, Timothy Rowe definió a Mammalia filogenéticamente como el grupo corona de los mamíferos, el clado que consiste en el ancestro común más reciente de los monotremas vivos ( equidnas y ornitorrincos ) y mamíferos terios ( marsupiales y placentarios ) y todos los descendientes de ese ancestro. [8] Dado que este ancestro vivió en el período Jurásico , la definición de Rowe excluye a todos los animales del Triásico anterior , a pesar del hecho de que los fósiles Triásicos en Haramiyida han sido referidos a Mammalia desde mediados del siglo XIX. [9] Si Mammalia se considera como el grupo corona, su origen puede fecharse aproximadamente como la primera aparición conocida de animales más estrechamente relacionados con algunos mamíferos existentes que con otros. Ambondro está más estrechamente relacionado con los monotremas que con los mamíferos terianos, mientras que Amphilestes y Amphitherium están más estrechamente relacionados con los terianos; como los fósiles de los tres géneros datan de hace unos 167 millones de años en el Jurásico Medio , esta es una estimación razonable para la aparición del grupo corona. [10]
TS Kemp ha proporcionado una definición más tradicional: " Sinápsidos que poseen una articulación mandibular dentario - escamosa y oclusión entre molares superiores e inferiores con un componente transversal en el movimiento" o, equivalentemente en la visión de Kemp, el clado que se originó con el último ancestro común de Sinoconodon y los mamíferos actuales. [11] El sinápsido más antiguo conocido que satisface las definiciones de Kemp es Tikitherium , datado hace 225 Ma , por lo que la aparición de mamíferos en este sentido más amplio puede recibir esta fecha del Triásico Tardío . [12] [13] Sin embargo, este animal puede haber evolucionado en realidad durante el Neógeno. [14]

A principios del siglo XXI, los estudios moleculares basados en el análisis de ADN han sugerido nuevas relaciones entre las familias de mamíferos. La mayoría de estos hallazgos han sido validados de forma independiente por datos de presencia/ausencia de retrotransposones . [16] Los sistemas de clasificación basados en estudios moleculares revelan tres grupos o linajes principales de mamíferos placentarios: Afrotheria , Xenarthra y Boreoeutheria , que divergieron en el Cretácico . Las relaciones entre estos tres linajes son polémicas, y se han propuesto las tres hipótesis posibles con respecto a qué grupo es basal . Estas hipótesis son Atlantogenata (Boreoeutheria basal), Epitheria (Xenarthra basal) y Exafroplacentalia (Afrotheria basal). [17] Boreoeutheria a su vez contiene dos linajes principales: Euarchontoglires y Laurasiatheria .
Las estimaciones de los tiempos de divergencia entre estos tres grupos placentarios varían entre 105 y 120 millones de años atrás, dependiendo del tipo de ADN utilizado (como el nuclear o el mitocondrial ) [18] y de las diferentes interpretaciones de los datos paleogeográficos . [17]
Synapsida , un clado que contiene mamíferos y sus parientes extintos, se originó durante el subperíodo Pensilvánico (hace unos 323 millones a unos 300 millones de años), cuando se separaron del linaje de los reptiles. Los mamíferos del grupo corona evolucionaron a partir de las formas anteriores de los mamíferos durante el Jurásico Temprano . El cladograma considera a Mammalia como el grupo corona. [22]

Los primeros vertebrados completamente terrestres fueron los amniotas . Al igual que sus predecesores anfibios tetrápodos , tenían pulmones y extremidades. Sin embargo, los huevos amnióticos tienen membranas internas que permiten que el embrión en desarrollo respire, pero mantienen el agua dentro. Por lo tanto, los amniotas pueden poner huevos en tierra firme, mientras que los anfibios generalmente necesitan poner sus huevos en el agua.
Los primeros amniotas aparentemente surgieron en el subperíodo Pensilvánico del Carbonífero . Descendían de tetrápodos anfibios reptiliomorfos anteriores , [23] que vivían en tierra que ya estaba habitada por insectos y otros invertebrados, así como helechos , musgos y otras plantas. En unos pocos millones de años, dos linajes importantes de amniotas se volvieron distintos: los sinápsidos , que más tarde incluirían al ancestro común de los mamíferos; y los saurópsidos , que ahora incluyen tortugas , lagartos , serpientes , cocodrilos y dinosaurios (incluidas las aves ). [24] Los sinápsidos tienen un solo orificio ( fenestra temporal ) bajo a cada lado del cráneo. Los sinápsidos primitivos incluían los animales más grandes y feroces del Pérmico temprano, como Dimetrodon . [25] Los sinápsidos no mamíferos fueron llamados tradicionalmente (e incorrectamente) "reptiles parecidos a mamíferos" o pelicosaurios ; ahora sabemos que no eran reptiles ni parte del linaje de los reptiles. [26] [27]
Los terápsidos , un grupo de sinápsidos, evolucionaron en el Pérmico medio , hace unos 265 millones de años, y se convirtieron en los vertebrados terrestres dominantes. [26] Se diferencian de los eupelicosaurios basales en varias características del cráneo y las mandíbulas, incluyendo: cráneos más grandes e incisivos que son iguales en tamaño en los terápsidos, pero no en los eupelicosaurios. [26] El linaje de los terápsidos que conduce a los mamíferos pasó por una serie de etapas, comenzando con animales que eran muy similares a sus primeros ancestros sinápsidos y terminando con los cinodontos probainognatios , algunos de los cuales podrían confundirse fácilmente con mamíferos. Esas etapas se caracterizaron por: [28]
La extinción masiva del Pérmico-Triásico , que tuvo lugar hace unos 252 millones de años y fue prolongada debido a la acumulación de varios pulsos de extinción, puso fin al predominio de los terápsidos carnívoros. [30] A principios del Triásico, la mayoría de los nichos de carnívoros terrestres medianos y grandes fueron ocupados por arcosaurios [31] que, durante un período prolongado (35 millones de años), llegaron a incluir a los crocodilomorfos , [32] los pterosaurios y los dinosaurios; [33] sin embargo, los grandes cinodontes como el Trucidocynodon y los traversodóntidos todavía ocupaban nichos de carnívoros y herbívoros de gran tamaño, respectivamente. En el Jurásico, los dinosaurios habían llegado a dominar también los grandes nichos de herbívoros terrestres. [34]
Los primeros mamíferos (en el sentido de Kemp) aparecieron en la época del Triásico Tardío (hace unos 225 millones de años), 40 millones de años después de los primeros terápsidos. Se expandieron fuera de su nicho de insectívoros nocturnos desde mediados del Jurásico en adelante; [35] el Castorocauda jurásico , por ejemplo, era un pariente cercano de los mamíferos verdaderos que tenían adaptaciones para nadar, cavar y atrapar peces. [36] Se cree que la mayoría, si no todos, siguieron siendo nocturnos (el cuello de botella nocturno ), lo que explica gran parte de los rasgos típicos de los mamíferos. [37] La mayoría de las especies de mamíferos que existieron en la Era Mesozoica eran multituberculados, eutriconodontos y espalacotheriids . [38] El metaterio conocido más antiguo es Sinodelphys , encontrado en esquisto del Cretácico Inferior de 125 millones de años en la provincia nororiental china de Liaoning . El fósil está casi completo e incluye mechones de pelo y huellas de tejidos blandos. [39]

El fósil más antiguo conocido entre los Eutheria ("bestias verdaderas") es la pequeña Juramaia sinensis , o "madre jurásica de China", con forma de musaraña, que data de hace 160 millones de años en el Jurásico tardío. [40] Un pariente euterio posterior, Eomaia , que data de hace 125 millones de años en el Cretácico temprano, poseía algunas características en común con los marsupiales pero no con los placentarios, evidencia de que estas características estaban presentes en el último ancestro común de los dos grupos pero luego se perdieron en el linaje placentario. [41] En particular, los huesos epipúbicos se extienden hacia adelante desde la pelvis. Estos no se encuentran en ningún placentario moderno, pero sí en marsupiales, monotremas, otros mamíferos no teriosos y Ukhaatherium , un animal del Cretácico temprano en el orden euterio Asioryctitheria . Esto también se aplica a los multituberculados. [42] Aparentemente son una característica ancestral, que posteriormente desapareció en el linaje placentario. Estos huesos epipúbicos parecen funcionar endureciendo los músculos durante la locomoción, reduciendo la cantidad de espacio que se presenta, que los placentarios requieren para contener a su feto durante los períodos de gestación. Una salida pélvica estrecha indica que las crías eran muy pequeñas al nacer y, por lo tanto, el embarazo fue corto, como en los marsupiales modernos. Esto sugiere que la placenta fue un desarrollo posterior. [43]
Uno de los primeros monotremas conocidos fue Teinolophos , que vivió hace unos 120 millones de años en Australia. [44] Los monotremas tienen algunas características que pueden heredarse de los amniotas originales, como el mismo orificio para orinar, defecar y reproducirse ( cloaca ) —como también lo hacen los lagartos y las aves— [45] y ponen huevos que son correosos y no calcificados. [46]
Hadrocodium , cuyos fósiles datan de hace aproximadamente 195 millones de años, en el Jurásico temprano , proporciona la primera evidencia clara de una articulación de la mandíbula formada únicamente por los huesos escamoso y dentario; no hay espacio en la mandíbula para el articular, un hueso involucrado en las mandíbulas de todos los sinápsidos tempranos. [47]

La evidencia clara más temprana de pelo o pelaje se encuentra en fósiles de Castorocauda y Megaconus , de hace 164 millones de años a mediados del Jurásico. En la década de 1950, se sugirió que los agujeros (pasajes) en los maxilares y premaxilares (huesos en la parte delantera de la mandíbula superior) de los cinodontos eran canales que suministraban vasos sanguíneos y nervios a las vibrisas ( bigotes ) y, por lo tanto, eran evidencia de pelo o pelaje; [48] [49] Sin embargo, pronto se señaló que los agujeros no muestran necesariamente que un animal tenía vibrisas, ya que el lagarto moderno Tupinambis tiene agujeros que son casi idénticos a los encontrados en el cinodonte no mamífero Thrinaxodon . [27] [50] Sin embargo, las fuentes populares continúan atribuyendo bigotes a Thrinaxodon . [51] Los estudios sobre coprolitos del Pérmico sugieren que los sinápsidos no mamíferos de la época ya tenían pelo, lo que posiblemente remonta la evolución del pelo a los dicinodontes . [52]
No se sabe con certeza cuándo apareció por primera vez la endotermia en la evolución de los mamíferos, aunque en general se acepta que evolucionó por primera vez en terápsidos no mamíferos . [52] [53] Los monotremas modernos tienen temperaturas corporales más bajas y tasas metabólicas más variables que los marsupiales y los placentarios, [54] pero hay evidencia de que algunos de sus ancestros, quizás incluidos los ancestros de los terios, pueden haber tenido temperaturas corporales como las de los terios modernos. [55] Del mismo modo, algunos terios modernos como los afroterios y los xenartros han desarrollado secundariamente temperaturas corporales más bajas. [56]
La evolución de las extremidades erectas en los mamíferos es incompleta: los monotremas vivos y fósiles tienen extremidades extendidas. La postura parasagital (no extendida) de las extremidades apareció en algún momento a finales del Jurásico o principios del Cretácico; se encuentra en el euterio Eomaia y el metaterio Sinodelphys , ambos datados hace 125 millones de años. [57] Los huesos epipúbicos , una característica que influyó fuertemente en la reproducción de la mayoría de los clados de mamíferos, se encuentran por primera vez en Tritylodontidae , lo que sugiere que es una sinapomorfía entre ellos y Mammaliaformes . Son omnipresentes en Mammaliaformes no placentarios, aunque Megazostrodon y Erythrotherium parecen haber carecido de ellos. [58]
Se ha sugerido que la función original de la lactancia ( producción de leche ) era mantener húmedos los huevos. Gran parte de este argumento se basa en los monotremas, los mamíferos que ponen huevos. [59] [60] En las hembras humanas, las glándulas mamarias se desarrollan completamente durante la pubertad, independientemente del embarazo. [61]

Los mamíferos terios se apoderaron de los nichos ecológicos de tamaño mediano a grande en el Cenozoico , después de que el evento de extinción del Cretácico-Paleógeno hace aproximadamente 66 millones de años vaciara el espacio ecológico que alguna vez estuvo ocupado por dinosaurios no aviares y otros grupos de reptiles, así como varios otros grupos de mamíferos, [63] y experimentaron un aumento exponencial en el tamaño corporal ( megafauna ). [64] Luego, los mamíferos se diversificaron muy rápidamente; tanto las aves como los mamíferos muestran un aumento exponencial en la diversidad. [63] Por ejemplo, el murciélago más antiguo conocido data de hace unos 50 millones de años, solo 16 millones de años después de la extinción de los dinosaurios no aviares. [65]
Los estudios filogenéticos moleculares inicialmente sugirieron que la mayoría de los órdenes placentarios divergieron hace unos 100 a 85 millones de años y que las familias modernas aparecieron en el período del Eoceno tardío al Mioceno . [66] Sin embargo, no se han encontrado fósiles placentarios anteriores al final del Cretácico. [67] Los primeros fósiles indiscutibles de placentarios provienen del Paleoceno temprano , después de la extinción de los dinosaurios no aviares. [67] (Los científicos identificaron un animal del Paleoceno temprano llamado Protungulatum donnae como uno de los primeros mamíferos placentarios, [68] pero desde entonces ha sido reclasificado como un euterio no placentario). [69] Las recalibraciones de las tasas de diversidad genética y morfológica han sugerido un origen del Cretácico tardío para los placentarios y un origen del Paleoceno para la mayoría de los clados modernos. [70]
El antepasado más antiguo conocido de los primates es Archicebus achilles [71], de hace unos 55 millones de años. [71] Este diminuto primate pesaba entre 20 y 30 gramos (0,7 y 1,1 onzas) y podía caber en la palma de una mano humana. [71]
Las especies de mamíferos vivos pueden identificarse por la presencia de glándulas sudoríparas , incluidas aquellas que están especializadas en producir leche para nutrir a sus crías. [72] Sin embargo, al clasificar los fósiles, se deben utilizar otras características, ya que las glándulas de tejido blando y muchas otras características no son visibles en los fósiles. [73]
Muchos rasgos compartidos por todos los mamíferos vivos aparecieron entre los primeros miembros del grupo:
En su mayor parte, estas características no estaban presentes en los ancestros Triásicos de los mamíferos. [79] Casi todos los mamíferos poseen un hueso epipúbico, con la excepción de los placentarios modernos. [80]

En promedio, los mamíferos machos son más grandes que las hembras, siendo los machos al menos un 10% más grandes que las hembras en más del 45% de las especies investigadas. La mayoría de los órdenes de mamíferos también exhiben dimorfismo sexual sesgado hacia los machos , aunque algunos órdenes no muestran ningún sesgo o están significativamente sesgados hacia las hembras ( Lagomorpha ). El dimorfismo de tamaño sexual aumenta con el tamaño corporal en los mamíferos ( regla de Rensch ), lo que sugiere que existen presiones de selección paralelas sobre el tamaño de los machos y las hembras. El dimorfismo sesgado hacia los machos se relaciona con la selección sexual de los machos a través de la competencia macho-macho por las hembras, ya que existe una correlación positiva entre el grado de selección sexual, como lo indican los sistemas de apareamiento , y el grado de dimorfismo de tamaño sesgado hacia los machos. El grado de selección sexual también está correlacionado positivamente con el tamaño de los machos y las hembras en los mamíferos. Además, la presión de selección paralela sobre la masa de las hembras se identifica en que la edad al destete es significativamente mayor en las especies más poligínicas , incluso cuando se corrige la masa corporal. Además, la tasa reproductiva es menor para las hembras más grandes, lo que indica que la selección de la fecundidad favorece la selección de hembras más pequeñas en los mamíferos. Aunque estos patrones se mantienen en todos los mamíferos en su conjunto, existe una variación considerable entre órdenes. [81]
La mayoría de los mamíferos tienen siete vértebras cervicales (huesos del cuello). Las excepciones son el manatí y el perezoso de dos dedos , que tienen seis, y el perezoso de tres dedos, que tiene nueve. [82] Todos los cerebros de los mamíferos poseen un neocórtex , una región cerebral exclusiva de los mamíferos. [83] Los cerebros placentarios tienen un cuerpo calloso , a diferencia de los monotremas y los marsupiales. [84]
El corazón de los mamíferos tiene cuatro cámaras, dos aurículas superiores , las cámaras receptoras, y dos ventrículos inferiores , las cámaras de descarga. [85] El corazón tiene cuatro válvulas, que separan sus cámaras y aseguran que la sangre fluya en la dirección correcta a través del corazón (evitando el reflujo). Después del intercambio de gases en los capilares pulmonares (vasos sanguíneos en los pulmones), la sangre rica en oxígeno regresa a la aurícula izquierda a través de una de las cuatro venas pulmonares . La sangre fluye casi continuamente de regreso a la aurícula, que actúa como cámara receptora, y desde aquí a través de una abertura hacia el ventrículo izquierdo. La mayor parte de la sangre fluye pasivamente hacia el corazón mientras las aurículas y los ventrículos están relajados, pero hacia el final del período de relajación ventricular , la aurícula izquierda se contraerá, bombeando sangre al ventrículo. El corazón también requiere nutrientes y oxígeno que se encuentran en la sangre como otros músculos, y se suministra a través de las arterias coronarias . [86]

Los pulmones de los mamíferos son esponjosos y con forma de panal. La respiración se realiza principalmente con el diafragma , que divide el tórax de la cavidad abdominal, formando una cúpula convexa hacia el tórax. La contracción del diafragma aplana la cúpula, aumentando el volumen de la cavidad pulmonar. El aire entra por las cavidades oral y nasal, y viaja a través de la laringe, la tráquea y los bronquios , y expande los alvéolos . La relajación del diafragma tiene el efecto contrario, disminuyendo el volumen de la cavidad pulmonar, haciendo que el aire sea expulsado de los pulmones. Durante el ejercicio, la pared abdominal se contrae , aumentando la presión sobre el diafragma, lo que obliga al aire a salir más rápido y con más fuerza. La caja torácica es capaz de expandir y contraer la cavidad torácica mediante la acción de otros músculos respiratorios. En consecuencia, el aire es succionado hacia dentro o expulsado hacia fuera de los pulmones, moviéndose siempre a favor de su gradiente de presión. [87] [88] Este tipo de pulmón se conoce como pulmón de fuelle debido a su parecido con el fuelle de herrero . [88]
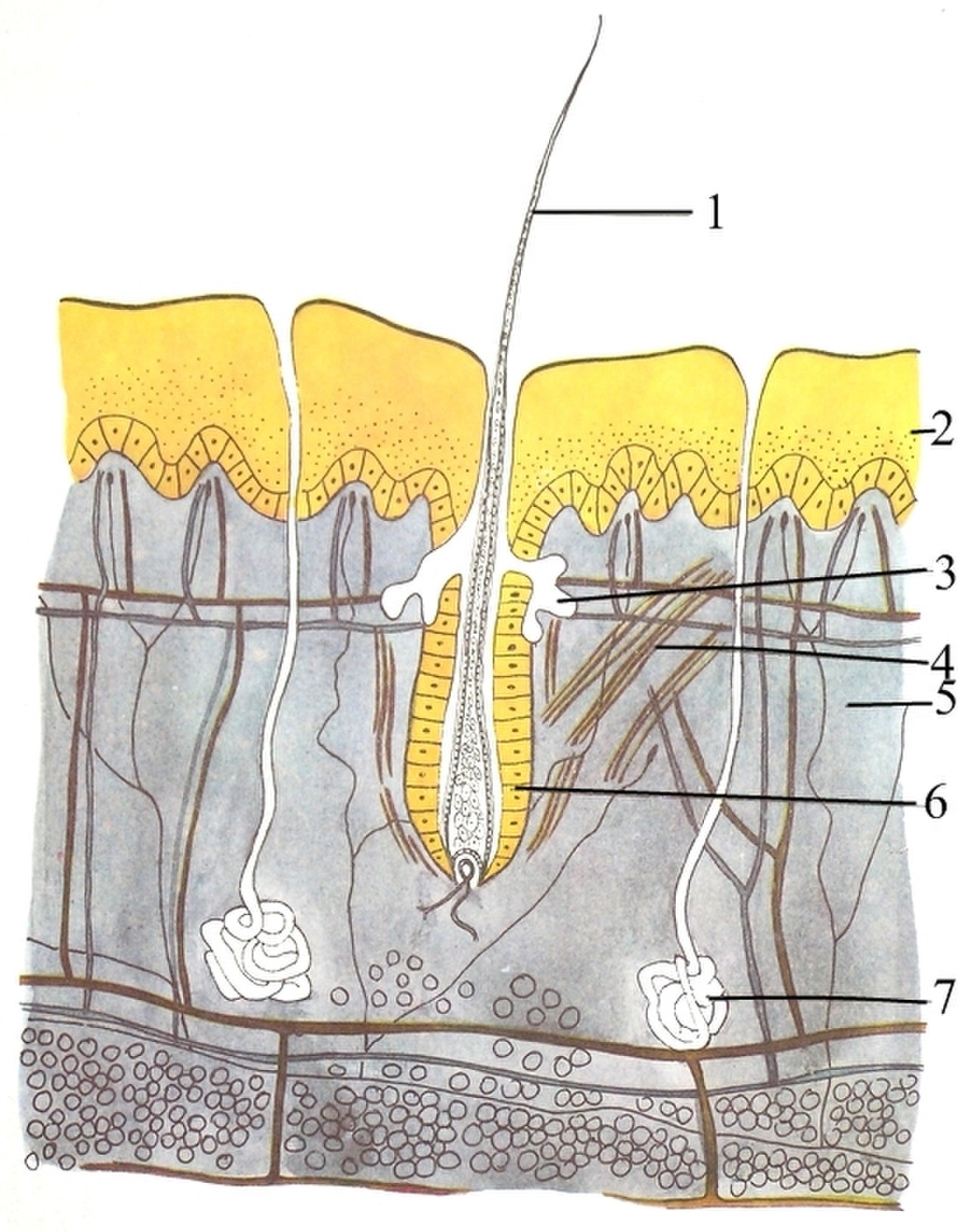
El sistema tegumentario (piel) está formado por tres capas: la epidermis más externa , la dermis y la hipodermis . La epidermis suele tener entre 10 y 30 células de grosor; su función principal es proporcionar una capa impermeable. Sus células más externas se pierden constantemente; sus células más inferiores se dividen constantemente y empujan hacia arriba. La capa intermedia, la dermis, es de 15 a 40 veces más gruesa que la epidermis. La dermis está formada por muchos componentes, como estructuras óseas y vasos sanguíneos. La hipodermis está formada por tejido adiposo , que almacena lípidos y proporciona amortiguación y aislamiento. El grosor de esta capa varía ampliamente de una especie a otra; [89] : 97 los mamíferos marinos requieren una hipodermis gruesa ( grasa ) para el aislamiento, y las ballenas francas tienen la grasa más gruesa con 20 pulgadas (51 cm). [90] Aunque otros animales tienen características como bigotes, plumas , cerdas o cilios que superficialmente se parecen a él, ningún otro animal aparte de los mamíferos tiene pelo . Es una característica definitiva de la clase, aunque algunos mamíferos tienen muy poco. [89] : 61
Los herbívoros han desarrollado una amplia gama de estructuras físicas para facilitar el consumo de material vegetal . Para romper los tejidos intactos de las plantas, los mamíferos han desarrollado estructuras dentales que reflejan sus preferencias alimentarias. Por ejemplo, los frugívoros (animales que se alimentan principalmente de fruta) y los herbívoros que se alimentan de follaje blando tienen dientes de corona baja especializados para triturar follaje y semillas . Los animales de pastoreo que tienden a comer pastos duros y ricos en sílice tienen dientes de corona alta, que son capaces de triturar tejidos vegetales duros y no se desgastan tan rápidamente como los dientes de corona baja. [91] La mayoría de los mamíferos carnívoros tienen dientes carnívoros (de longitud variable según la dieta), caninos largos y patrones de reemplazo dentario similares. [92]
El estómago de los ungulados de dedos pares (Artiodactyla) se divide en cuatro secciones: el rumen , el retículo , el omaso y el abomaso (solo los rumiantes tienen rumen). Después de consumir el material vegetal, se mezcla con saliva en el rumen y el retículo y se separa en material sólido y líquido. Los sólidos se agrupan para formar un bolo (o bolo alimenticio ) y se regurgita. Cuando el bolo entra en la boca, el líquido se exprime con la lengua y se traga nuevamente. El alimento ingerido pasa al rumen y al retículo donde los microbios celulolíticos ( bacterias , protozoos y hongos ) producen celulasa , que es necesaria para descomponer la celulosa en las plantas. [93] Los perisodáctilos , a diferencia de los rumiantes, almacenan el alimento digerido que ha salido del estómago en un ciego agrandado , donde es fermentado por bacterias. [94] Los carnívoros tienen un estómago simple adaptado para digerir principalmente carne, en comparación con los elaborados sistemas digestivos de los animales herbívoros, que son necesarios para descomponer fibras vegetales duras y complejas. El ciego está ausente o es corto y simple, y el intestino grueso no está saculado o es mucho más ancho que el intestino delgado. [95]

_(20732795545).jpg/1280px-Image_from_page_702_of_%22Outlines_of_zoology%22_(1895)_(20732795545).jpg)
El sistema excretor de los mamíferos implica muchos componentes. Como la mayoría de los demás animales terrestres, los mamíferos son ureotélicos y convierten el amoníaco en urea , lo que se realiza en el hígado como parte del ciclo de la urea . [96] La bilirrubina , un producto de desecho derivado de las células sanguíneas , pasa a través de la bilis y la orina con la ayuda de enzimas excretadas por el hígado. [97] El paso de la bilirrubina a través de la bilis a través del tracto intestinal da a las heces de los mamíferos una coloración marrón distintiva. [98] Las características distintivas del riñón de los mamíferos incluyen la presencia de la pelvis renal y las pirámides renales , y de una corteza y médula claramente distinguibles , que se debe a la presencia de asas de Henle alargadas . Solo el riñón de los mamíferos tiene forma de frijol, aunque hay algunas excepciones, como los riñones reniculados multilobulados de los pinnípedos, los cetáceos y los osos. [99] [100] La mayoría de los mamíferos placentarios adultos no tienen rastros restantes de la cloaca . En el embrión, la cloaca embrionaria se divide en una región posterior que se convierte en parte del ano, y una región anterior que tiene diferentes destinos según el sexo del individuo: en las hembras, se desarrolla en el vestíbulo o seno urogenital que recibe la uretra y la vagina , mientras que en los machos forma la totalidad de la uretra peneana . [100] [101] Sin embargo, los tenrecs , topos dorados y algunas musarañas conservan una cloaca como adultos. [102] En los marsupiales, el tracto genital está separado del ano, pero un rastro de la cloaca original permanece externamente. [100] Los monotremas, que se traduce del griego como "agujero único", tienen una cloaca verdadera. [103] La orina fluye desde los uréteres hacia la cloaca en los monotremas y hacia la vejiga en los mamíferos placentarios. [100]
Al igual que en todos los demás tetrápodos, los mamíferos tienen una laringe que puede abrirse y cerrarse rápidamente para producir sonidos, y un tracto vocal supralaríngeo que filtra este sonido. Los pulmones y la musculatura circundante proporcionan la corriente de aire y la presión necesarias para fonar . La laringe controla el tono y el volumen del sonido, pero la fuerza que ejercen los pulmones para exhalar también contribuye al volumen. Los mamíferos más primitivos, como el equidna, solo pueden silbar, ya que el sonido se logra únicamente a través de la exhalación a través de una laringe parcialmente cerrada. Otros mamíferos fonan utilizando las cuerdas vocales . El movimiento o la tensión de las cuerdas vocales puede dar lugar a muchos sonidos, como ronroneos y gritos . Los mamíferos pueden cambiar la posición de la laringe, lo que les permite respirar por la nariz mientras tragan por la boca, y formar sonidos tanto orales como nasales ; los sonidos nasales, como el gemido de un perro, son generalmente sonidos suaves, y los sonidos orales, como el ladrido de un perro, son generalmente fuertes. [104]
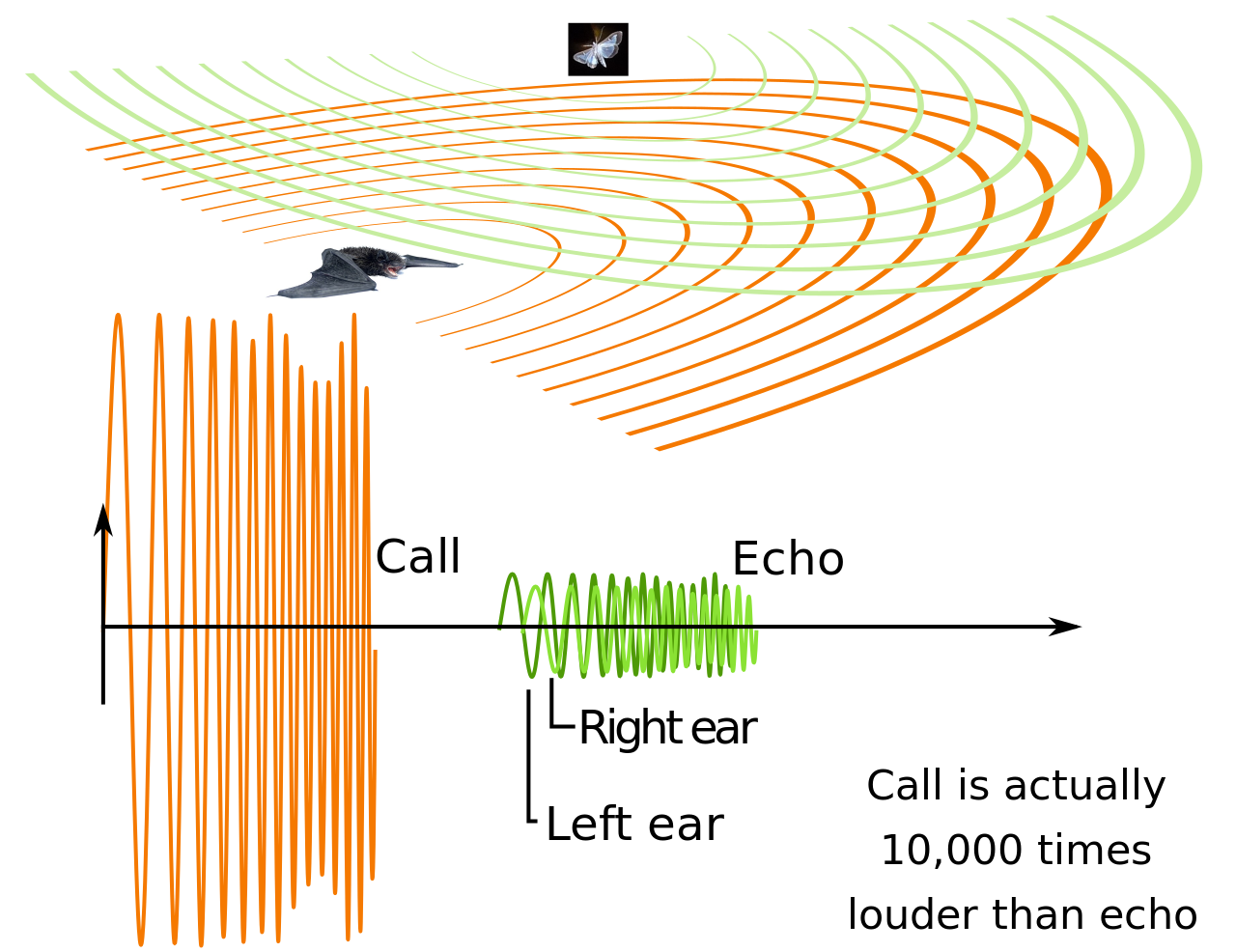
Algunos mamíferos tienen una laringe grande y, por lo tanto, una voz grave, a saber, el murciélago con cabeza de martillo ( Hypsignathus monstrosus ), donde la laringe puede ocupar la totalidad de la cavidad torácica mientras empuja los pulmones, el corazón y la tráquea hacia el abdomen . [105] Las almohadillas vocales grandes también pueden bajar el tono, como en los rugidos graves de los grandes felinos . [106] La producción de infrasonidos es posible en algunos mamíferos como el elefante africano ( Loxodonta spp.) y las ballenas barbadas . [107] [108] Los mamíferos pequeños con laringes pequeñas tienen la capacidad de producir ultrasonidos , que pueden detectarse mediante modificaciones en el oído medio y la cóclea . Los ultrasonidos son inaudibles para las aves y los reptiles, lo que podría haber sido importante durante el Mesozoico, cuando las aves y los reptiles eran los depredadores dominantes. Este canal privado es utilizado por algunos roedores, por ejemplo, en la comunicación entre madres y crías, y por los murciélagos para la ecolocalización. Las ballenas dentadas también utilizan la ecolocalización, pero, a diferencia de la membrana vocal que se extiende hacia arriba desde los pliegues vocales, tienen un melón para manipular los sonidos. Algunos mamíferos, en concreto los primates, tienen sacos de aire unidos a la laringe, que pueden funcionar para reducir las resonancias o aumentar el volumen del sonido. [104]
El sistema de producción vocal está controlado por los núcleos de los nervios craneales en el cerebro, e irrigado por el nervio laríngeo recurrente y el nervio laríngeo superior , ramas del nervio vago . El tracto vocal está irrigado por el nervio hipogloso y los nervios faciales . La estimulación eléctrica de la región gris periacueductal (PEG) del mesencéfalo de los mamíferos provoca vocalizaciones. La capacidad de aprender nuevas vocalizaciones solo se ejemplifica en humanos, focas, cetáceos, elefantes y posiblemente murciélagos; en los humanos, esto es el resultado de una conexión directa entre la corteza motora , que controla el movimiento, y las neuronas motoras en la médula espinal. [104]

La función principal del pelaje de los mamíferos es la termorregulación . Otras funciones incluyen la protección, los fines sensoriales, la impermeabilización y el camuflaje. [109] Los diferentes tipos de pelaje cumplen distintas funciones: [89] : 99
La longitud del pelo no es un factor en la termorregulación: por ejemplo, algunos mamíferos tropicales como los perezosos tienen la misma longitud de pelaje que algunos mamíferos árticos pero con menos aislamiento; y, por el contrario, otros mamíferos tropicales con pelo corto tienen el mismo valor aislante que los mamíferos árticos. La densidad del pelaje puede aumentar el valor aislante de un animal, y los mamíferos árticos especialmente tienen un pelaje denso; por ejemplo, el buey almizclero tiene pelos de protección que miden 30 cm (12 pulgadas), así como un subpelo denso, que forma una capa hermética, lo que les permite sobrevivir en temperaturas de −40 °C (−40 °F). [89] : 162–163 Algunos mamíferos del desierto, como los camellos, usan un pelaje denso para evitar que el calor solar llegue a su piel, lo que permite que el animal se mantenga fresco; el pelaje de un camello puede alcanzar los 70 °C (158 °F) en el verano, pero la piel se mantiene a 40 °C (104 °F). [89] : 188 Los mamíferos acuáticos , por el contrario, atrapan el aire en su pelaje para conservar el calor manteniendo la piel seca. [89] : 162–163

Los pelajes de los mamíferos tienen colores por diversas razones, entre las que se incluyen las principales presiones selectivas: camuflaje , selección sexual , comunicación y termorregulación. La coloración tanto del pelo como de la piel de los mamíferos está determinada principalmente por el tipo y la cantidad de melanina ; las eumelaninas para los colores marrón y negro y la feomelanina para una gama de colores amarillentos a rojizos, lo que da a los mamíferos un tono terroso . [110] [111] Algunos mamíferos tienen colores más vibrantes; ciertos monos, como los mandriles y los monos vervet , y las zarigüeyas, como las zarigüeyas ratón mexicanas y las zarigüeyas lanudas de Derby , tienen piel azul debido a la difracción de la luz en las fibras de colágeno . [112] Muchos perezosos parecen verdes porque su pelaje alberga algas verdes ; esta puede ser una relación simbiótica que proporciona camuflaje a los perezosos. [113]
El camuflaje es una influencia poderosa en un gran número de mamíferos, ya que ayuda a ocultar a los individuos de los depredadores o presas. [114] En mamíferos árticos y subárticos como el zorro ártico ( Alopex lagopus ), el lemming de collar ( Dicrostonyx groenlandicus ), el armiño ( Mustela erminea ) y la liebre de raquetas de nieve ( Lepus americanus ), el cambio de color estacional entre marrón en verano y blanco en invierno es impulsado en gran medida por el camuflaje. [115] Algunos mamíferos arbóreos, en particular los primates y marsupiales, tienen tonos de piel violeta, verde o azul en partes de sus cuerpos, lo que indica una clara ventaja en su hábitat principalmente arbóreo debido a la evolución convergente . [112]
El aposematismo , que es la forma de advertir a los posibles depredadores, es la explicación más probable del pelaje blanco y negro de muchos mamíferos que pueden defenderse, como el maloliente zorrillo y el poderoso y agresivo tejón de miel . [116] El color del pelaje a veces es sexualmente dimórfico , como en muchas especies de primates . [117] Las diferencias en el color del pelaje de las hembras y los machos pueden indicar nutrición y niveles hormonales, importantes en la selección de pareja. [118] El color del pelaje puede influir en la capacidad de retener el calor, dependiendo de la cantidad de luz que se refleja. Los mamíferos con un pelaje de color más oscuro pueden absorber más calor de la radiación solar y mantenerse más calientes, y algunos mamíferos más pequeños, como los topillos , tienen un pelaje más oscuro en invierno. El pelaje blanco y sin pigmentos de los mamíferos árticos, como el oso polar, puede reflejar más radiación solar directamente sobre la piel. [89] : 166–167 [109] Las deslumbrantes rayas blancas y negras de las cebras parecen proporcionar cierta protección contra las picaduras de moscas. [119]

Los mamíferos se reproducen por fertilización interna [120] y son únicamente gonocóricos (un animal nace con genitales masculinos o femeninos, a diferencia de los hermafroditas donde no hay tal cisma). [121] Los mamíferos machos inseminan a las hembras durante la cópula y eyaculan el semen en el tracto reproductivo femenino a través de un pene , que puede estar contenido en un prepucio cuando no está erecto. Los placentarios machos también orinan a través de un pene, y algunos placentarios también tienen un hueso del pene ( báculo ). [122] [123] [120] Los marsupiales suelen tener penes bifurcados, [124] mientras que el pene del equidna generalmente tiene cuatro cabezas con solo dos funcionando. [125] Dependiendo de la especie, una erección puede ser impulsada por el flujo sanguíneo en el tejido vascular, esponjoso o por la acción muscular. [122] Los testículos de la mayoría de los mamíferos descienden al escroto , que normalmente es posterior al pene, pero a menudo es anterior en los marsupiales. Las hembras de los mamíferos generalmente tienen una vulva ( clítoris y labios ) en el exterior, mientras que el sistema interno contiene oviductos pareados , 1-2 úteros , 1-2 cérvix y una vagina . [126] [127] Los marsupiales tienen dos vaginas laterales y una vagina medial. La "vagina" de los monotremas se entiende mejor como un "seno urogenital". Los sistemas uterinos de los mamíferos placentarios pueden variar entre un dúplex, donde hay dos úteros y cérvix que se abren a la vagina, un bipartito, donde dos cuernos uterinos tienen un solo cérvix que se conecta a la vagina, un bicorne, que consiste en dos cuernos uterinos que están conectados distalmente pero separados medialmente creando una forma de Y, y un simple, que tiene un solo útero. [128] [129] [89] : 220–221, 247

La condición ancestral para la reproducción de los mamíferos es el nacimiento de crías relativamente subdesarrolladas, ya sea a través de viviparidad directa o un corto período como huevos de cáscara blanda. Esto probablemente se debe al hecho de que el torso no podía expandirse debido a la presencia de huesos epipúbicos . La demostración más antigua de este estilo reproductivo es con Kayentatherium , que produjo perinatos no desarrollados , pero en tamaños de camada mucho más altos que cualquier mamífero moderno, 38 especímenes. [130] La mayoría de los mamíferos modernos son vivíparos y dan a luz crías vivas. Sin embargo, las cinco especies de monotremas, el ornitorrinco y las cuatro especies de equidna, ponen huevos. Los monotremas tienen un sistema de determinación sexual diferente al de la mayoría de los demás mamíferos. [131] En particular, los cromosomas sexuales de un ornitorrinco son más parecidos a los de un pollo que a los de un mamífero terio. [132]
Los mamíferos vivíparos pertenecen a la subclase Theria; los que viven hoy en día pertenecen a las infraclasas marsupiales y placentarios. Los marsupiales tienen un período de gestación corto , típicamente más corto que su ciclo estral y generalmente dan a luz a una cantidad de recién nacidos no desarrollados que luego experimentan un mayor desarrollo; en muchas especies, esto tiene lugar dentro de un saco similar a una bolsa, el marsupio , ubicado en la parte delantera del abdomen de la madre . Esta es la condición plesiomórfica entre los mamíferos vivíparos; la presencia de huesos epipúbicos en todos los mamíferos no placentarios evita la expansión del torso necesaria para un embarazo completo. [80] Incluso los euterios no placentarios probablemente se reprodujeron de esta manera. [42] Los placentarios dan a luz crías relativamente completas y desarrolladas, generalmente después de largos períodos de gestación. [133] Reciben su nombre de la placenta , que conecta al feto en desarrollo con la pared uterina para permitir la absorción de nutrientes. [134] En los mamíferos placentarios, el epipúbico se pierde completamente o se convierte en el báculo, lo que permite que el torso pueda expandirse y así dar a luz crías desarrolladas. [130]
Las glándulas mamarias de los mamíferos están especializadas en producir leche, la principal fuente de nutrición para los recién nacidos. Los monotremas se ramificaron tempranamente de otros mamíferos y no tienen las tetinas que se ven en la mayoría de los mamíferos, pero sí tienen glándulas mamarias. Las crías lamen la leche de una mancha mamaria en el vientre de la madre. [135] En comparación con los mamíferos placentarios, la leche de los marsupiales cambia mucho tanto en la tasa de producción como en la composición de nutrientes, debido al subdesarrollo de las crías. Además, las glándulas mamarias tienen más autonomía, lo que les permite suministrar leches separadas a las crías en diferentes etapas de desarrollo. [136] La lactosa es el azúcar principal en la leche de los mamíferos placentarios, mientras que la leche de los monotremas y marsupiales está dominada por oligosacáridos . [137] El destete es el proceso en el que un mamífero se vuelve menos dependiente de la leche de su madre y más de los alimentos sólidos. [138]
Casi todos los mamíferos son endotérmicos ("de sangre caliente"). La mayoría de los mamíferos también tienen pelo para ayudarlos a mantenerse calientes. Al igual que las aves, los mamíferos pueden buscar alimento o cazar en climas demasiado fríos para los reptiles e insectos ectotérmicos ("de sangre fría"). La endotermia requiere mucha energía alimentaria, por lo que los mamíferos comen más comida por unidad de peso corporal que la mayoría de los reptiles. [139] Los pequeños mamíferos insectívoros comen cantidades prodigiosas para su tamaño. Una rara excepción, la rata topo desnuda produce poco calor metabólico, por lo que se considera un poiquilotermo operativo . [140] Las aves también son endotérmicas, por lo que la endotermia no es exclusiva de los mamíferos. [141]
Entre los mamíferos, la esperanza de vida máxima de las especies varía significativamente (por ejemplo, la musaraña tiene una esperanza de vida de dos años, mientras que se registra que la ballena de Groenlandia más vieja tiene 211 años). [142] Aunque la base subyacente de estas diferencias en la esperanza de vida aún es incierta, numerosos estudios indican que la capacidad de reparar el daño del ADN es un determinante importante de la esperanza de vida de los mamíferos. En un estudio de 1974 de Hart y Setlow, [143] se encontró que la capacidad de reparación por escisión del ADN aumentaba sistemáticamente con la esperanza de vida de las especies entre siete especies de mamíferos. Se observó que la esperanza de vida de las especies estaba sólidamente correlacionada con la capacidad de reconocer roturas de doble cadena del ADN, así como con el nivel de la proteína de reparación del ADN Ku80 . [142] En un estudio de las células de dieciséis especies de mamíferos, se encontró que los genes empleados en la reparación del ADN estaban regulados positivamente en las especies de vida más larga. [144] En un estudio de 13 especies de mamíferos, se encontró que el nivel celular de la enzima de reparación del ADN, poli ADP ribosa polimerasa, se correlacionaba con la longevidad de las especies. [145] Tres estudios adicionales de una variedad de especies de mamíferos también informaron una correlación entre la longevidad de las especies y la capacidad de reparación del ADN. [146] [147] [148]

La mayoría de los vertebrados (los anfibios, los reptiles y algunos mamíferos como los humanos y los osos) son plantígrados y caminan sobre toda la parte inferior del pie. Muchos mamíferos, como los gatos y los perros, son digitígrados y caminan sobre los dedos de los pies, ya que la mayor longitud de zancada les permite una mayor velocidad. Algunos animales, como los caballos , son ungulígrados y caminan sobre las puntas de los dedos de los pies. Esto aumenta aún más la longitud de su zancada y, por lo tanto, su velocidad. [149] También se sabe que algunos mamíferos, a saber, los grandes simios, caminan sobre los nudillos , al menos en las patas delanteras. Los osos hormigueros gigantes [150] y los ornitorrincos [151] también caminan sobre los nudillos. Algunos mamíferos son bípedos y utilizan solo dos extremidades para la locomoción, lo que se puede ver, por ejemplo, en los humanos y los grandes simios. Las especies bípedas tienen un campo de visión más amplio que los cuadrúpedos, conservan más energía y tienen la capacidad de manipular objetos con las manos, lo que les ayuda a buscar alimento. En lugar de caminar, algunos bípedos saltan, como los canguros y las ratas canguro . [152] [153]
Los animales utilizan diferentes marchas para diferentes velocidades, terrenos y situaciones. Por ejemplo, los caballos muestran cuatro marchas naturales, la marcha más lenta del caballo es el paso , luego hay tres marchas más rápidas que, de más lentas a más rápidas, son el trote , el galope y el galope . Los animales también pueden tener marchas inusuales que se utilizan ocasionalmente, como para moverse de lado o hacia atrás. Por ejemplo, las principales marchas humanas son caminar bípedo y correr , pero emplean muchas otras marchas ocasionalmente, incluido el gateo de cuatro patas en espacios reducidos. [154] Los mamíferos muestran una amplia gama de marchas , el orden en que colocan y levantan sus apéndices en la locomoción. Las marchas se pueden agrupar en categorías según sus patrones de secuencia de apoyo. Para los cuadrúpedos, hay tres categorías principales: marchas de caminata, marchas de carrera y marchas de salto . [155] La marcha es el modo de andar más común, en el que algunos pies están en el suelo en un momento dado, y se encuentra en casi todos los animales con patas. Se considera que se corre cuando en algún punto de la zancada todos los pies están fuera del suelo en un momento de suspensión. [154]
.jpg/1280px-Brachiating_Gibbon_(Some_rights_reserved).jpg)
Los animales arbóreos frecuentemente tienen extremidades alargadas que los ayudan a cruzar huecos, alcanzar frutas u otros recursos, probar la firmeza del apoyo que tienen por delante y, en algunos casos, a braquiar (balancearse entre árboles). [156] Muchas especies arbóreas, como los puercoespines arbóreos, los osos hormigueros sedosos , los monos araña y las zarigüeyas , usan colas prensiles para agarrar ramas. En el mono araña, la punta de la cola tiene un parche desnudo o una almohadilla adhesiva, que proporciona mayor fricción. Las garras se pueden usar para interactuar con sustratos ásperos y reorientar la dirección de las fuerzas que aplica el animal. Esto es lo que permite a las ardillas trepar troncos de árboles que son tan grandes que son esencialmente planos desde la perspectiva de un animal tan pequeño. Sin embargo, las garras pueden interferir con la capacidad de un animal para agarrar ramas muy pequeñas, ya que pueden enrollarse demasiado y pinchar la propia pata del animal. Los primates utilizan el agarre por fricción, apoyándose en las yemas de los dedos sin pelo. Apretar la rama entre las yemas de los dedos genera una fuerza de fricción que mantiene la mano del animal pegada a la rama. Sin embargo, este tipo de agarre depende del ángulo de la fuerza de fricción, y por tanto del diámetro de la rama, y las ramas más grandes reducen la capacidad de agarre. Para controlar el descenso, especialmente por ramas de gran diámetro, algunos animales arbóreos como las ardillas han desarrollado articulaciones de tobillo muy móviles que permiten girar el pie a una postura "invertida". Esto permite que las garras se enganchen en la superficie rugosa de la corteza, oponiéndose a la fuerza de la gravedad. El tamaño pequeño proporciona muchas ventajas a las especies arbóreas: como el aumento del tamaño relativo de las ramas con respecto al animal, un centro de masa más bajo, una mayor estabilidad, una masa más baja (que permite el movimiento en ramas más pequeñas) y la capacidad de moverse a través de un hábitat más desordenado. [156] El tamaño en relación con el peso afecta a los animales que planean, como el petauro del azúcar . [157] Algunas especies de primates, murciélagos y todas las especies de perezosos consiguen una estabilidad pasiva colgándose debajo de la rama. Tanto el cabeceo como el vuelco se vuelven irrelevantes, ya que el único método de falla sería perder el agarre. [156]
Los murciélagos son los únicos mamíferos que pueden volar de verdad. Vuelan por el aire a una velocidad constante moviendo sus alas hacia arriba y hacia abajo (generalmente también con algún movimiento hacia adelante y hacia atrás). Debido a que el animal está en movimiento, hay un flujo de aire relativo a su cuerpo que, combinado con la velocidad de las alas, genera un flujo de aire más rápido que se mueve sobre el ala. Esto genera un vector de fuerza de sustentación que apunta hacia adelante y hacia arriba, y un vector de fuerza de arrastre que apunta hacia atrás y hacia arriba. Los componentes ascendentes de estos contrarrestan la gravedad, manteniendo el cuerpo en el aire, mientras que el componente delantero proporciona empuje para contrarrestar tanto el arrastre del ala como del cuerpo en su conjunto. [158]
Las alas de los murciélagos son mucho más delgadas y están formadas por más huesos que las de las aves, lo que les permite maniobrar con mayor precisión y volar con más sustentación y menos resistencia. [159] [160] Al doblar las alas hacia adentro en dirección a su cuerpo en la carrera ascendente, utilizan un 35% menos de energía durante el vuelo que las aves. [161] Las membranas son delicadas y se rasgan con facilidad; sin embargo, el tejido de la membrana del murciélago es capaz de volver a crecer, de modo que los pequeños desgarros pueden sanar rápidamente. [162] La superficie de sus alas está equipada con receptores sensibles al tacto en pequeñas protuberancias llamadas células de Merkel , que también se encuentran en las yemas de los dedos humanos. Estas áreas sensibles son diferentes en los murciélagos, ya que cada protuberancia tiene un pequeño pelo en el centro, lo que la hace aún más sensible y permite al murciélago detectar y recopilar información sobre el aire que fluye sobre sus alas, y volar de manera más eficiente cambiando la forma de sus alas en respuesta. [163]
Un fosorial (del latín fossor , que significa "cavador") es un animal adaptado a la excavación que vive principalmente, pero no exclusivamente, bajo tierra. Algunos ejemplos son los tejones y las ratas topo desnudas . Muchas especies de roedores también se consideran fosoriales porque viven en madrigueras durante la mayor parte del día, pero no todo el día. Las especies que viven exclusivamente bajo tierra son subterráneas, y aquellas con adaptaciones limitadas a un estilo de vida fosorial son subfosoriales. Algunos organismos son fosoriales para ayudar a regular la temperatura , mientras que otros usan el hábitat subterráneo para protegerse de los depredadores o para almacenar alimentos . [164]
Los mamíferos fosoriales tienen un cuerpo fusiforme, más grueso en los hombros y afinándose en la cola y la nariz. Incapaces de ver en las madrigueras oscuras, la mayoría tienen ojos degenerados, pero la degeneración varía entre especies; las tuzas de bolsillo , por ejemplo, son solo semifosoriales y tienen ojos muy pequeños pero funcionales, en el topo marsupial completamente fosorial los ojos están degenerados e inútiles, los topos talpa tienen ojos vestigiales y el topo dorado del cabo tiene una capa de piel que cubre los ojos. Las aletas auriculares externas también son muy pequeñas o están ausentes. Los mamíferos verdaderamente fosoriales tienen patas cortas y robustas, ya que la fuerza es más importante que la velocidad para un mamífero excavador, pero los mamíferos semifosoriales tienen patas cursoriales . Las patas delanteras son anchas y tienen garras fuertes para ayudar a aflojar la tierra mientras excavan madrigueras, y las patas traseras tienen membranas, así como garras, que ayudan a arrojar la tierra suelta hacia atrás. La mayoría tiene incisivos grandes para evitar que la tierra vuele hacia su boca. [165]
Muchos mamíferos fosoriales, como musarañas, erizos y topos, fueron clasificados bajo el orden ahora obsoleto Insectivora . [166]
Los mamíferos completamente acuáticos, los cetáceos y los sirénidos , han perdido sus patas y tienen una aleta caudal para impulsarse a través del agua. El movimiento de las aletas es continuo. Las ballenas nadan moviendo su aleta caudal y la parte inferior del cuerpo hacia arriba y hacia abajo, impulsándose a través del movimiento vertical, mientras que sus aletas se utilizan principalmente para dirigirse. Su anatomía esquelética les permite ser nadadores rápidos. La mayoría de las especies tienen una aleta dorsal para evitar que se den vuelta en el agua. [167] [168] Las aletas caudales de los sirénidos se elevan hacia arriba y hacia abajo en brazadas largas para mover al animal hacia adelante, y se pueden torcer para girar. Las extremidades anteriores son aletas similares a remos que ayudan a girar y reducir la velocidad. [169]
Los mamíferos semiacuáticos , como los pinnípedos, tienen dos pares de aletas en la parte delantera y trasera, las aletas delanteras y las aletas traseras. Los codos y los tobillos están encerrados dentro del cuerpo. [170] [171] Los pinnípedos tienen varias adaptaciones para reducir la resistencia . Además de sus cuerpos aerodinámicos, tienen redes suaves de haces musculares en su piel que pueden aumentar el flujo laminar y hacer que les resulte más fácil deslizarse por el agua. También carecen de erectores del pelo , por lo que su pelaje puede ser aerodinámico mientras nadan. [172] Dependen de sus aletas delanteras para la locomoción de una manera similar a las alas, similar a los pingüinos y las tortugas marinas . [173] El movimiento de las aletas delanteras no es continuo y el animal se desliza entre cada brazada. [171] En comparación con los carnívoros terrestres, las extremidades anteriores son de longitud reducida, lo que proporciona a los músculos locomotores de las articulaciones de los hombros y los codos una mayor ventaja mecánica; [170] las aletas traseras sirven como estabilizadores. [172] Otros mamíferos semiacuáticos incluyen castores, hipopótamos , nutrias y ornitorrincos. [174] Los hipopótamos son mamíferos semiacuáticos muy grandes, y sus cuerpos en forma de barril tienen estructuras esqueléticas graviportales, [175] adaptadas para soportar su enorme peso, y su gravedad específica les permite hundirse y moverse a lo largo del fondo de un río. [176]

Muchos mamíferos se comunican vocalizando. La comunicación vocal sirve para muchos propósitos, incluyendo en rituales de apareamiento, como llamadas de advertencia , [178] para indicar fuentes de alimento y para propósitos sociales. Los machos a menudo llaman durante los rituales de apareamiento para alejar a otros machos y atraer a las hembras, como en el rugido de los leones y los ciervos rojos . [179] Los cantos de la ballena jorobada pueden ser señales para las hembras; [180] tienen diferentes dialectos en diferentes regiones del océano. [181] Las vocalizaciones sociales incluyen las llamadas territoriales de los gibones y el uso de la frecuencia en los murciélagos de nariz de lanza mayor para distinguir entre grupos. [182] El mono vervet da una llamada de alarma distintiva para cada uno de al menos cuatro depredadores diferentes, y las reacciones de otros monos varían según la llamada. Por ejemplo, si una llamada de alarma señala a una pitón, los monos trepan a los árboles, mientras que la alarma del águila hace que los monos busquen un escondite en el suelo. [177] Los perros de las praderas también tienen llamadas complejas que indican el tipo, el tamaño y la velocidad de un depredador que se acerca. [183] Los elefantes se comunican socialmente con una variedad de sonidos que incluyen resoplidos, gritos, trompetas, rugidos y retumbos. Algunos de los retumbos son infrasónicos , por debajo del rango de audición de los humanos, y pueden ser escuchados por otros elefantes hasta a 6 millas (9,7 km) de distancia en horas tranquilas cerca del amanecer y el atardecer. [184]
Los mamíferos envían señales por diversos medios. Muchos dan señales visuales contra los depredadores , como cuando los ciervos y las gacelas se balancean , indicando honestamente su condición física y su capacidad para escapar, [185] [186] o cuando los ciervos de cola blanca y otros mamíferos presas se marcan la cola con marcas visibles cuando están alarmados, informando al depredador de que han sido detectados. [187] Muchos mamíferos hacen uso del marcado con olor , a veces posiblemente para ayudar a defender el territorio, pero probablemente con una variedad de funciones tanto dentro como entre especies. [188] [189] [190] Los micro murciélagos y las ballenas dentadas, incluidos los delfines oceánicos, vocalizan tanto socialmente como en ecolocalización . [191] [192] [193]
Mantener una temperatura corporal alta y constante es costoso en términos de energía, por lo que los mamíferos necesitan una dieta nutritiva y abundante. Si bien los primeros mamíferos probablemente fueron depredadores, desde entonces diferentes especies se han adaptado para satisfacer sus requisitos dietéticos de diversas maneras. Algunos comen otros animales: esta es una dieta carnívora (e incluye dietas insectívoras). Otros mamíferos, llamados herbívoros , comen plantas, que contienen carbohidratos complejos como la celulosa. Una dieta herbívora incluye subtipos como granivoría (comer semillas), folivoría (comer hojas), frugivoría (comer frutas), nectarivoría (comer néctar), gomoría (comer goma) y micofagia (comer hongos). El tracto digestivo de un herbívoro alberga bacterias que fermentan estas sustancias complejas y las hacen disponibles para la digestión, que se alojan en el estómago de múltiples cámaras o en un gran ciego. [93] Algunos mamíferos son coprófagos y consumen heces para absorber los nutrientes no digeridos cuando se ingirió el alimento por primera vez. [89] : 131–137 Un omnívoro come tanto presas como plantas. Los mamíferos carnívoros tienen un tracto digestivo simple porque las proteínas , lípidos y minerales que se encuentran en la carne requieren poca digestión especializada. Las excepciones a esto incluyen a las ballenas barbadas que también albergan la flora intestinal en un estómago de múltiples cámaras, como los herbívoros terrestres. [194]
El tamaño de un animal también es un factor para determinar el tipo de dieta ( regla de Allen ). Dado que los mamíferos pequeños tienen una alta proporción de área de superficie que pierde calor con respecto al volumen que lo genera, tienden a tener altos requerimientos de energía y una alta tasa metabólica . Los mamíferos que pesan menos de aproximadamente 18 onzas (510 g; 1,1 lb) son en su mayoría insectívoros porque no pueden tolerar el lento y complejo proceso digestivo de un herbívoro. Los animales más grandes, por otro lado, generan más calor y se pierde menos de este calor. Por lo tanto, pueden tolerar un proceso de recolección más lento (carnívoros que se alimentan de vertebrados más grandes) o un proceso digestivo más lento (herbívoros). [195] Además, los mamíferos que pesan más de 18 onzas (510 g; 1,1 lb) generalmente no pueden recolectar suficientes insectos durante sus horas de vigilia para mantenerse. Los únicos mamíferos insectívoros grandes son los que se alimentan de enormes colonias de insectos ( hormigas o termitas ). [196]
Algunos mamíferos son omnívoros y muestran distintos grados de carnivoría y herbívoría, generalmente inclinándose a favor de una más que de la otra. Dado que las plantas y la carne se digieren de forma diferente, existe una preferencia por una sobre la otra, como en el caso de los osos, donde algunas especies pueden ser principalmente carnívoras y otras principalmente herbívoras. [198] Se agrupan en tres categorías: mesocarnívoros (50-70% de carne), hipercarnívoros (70% y más de carne) e hipocarnívoros (50% o menos de carne). La dentición de los hipocarnívoros consiste en dientes carnívoros triangulares y romos destinados a moler alimentos. Los hipercarnívoros, sin embargo, tienen dientes cónicos y carnívoros afilados destinados a cortar, y en algunos casos mandíbulas fuertes para triturar huesos, como en el caso de las hienas , lo que les permite consumir huesos; algunos grupos extintos, en particular los Machairodontinae , tenían caninos en forma de sable . [197]
Algunos carnívoros fisiológicos consumen materia vegetal y algunos herbívoros fisiológicos consumen carne. Desde un aspecto conductual, esto los convertiría en omnívoros, pero desde el punto de vista fisiológico, esto puede deberse a la zoofarmacognosia . Fisiológicamente, los animales deben poder obtener tanto energía como nutrientes de materiales vegetales y animales para ser considerados omnívoros. Por lo tanto, estos animales aún pueden clasificarse como carnívoros y herbívoros cuando solo obtienen nutrientes de materiales que se originan de fuentes que aparentemente no complementan su clasificación. [199] Por ejemplo, está bien documentado que algunos ungulados como las jirafas, los camellos y el ganado, roen huesos para consumir minerales y nutrientes particulares. [200] Además, los gatos, que generalmente se consideran carnívoros obligados, ocasionalmente comen hierba para regurgitar material no digerible (como bolas de pelo ), ayudar con la producción de hemoglobina y como laxante. [201]
Muchos mamíferos, en ausencia de suficientes requerimientos alimenticios en un ambiente, suprimen su metabolismo y conservan energía en un proceso conocido como hibernación . [202] En el período que precede a la hibernación, los mamíferos más grandes, como los osos, se vuelven polífagos para aumentar las reservas de grasa, mientras que los mamíferos más pequeños prefieren recolectar y almacenar comida. [203] La desaceleración del metabolismo se acompaña de una disminución de la frecuencia cardíaca y respiratoria, así como de una caída de las temperaturas internas, que pueden rondar la temperatura ambiente en algunos casos. Por ejemplo, las temperaturas internas de las ardillas terrestres árticas que hibernan pueden bajar a -2,9 °C (26,8 °F); sin embargo, la cabeza y el cuello siempre se mantienen por encima de los 0 °C (32 °F). [204] Algunos mamíferos en ambientes cálidos estivan en épocas de sequía o calor extremo, por ejemplo, el lémur enano de cola gorda ( Cheirogaleus medius ). [205]

Por necesidad, los animales terrestres en cautiverio se acostumbran a beber agua, pero la mayoría de los animales que deambulan libremente se mantienen hidratados a través de los líquidos y la humedad de los alimentos frescos, [206] y aprenden a buscar activamente alimentos con alto contenido de líquidos. [207] Cuando las condiciones los impulsan a beber de cuerpos de agua, los métodos y movimientos difieren mucho entre las especies. [208]
Los gatos , los caninos y los rumiantes bajan el cuello y lamen el agua con sus poderosas lenguas. [208] Los gatos y los caninos lamen el agua con la lengua en forma de cuchara. [209] Los caninos lamen el agua recogiéndola en su boca con una lengua que ha tomado la forma de un cucharón. Sin embargo, con los gatos, solo la punta de su lengua (que es suave) toca el agua, y luego el gato rápidamente retrae su lengua dentro de su boca que pronto se cierra; esto da como resultado una columna de líquido que es succionada hacia la boca del gato, que luego es asegurada por su boca cerrada. [210] Los rumiantes y la mayoría de los demás herbívoros sumergen parcialmente la punta de la boca para absorber agua mediante una acción de inmersión con la lengua mantenida recta. [211] Los gatos beben a un ritmo significativamente más lento que los rumiantes, quienes enfrentan mayores peligros naturales de depredación. [208]
Muchos animales del desierto no beben incluso si hay agua disponible, sino que dependen de comer plantas suculentas . [208] En ambientes fríos y helados, algunos animales como las liebres , las ardillas arbóreas y los borregos cimarrones recurren a consumir nieve y carámbanos. [212] En las sabanas , el método de bebida de las jirafas ha sido una fuente de especulación por su aparente desafío a la gravedad; la teoría más reciente contempla que el cuello largo del animal funciona como una bomba de émbolo . [213] De manera única, los elefantes extraen agua con sus trompas y la arrojan a sus bocas. [208]En los mamíferos inteligentes, como los primates , el cerebro es más grande en relación con el resto del cerebro. La inteligencia en sí no es fácil de definir, pero los indicadores de inteligencia incluyen la capacidad de aprender, combinada con la flexibilidad conductual. Las ratas , por ejemplo, se consideran muy inteligentes, ya que pueden aprender y realizar nuevas tareas, una capacidad que puede ser importante cuando colonizan por primera vez un nuevo hábitat . En algunos mamíferos, la recolección de alimentos parece estar relacionada con la inteligencia: un ciervo que se alimenta de plantas tiene un cerebro más pequeño que un gato, que debe pensar para burlar a su presa. [196]

El uso de herramientas por parte de los animales puede indicar diferentes niveles de aprendizaje y cognición . La nutria marina utiliza rocas como parte esencial y regular de su comportamiento de búsqueda de alimento (aplastando abulones de las rocas o rompiendo conchas), y algunas poblaciones pasan el 21% de su tiempo fabricando herramientas. [214] Otros usos de herramientas, como los chimpancés que usan ramitas para "pescar" termitas, pueden desarrollarse al observar a otros usar herramientas e incluso pueden ser un verdadero ejemplo de enseñanza animal. [215] Las herramientas incluso pueden usarse para resolver acertijos en los que el animal parece experimentar un "momento Eureka" . [216] Otros mamíferos que no usan herramientas, como los perros, también pueden experimentar un momento Eureka. [217]
El tamaño del cerebro se consideraba anteriormente un indicador importante de la inteligencia de un animal. Dado que la mayor parte del cerebro se utiliza para mantener las funciones corporales, una mayor proporción de masa cerebral respecto de la masa corporal puede aumentar la cantidad de masa cerebral disponible para tareas cognitivas más complejas. El análisis alométrico indica que el tamaño del cerebro de los mamíferos se escala aproximadamente en el exponente 2 ⁄ 3 o 3 ⁄ 4 de la masa corporal. La comparación del tamaño del cerebro de un animal en particular con el tamaño del cerebro esperado en función de dicho análisis alométrico proporciona un cociente de encefalización que puede usarse como otra indicación de la inteligencia animal. [218] Los cachalotes tienen la mayor masa cerebral de cualquier animal en la Tierra, con un promedio de 8000 centímetros cúbicos (490 pulgadas cúbicas) y 7,8 kilogramos (17 libras) en los machos maduros. [219]
La autoconciencia parece ser un signo de pensamiento abstracto. Se cree que la autoconciencia, aunque no está bien definida, es un precursor de procesos más avanzados como el razonamiento metacognitivo . El método tradicional para medir esto es la prueba del espejo , que determina si un animal posee la capacidad de autorreconocimiento. [220] Los mamíferos que han pasado la prueba del espejo incluyen elefantes asiáticos (algunos pasan, otros no); [221] chimpancés; [222] bonobos ; [223] orangutanes ; [224] humanos, a partir de los 18 meses ( etapa del espejo ); [225] delfines mulares comunes ; [a] [226] orcas ; [227] y falsas orcas . [227]

La eusocialidad es el nivel más alto de organización social. Estas sociedades tienen una superposición de generaciones adultas, la división del trabajo reproductivo y el cuidado cooperativo de las crías. Por lo general, los insectos, como las abejas , las hormigas y las termitas, tienen un comportamiento eusocial, pero se demuestra en dos especies de roedores: la rata topo desnuda [228] y la rata topo de Damaraland [229] .
La presocialidad es cuando los animales exhiben más que solo interacciones sexuales con miembros de la misma especie, pero no llegan a calificar como eusociales. Es decir, los animales presociales pueden mostrar vida comunitaria, cuidado cooperativo de las crías o división primitiva del trabajo reproductivo, pero no muestran los tres rasgos esenciales de los animales eusociales. Los humanos y algunas especies de Callitrichidae ( titíes y tamarinos ) son únicos entre los primates en su grado de cuidado cooperativo de las crías. [230] Harry Harlow realizó un experimento con monos rhesus , primates presociales, en 1958; los resultados de este estudio mostraron que los encuentros sociales son necesarios para que los monos jóvenes se desarrollen tanto mental como sexualmente. [231]
Una sociedad de fisión-fusión es una sociedad que cambia frecuentemente en su tamaño y composición, formando un grupo social permanente llamado "grupo padre". Las redes sociales permanentes están formadas por todos los miembros individuales de una comunidad y a menudo varían para seguir los cambios en su entorno. En una sociedad de fisión-fusión, el grupo padre principal puede fracturarse (fisión) en subgrupos o individuos estables más pequeños para adaptarse a las circunstancias ambientales o sociales. Por ejemplo, varios machos pueden separarse del grupo principal para cazar o buscar comida durante el día, pero por la noche pueden volver a unirse (fusión) al grupo primario para compartir comida y participar en otras actividades. Muchos mamíferos exhiben esto, como los primates (por ejemplo, los orangutanes y los monos araña ), [232] elefantes, [233] hienas manchadas , [234] leones, [235] y delfines. [236]
Los animales solitarios defienden un territorio y evitan las interacciones sociales con los miembros de su especie, excepto durante la temporada de reproducción. Esto es para evitar la competencia por los recursos, ya que dos individuos de la misma especie ocuparían el mismo nicho, y para prevenir el agotamiento de los alimentos. [237] Un animal solitario, mientras busca alimento, también puede ser menos visible para los depredadores o las presas. [238]

En una jerarquía , los individuos son dominantes o sumisos. Una jerarquía despótica es donde un individuo es dominante mientras que los otros son sumisos, como en los lobos y los lémures, [239] y un orden jerárquico es una clasificación lineal de individuos donde hay un individuo superior y un individuo inferior. Los órdenes jerárquicos también pueden clasificarse por sexo, donde el individuo más bajo de un sexo tiene una clasificación más alta que el individuo superior del otro sexo, como en las hienas. [240] Los individuos dominantes, o alfas, tienen una alta probabilidad de éxito reproductivo, especialmente en harenes donde uno o unos pocos machos (machos residentes) tienen derechos exclusivos de reproducción para las hembras en un grupo. [241] Los machos no residentes también pueden ser aceptados en harenes, pero algunas especies, como el murciélago vampiro común ( Desmodus rotundus ), pueden ser más estrictas. [242]
Algunos mamíferos son perfectamente monógamos , lo que significa que se aparean de por vida y no toman otras parejas (incluso después de la muerte de la pareja original), como los lobos, los castores euroasiáticos y las nutrias. [243] [244] Hay tres tipos de poligamia: uno o varios machos dominantes tienen derechos de reproducción ( poligamia ), varios machos con los que se aparean las hembras (poliandria) o varios machos tienen relaciones exclusivas con varias hembras ( poliginandria ). Es mucho más común que ocurra el apareamiento poligínico, que, excluyendo los leks , se estima que ocurre en hasta el 90% de los mamíferos. [245] El apareamiento lek ocurre cuando los machos se congregan alrededor de las hembras y tratan de atraerlas con varias exhibiciones de cortejo y vocalizaciones, como en las focas comunes. [246]
Todos los mamíferos superiores (excluyendo a los monotremas) comparten dos adaptaciones principales para el cuidado de las crías: el nacimiento vivo y la lactancia. Esto implica una elección grupal de un grado de cuidado parental . Pueden construir nidos y cavar madrigueras para criar a sus crías, o alimentarlas y cuidarlas a menudo durante un período prolongado de tiempo. Muchos mamíferos son K-seleccionados e invierten más tiempo y energía en sus crías que los animales r-seleccionados . Cuando dos animales se aparean, ambos comparten un interés en el éxito de la descendencia, aunque a menudo en diferentes extremos. Las hembras de mamíferos muestran cierto grado de agresión maternal, otro ejemplo de cuidado parental, que puede estar dirigido contra otras hembras de la especie o las crías de otras hembras; sin embargo, algunos mamíferos pueden "tía" a las crías de otras hembras y cuidarlas. Los machos de mamíferos pueden desempeñar un papel en la crianza de los niños, como en el caso de los tenrecs, sin embargo, esto varía de una especie a otra, incluso dentro del mismo género. Por ejemplo, los machos del macaco de cola de cerdo del sur ( Macaca nemestrina ) no participan en el cuidado de los niños, mientras que los machos del macaco japonés ( M. fuscata ) sí lo hacen. [247]

Los mamíferos no humanos desempeñan una amplia variedad de funciones en la cultura humana. Son las mascotas más populares , con decenas de millones de perros, gatos y otros animales, incluidos conejos y ratones, mantenidos por familias de todo el mundo. [248] [249] [250] Los mamíferos como los mamuts , los caballos y los ciervos se encuentran entre los primeros temas de arte, y se encuentran en pinturas rupestres del Paleolítico superior , como en Lascaux . [251] Los principales artistas como Alberto Durero , George Stubbs y Edwin Landseer son conocidos por sus retratos de mamíferos. [252] Muchas especies de mamíferos han sido cazadas por deporte y como alimento; los ciervos y los jabalíes son especialmente populares como animales de caza . [253] [254] [255] Los mamíferos como los caballos y los perros son ampliamente objeto de carreras deportivas, a menudo combinadas con apuestas sobre el resultado . [256] [257] Existe una tensión entre el papel de los animales como compañeros de los humanos y su existencia como individuos con derechos propios . [258] Los mamíferos desempeñan además una amplia variedad de papeles en la literatura, [259] [260] [261] el cine, [262] la mitología y la religión. [263] [264] [265]

La domesticación de mamíferos fue fundamental para el desarrollo neolítico de la agricultura y la civilización , y provocó que los agricultores sustituyeran a los cazadores-recolectores en todo el mundo. [b] [267] Esta transición de la caza y la recolección al pastoreo de rebaños y el cultivo de cosechas fue un paso importante en la historia de la humanidad. Las nuevas economías agrícolas, basadas en mamíferos domesticados, provocaron "una reestructuración radical de las sociedades humanas, alteraciones mundiales de la biodiversidad y cambios significativos en las formas del relieve de la Tierra y su atmósfera... resultados trascendentales". [268]
Los mamíferos domésticos forman una gran parte del ganado criado para carne en todo el mundo. Incluyen (2009) alrededor de 1.400 millones de vacas , mil millones de ovejas , mil millones de cerdos domésticos , [269] [270] y (1985) más de 700 millones de conejos. [271] Los animales domésticos de trabajo, incluidos el ganado vacuno y los caballos, se han utilizado para el trabajo y el transporte desde los orígenes de la agricultura, y su número disminuyó con la llegada del transporte mecanizado y la maquinaria agrícola . En 2004 todavía proporcionaban alrededor del 80% de la energía para las granjas principalmente pequeñas del tercer mundo, y alrededor del 20% del transporte mundial, nuevamente principalmente en áreas rurales. En las regiones montañosas no aptas para vehículos con ruedas, los animales de carga continúan transportando mercancías. [272] Las pieles de mamíferos proporcionan cuero para zapatos , ropa y tapicería . La lana de mamíferos, incluidas las ovejas, las cabras y las alpacas, se ha utilizado durante siglos para la ropa. [273] [274]

Los mamíferos desempeñan un papel importante en la ciencia como animales de experimentación , tanto en la investigación biológica fundamental, como en la genética, [276] y en el desarrollo de nuevos medicamentos, que deben probarse exhaustivamente para demostrar su seguridad . [277] Millones de mamíferos, especialmente ratones y ratas, se utilizan en experimentos cada año. [278] Un ratón knockout es un ratón modificado genéticamente con un gen inactivado , reemplazado o alterado con un trozo artificial de ADN. Permiten el estudio de genes secuenciados cuyas funciones son desconocidas. [279] Un pequeño porcentaje de los mamíferos son primates no humanos, utilizados en la investigación por su similitud con los humanos. [280] [281] [282]
A pesar de los beneficios que los mamíferos domésticos han aportado al desarrollo humano, los seres humanos ejercen un efecto cada vez más perjudicial sobre los mamíferos salvajes en todo el mundo. Se ha estimado que la masa de todos los mamíferos salvajes ha disminuido a tan solo el 4% de todos los mamíferos, y que en la actualidad el 96% de los mamíferos son seres humanos y su ganado (véase la figura). De hecho, los mamíferos salvajes terrestres representan solo el 2% de todos los mamíferos. [283] [284]
Los híbridos son descendientes resultantes de la cría de dos individuos genéticamente distintos, lo que generalmente dará como resultado un alto grado de heterocigosidad, aunque híbrido y heterocigoto no son sinónimos. La hibridación deliberada o accidental de dos o más especies de animales estrechamente relacionados a través de la cría en cautiverio es una actividad humana que ha existido durante milenios y ha crecido con fines económicos. [285] Los híbridos entre diferentes subespecies dentro de una especie (como entre el tigre de Bengala y el tigre siberiano ) se conocen como híbridos intraespecíficos. Los híbridos entre diferentes especies dentro del mismo género (como entre leones y tigres) se conocen como híbridos interespecíficos o cruces. Los híbridos entre diferentes géneros (como entre ovejas y cabras) se conocen como híbridos intergenéricos. [286] Los híbridos naturales se producirán en zonas híbridas , donde dos poblaciones de especies dentro del mismo género o especie que viven en las mismas áreas o áreas adyacentes se cruzarán entre sí. Algunos híbridos han sido reconocidos como especies, como el lobo rojo (aunque esto es controvertido). [287]
La selección artificial , la crianza selectiva deliberada de animales domésticos, se está utilizando para reproducir animales recientemente extintos en un intento de lograr una raza animal con un fenotipo que se parezca a ese ancestro de tipo salvaje extinto . Un híbrido de reproducción (intraespecífico) puede ser muy similar al tipo salvaje extinto en apariencia, nicho ecológico y hasta cierto punto genética, pero el acervo genético inicial de ese tipo salvaje se pierde para siempre con su extinción . Como resultado, las razas de reproducción son, en el mejor de los casos, vagas semejanzas de los tipos salvajes extintos, como el ganado Heck lo es del uro . [288]
Las especies silvestres de raza pura que evolucionaron hasta una ecología específica pueden verse amenazadas de extinción [289] a través del proceso de contaminación genética , la hibridación incontrolada, la introgresión o la inundación genética que conduce a la homogeneización o la competencia de las especies híbridas heterosicas . [290] Cuando las personas importan o crían selectivamente nuevas poblaciones, o cuando la modificación del hábitat pone en contacto especies previamente aisladas, es posible la extinción de algunas especies, especialmente variedades raras. [291] El mestizaje puede inundar el acervo genético más raro y crear híbridos, agotando el acervo genético de raza pura. Por ejemplo, el búfalo de agua salvaje en peligro de extinción está más amenazado de extinción por la contaminación genética del búfalo de agua doméstico . Tales extinciones no siempre son evidentes desde un punto de vista morfológico . Un cierto grado de flujo genético es un proceso evolutivo normal, sin embargo, la hibridación amenaza la existencia de especies raras. [292] [293]

La pérdida de especies de las comunidades ecológicas, la defaunación , es impulsada principalmente por la actividad humana. [294] Esto ha dado como resultado bosques vacíos y comunidades ecológicas despojadas de grandes vertebrados. [295] [296] En el evento de extinción cuaternario , la muerte masiva de la variedad de megafaunal coincidió con la aparición de los humanos, lo que sugiere una influencia humana. Una hipótesis es que los humanos cazaron grandes mamíferos, como el mamut lanudo , hasta su extinción. [297] [298] El Informe de evaluación mundial de 2019 sobre diversidad biológica y servicios ecosistémicos de IPBES afirma que la biomasa total de mamíferos salvajes ha disminuido en un 82 por ciento desde el comienzo de la civilización humana. [299] [300] Los animales salvajes representan solo el 4% de la biomasa de mamíferos en la Tierra, mientras que los humanos y sus animales domésticos representan el 96%. [284]
Se predice que varias especies se extinguirán en un futuro cercano , [301] entre ellas el rinoceronte , [302] las jirafas , [303] y especies de primates [304] y pangolines . [305] Según el Informe Planeta Vivo 2020 de WWF, las poblaciones de vida silvestre vertebrada han disminuido en un 68% desde 1970 como resultado de las actividades humanas, en particular el consumo excesivo , el crecimiento de la población y la agricultura intensiva , lo que es evidencia de que los humanos han desencadenado un sexto evento de extinción masiva. [306] [307] La caza por sí sola amenaza a cientos de especies de mamíferos en todo el mundo. [308] [309] Los científicos afirman que la creciente demanda de carne está contribuyendo a la pérdida de biodiversidad , ya que este es un factor importante de la deforestación y la destrucción del hábitat ; los hábitats ricos en especies, como porciones significativas de la selva amazónica , se están convirtiendo en tierras agrícolas para la producción de carne. [310] [311] [312] Otra influencia es la caza excesiva y la caza furtiva , que pueden reducir la población general de animales de caza, [313] especialmente aquellos ubicados cerca de aldeas, [314] como en el caso de los pecaríes . [315] Los efectos de la caza furtiva se pueden ver especialmente en el comercio de marfil con elefantes africanos. [316] Los mamíferos marinos corren el riesgo de enredarse en las artes de pesca, en particular los cetáceos , con mortalidades por descarte que van de 65.000 a 86.000 individuos al año. [317]
Se está prestando atención a las especies en peligro de extinción a nivel mundial, en particular a través del Convenio sobre la Diversidad Biológica , también conocido como el Acuerdo de Río, que incluye 189 países signatarios que se centran en la identificación de especies y hábitats en peligro de extinción. [318] Otra organización de conservación notable es la UICN, que tiene una membresía de más de 1.200 organizaciones gubernamentales y no gubernamentales . [319]
Las extinciones recientes pueden atribuirse directamente a influencias humanas. [320] [294] La UICN caracteriza las extinciones "recientes" como aquellas que han ocurrido más allá del punto de corte de 1500, [321] y alrededor de 80 especies de mamíferos se han extinguido desde ese momento y 2015. [322] Algunas especies, como el ciervo del Père David [323] están extintas en la naturaleza y sobreviven únicamente en poblaciones cautivas. Otras especies, como la pantera de Florida , están ecológicamente extintas , sobreviviendo en números tan bajos que esencialmente no tienen impacto en el ecosistema. [324] : 318 Otras poblaciones solo están extintas localmente (extirpadas), aún existen en otros lugares, pero su distribución está reducida, [324] : 75–77 como con la extinción de las ballenas grises en el Atlántico . [325]
{{cite journal}}: CS1 maint: DOI inactive as of September 2024 (link){kind=link}