Cilio
[18] Significativamente, estos organismos tampoco poseen las tubulinas "especiales" (δ, ε, ζ y η) que permiten organizar el centriolo.
[19] En vertebrados, prácticamente todos los tipos celulares tienen cilios primarios o proceden de células que los tuvieron durante su desarrollo embrionario.
Los cilios sensoriales contribuyen al reconocimiento de individuos compatibles en el apareamiento de protistas, reconocimiento y anclaje al hospedador en los protistas parásitos, geotaxia en los moluscos, mecanorrecepción en artrópodos, y en la quimiorrecepción en los vertebrados.
[24] A nivel macroscópico, existen pruebas diagnósticas que permiten verificar la función ciliar con fines clínicos.
La inmunofluorescencia se emplea especialmente en la inmunolocalización de algún componente concreto del cilio, o bien para trazar el transporte intraflagelar.
Esto se puede efectuar confinándolas entre dos vidrios con una separación estrecha, procurando no distorsionar las estructuras celulares.
Los primeros estudios con microscopía electrónica lograron determinar las características ultraestructurales del axonema y otros elementos de la estructura ciliar.
Una nueva técnica, la tomografía crioelectrónica, en la que las muestras son vitrificadas a temperaturas muy bajas (algunos microscopios son enfriados con helio líquido) y posteriormente fotografiadas desde todos los ángulos, reconstruyéndose posteriormente una imagen tridimensional de la estructura con una resolución que alcanza los 2 nm.
Una ventaja adicional de esta técnica debido a la rápida congelación, es que permite detener la célula o los complejos proteicos en un estado particular.
[35] Algunos elementos celulares, como los estereocilios pueden confundirse con los cilios al microscopio óptico, pero en realidad están estructuralmente relacionados con las microvellosidades.
[36] Las cilias son orgánulos cortos que están rodeados por membrana plasmática en su porción extracelular.
Muestran una forma cilíndrica, de diámetro uniforme en toda su longitud, con una terminación libre redondeada, semiesférica.
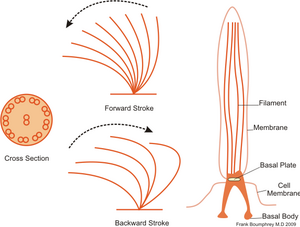
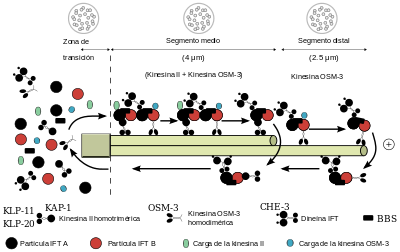
Estos orgánulos están dotados de un armazón compleja, basada en microtúbulos y que es denominada axonema.
El axonema se origina en la base del cilio y por debajo de la membrana plasmática, en una estructura compleja denominada cuerpo basal.
Algunos estudios indican que al menos, en algunos casos, la fluidez de la membrana puede responder a señales como el ATP extracelular.
Es especialmente importante en las células fotorreceptoras, que como se dijo anteriormente son cilios especializados.
[42] En esta región se observa a veces una depresión de la membrana plasmática llamada "caveola".
[5] Aunque se extiende y es continua con la membrana plasmática, existen notables diferencias en la composición, en especial, de proteínas.
Todo ello permitiría dirigir el impulso de los cilios a las condiciones más favorables.
[48] También en muchas ocasiones puede observarse en la parte más alta del cilio un número variable (3-7) de púas.
Algunas funciones que se le suponen son el ensamblaje de microtúbulos, movimiento ciliar y transporte mucociliar.
[49] En Trypanosoma se ha comprobado que la extensión de la membrana flagelar no depende del transporte intraciliar.
[53] La distinción entre estos cilias y flagelos se basa principalmente en su tamaño (unos 12-14 μm), número por célula (suelen ser muchos, con excepción de los cilios primarios y nodales,[14] mientras que los flagelos uno o dos) y en su caso, por el patrón de movimiento (los cilios baten como un remo, son inmóviles o crean un vórtice, mientras que los flagelos no ondulan).
Correspondiendo con estas diferencias estructurales, también existen diferencias funcionales: los flagelos pueden propulsar células móviles en un líquido, mientras que los cilios se sitúan normalmente en células estacionarias, y gracias a su impulso mueven líquidos o elementos contenidos en él.
[56] El que sigue en fila halla así una dirección favorecida y se mueve por ella con un pequeño retraso, conducta llamada metacronismo.
Finalmente, esta estructura se propaga y crecen hasta adquirir la disposición habitual de 9+0 tripletes.
Además, cada complejo parecen unidas por fibras entre sí, con la membrana plasmática y con los túbulos B de los dobletes externos.
[67] Se ha propuesto que el ciclo completo tiene lugar en seis pasos:[68] Existen diversas teorías sobre el origen de los undulipodia en eucariotas, a los que en los análisis evolutivos se les suele llamar "cilios" conjuntamente para no crear confusión con los flagelos bacterianos.
El undulipodio permanecería unido al núcleo por un rizoplasto y un aparato parabasal (Golgi).
Se predice que la tectina y otras proteínas "en cinta" serían aportadas por el virus hipotético.