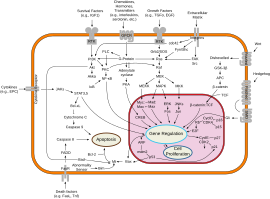
Transducción de señal
Cada proceso se realiza en intervalos de tiempo muy pequeños, como milisegundos, o en periodos más largos como algunos segundos.
Así las células modulan todas sus funciones, desde las más generales: replicación, crecimiento, diferenciación o maduración, apoptosis, etc.; hasta otras funciones más finas: contracción, secreción, meiosis, etc.
Las células que forman los organismos multicelulares responden a una gran cantidad de estímulos químicos.
Unos, como los neurotransmisores, las hormonas y los factores de crecimiento, son producidos por las propias células del organismo y alcanzan a las células diana a través del medio interno.
Otros, aunque también alcanzan a las células a través del medio interno proceden del exterior como el oxígeno, un gran número de nutrientes, estímulos olfatorios y gustatorios que generan respuestas específicas en ciertos grupos celulares.
Esta convergencia en unas pocas cadenas de transducción comunes en plantas y animales ocurre en primer lugar en los receptores celulares.
Tal conjunto de activación a menudo se llama programa genético.
Estas sustancias de señalización externa se sitúan en un lugar del receptor y provocan un cambio en la superficie o conformación espacial del mismo; que ocurre cuando la molécula de señalización se une al receptor.
Existen muchas moléculas que pueden funcionar como portadoras extracelulares de información.
A continuación se describirán: A menudo, pero no siempre, los eventos intracelulares activados por las señales externas son considerados desde el punto de vista de transducción en sí mismo, el cual en sentido estricto se refiere solo al paso que convierte la señal extracelular en señal intracelular.
Las moléculas de señalización intracelular en células eucariotas incluyen proteínas G heterotriméricas, pequeñas GTP-asas, nucleótidos cíclicos como AMP cíclico (AMPc) y GMP cíclico (GMPc), ion calcio, derivados fosfoinositoles como fosfatidilinositol 4,5-bifosfato (PIP2), diacilglicerol (DAG) e inositoltrifosfato (IP3) y varias proteínas quinasas y fosfatasas.
La comunicación intercelular está unida a señales extracelulares y esto ocurre en organismos complejos que están formados por muchas células.
En este caso, la hormona no atraviesa ella misma la membrana plasmática para penetrar en la célula.
La equivalencia entre la unión hormona-receptor y la hormona libre es igual a: [H] + [R] <-> [HR], con
Primero, el receptor tiene que reconocer la hormona con su dominio extracelular, después activar a otras proteínas situadas en el citosol por medio de su dominio citoplasmático, gracias a un cambio de conformación proteico.
Por medio de una modificación postraduccional como miristilación, pamitorilación, farnesilación, geranilación y unión a glucosil-fosfatidilinositol, muchas proteínas asociadas a membrana pueden ser activadas por turnos o estar juntas formando un complejo multiproteico que finalmente envía la señal por medio de moléculas solubles al interior de la célula.
Cuando se une el ligando, el receptor se dimeriza, lo que induce la autofosforilación de las tirosinas del dominio intracelular y activa la tirosina quinasa, que fosforila (y por tanto activa) muchas moléculas efectoras en cascada, de forma directa o mediante proteínas adaptadoras.
Estos receptores pueden activar cascadas de señalización diferentes, como por ejemplo: En muchos tipos de cáncer se han detectado alteraciones en la actividad tirosina quinasa del receptor y mutaciones, por lo que estas moléculas son dianas terapéuticas muy importantes.
Estas quinasas activan factores de transcripción citoplásmicos llamados STATs (por signal transducers and activation of transcription), que se translocan al núcleo y activan la transcripción de genes específicos.
Hay un gran número de ligandos que utilizan estos receptores, como las quimiokinas, vasopresina, serotonina, histamina, adrenalina, noradrenalina, calcitonina, glucagón y hormona paratiroidea, entre otros.
La unión del ligando provoca cambio de conformación y activación del receptor, que interacciona continuamente con tantas proteínas G como pueda durante todo el tiempo que tiene unido el ligando, haciendo que estas intercambien el GDP por GTP en la subunidad α, permitiendo así la disociación de la proteína en sus dos componentes activos: el complejo α-GTP y el complejo β-γ, ambos con actividad reguladora.
Un canal iónico activado por un ligando puede reconocer otras moléculas, y después de sufrir un cambio estructural se abre un canal en la membrana plasmática a través del cual pueden pasar determinados iones.
Los detalles de la regulación genética todavía no son del todo conocidos.
Las secuencias de ADN que hacen juego con el receptor son normalmente repetición hexaméricas, tanto invertidas como evertidas.
Cuando el calcio es liberado por el IP3 que abre canales de calcio (es importante tener en cuenta que el IP3 no es un segundo mensajero) y por lo tanto es activo y junto con el diacilglicerol activan proteínas quinasas C, actúa en un espacio muy limitado de tiempo.
El complejo calcio-calmodulina ejerce una función importante en la proliferación, mitosis y transducción de señal neuronal.
El NO se forma a partir de la arginina y el oxígeno por la enzima óxido nítrico sintasa, con citrulina como producto.
El NO funciona principalmente a través de receptores diana, la enzima soluble guanilato ciclasa, que cuando se activa produce el segundo mensajero guanosín monofosfato cíclico (GMPc).