Nicotinamida adenina dinucleótido
El dinucleótido de nicotinamida y adenina, también conocido como nicotin adenin dinucleótido o nicotinamida adenina dinucleótido (abreviado NAD+ en su forma oxidada y NADH en su forma reducida), es una coenzima que se halla en las células vivas y que está compuesta por un dinucleótido, es decir, por dos nucleótidos, unidos a través de grupos fosfatos: uno de ellos es una base de adenina y el otro, una nicotinamida.
Debido a esto, la coenzima se encuentra en dos formas: como un agente oxidante, que acepta electrones de otras moléculas.
[4] Notificaron que al adherir extracto de levadura hervido y filtrado, se aceleraba en gran medida la fermentación alcohólica en extractos de levadura sin hervir.
Cofermento fue como ellos llamaron al factor no identificado responsable de este efecto.
[8] A principios de la década de 1940, Arthur Kornberg realizó otra importante contribución para el avance hacia la comprensión del metabolismo del NAD+, pues fue el primer científico en detectar una enzima en la ruta biosintética.
[9] Subsecuentemente, en 1949, los bioquímicos estadounidenses Morris Friedkin y Albert L. Lehninger descubrieron que el NADH se encontraba enlazado a rutas metabólicas como el ciclo del ácido cítrico con la síntesis de ATP en la fosforilación oxidativa.
Las funciones no redox del NAD y el NADP son un reciente descubrimiento.
[18] La reacción es fácilmente reversible cuando el NADH reduce otra molécula y es re-oxidada a NAD+.
Esto significa que esta coenzima puede permanecer continuamente en un ciclo entre sus formas de NAD+ y NADH sin ser consumida.
[20] Tanto la NAD+ como el NADH absorben fuertemente en la banda ultravioleta debido a la base de adenina.
[28] Los datos sobre otros compartimentos celulares son limitados, aunque en la mitocondria la concentración de NAD+ es similar a la del citosol.
[29] El balance entre la forma oxidada y reducida de la nicotinamida adenina dinucleótido se llama proporción NAD+/NADH.
[34] Estas distintas proporciones son la clave para los diferentes roles metabólicos del NADH y el NADPH.
[37] En la mayoría de los organismos, esta enzima utiliza ATP como fuente del grupo fosfato, aunque en algunas bacterias, como la Mycobacterium tuberculosis y algunas arqueobacterias hipertermofílas como la Pyrococcus horikoshii del género Pyrococcus, usan polifosfatos inorgánicos como un donante alternativo de fosforilo.
[45][46] Aún más sorprendente es el patógeno intracelular Chlamydia trachomatis, que carece de candidatos reconocibles para cualquiera de los genes implicados en la biosíntesis o el rescate tanto del NAD+ como del NADP+, por lo cual debe adquirir estas coenzimas a partir su hospedante.
[52] Este plegamiento contiene tres o más láminas beta paralelas unidas por dos hélices alfa en orden beta-alfa-beta-alfa-beta.
Esto forma una hoja beta flanqueada por una capa de hélices alfa a cada lado.
[54] A pesar de esta similitud en la forma en que las proteínas se unen a las dos coenzimas, las enzimas casi siempre muestran un alto nivel de especificidad, ya sea por el NAD+ o el NADP+.
Sin embargo, hay algunas pocas excepciones a esta regla general, y enzimas como la aldosa reductasa, Glucosa-6-fosfato deshidrogenasa (G6FD), y la metilentetrahidrofolato reductasa pueden utilizar ambas coenzimas en algunas especies.
En células eucariotas, los electrones transportados por el NADH que se produce en el citoplasma por glucólisis, son transferidos al interior de la mitocondria (para reducir el NAD+ mitocondrial) por lanzaderas mitocondriales, como la lanzadera malato-aspartato.
Puesto que el NADPH es necesario para conducir las reacciones redox como un fuerte agente reductor, la proporción NADP+/NADPH se mantiene muy baja.
Por ejemplo, las bacterias nitrificantes como Nitrobacter oxidan el nitrito a nitrato, lo que libera energía suficiente para bombear los protones y generar ATP, pero no para producir NADH directamente.
[65] La ADP-ribosilación implica ya sea la adición de un solo grupo ADP-ribosa; en mono-ADP-ribosilación, o la transferencia de ADP-ribosa a las proteínas en cadenas largas ramificadas, que es llamada poli(ADP-ribosil)ación.
[66][69] La estructura de la poli-(ADP-ribosa) está implicada en la regulación de varios eventos celulares, y es la más importante en el núcleo celular, actuando en procesos como la reparación del ADN y el mantenimiento del telómero.
[75] Sin embargo, las proteínas no histonas pueden ser así mismo desacetilizadas por las sirtuinas.
Estas actividades de las sirtuinas son particularmente interesantes debido a su importancia en la regulación del envejecimiento.
Este intermediario es entonces atacado por el grupo hidroxilo 3' del otro extremo de ADN, formando un nuevo enlace fosfodiéster.
[77] Esto contrasta con las ADN ligasas eucariontes, que utilizan ATP para formar el intermediario ADN-AMP.
La Isoniazida es un profármaco y una vez que ha entrado en la bacteria, es activada por una peroxidasa, la cual oxida el compuesto en su forma de radical libre.
[92][93] Por ejemplo, la enzima nicotinamidasa, que convierte la nicotinamida en ácido nicotínico, es un blanco para el diseño de fármacos, puesto que esta enzima está ausente en humanos, pero presente en hongos unicelulares y en bacterias.



y la NADH.
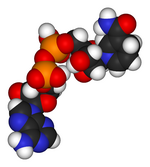

en los vertebrados . Las abreviaciones están definidas en el texto.

.

en rojo, las beta-láminas en amarillo, y las hélices alfa en morado. [ 48 ]

.

y el NADH enlazan el ciclo de Krebs y la fosforilación oxidativa .
