
El parasitismo es una relación estrecha entre especies , donde un organismo, el parásito, vive sobre o dentro de otro organismo, el huésped , causándole algún daño, y está adaptado estructuralmente a esta forma de vida. [1] El entomólogo EO Wilson caracterizó a los parásitos como "depredadores que comen presas en unidades de menos de uno". [2] Los parásitos incluyen protozoos unicelulares como los agentes de la malaria , la enfermedad del sueño y la disentería amebiana ; animales como anquilostomas , piojos , mosquitos y murciélagos vampiros ; hongos como el hongo de la miel y los agentes de la tiña ; y plantas como el muérdago , la cuscuta y las retamas .
Existen seis estrategias parasitarias principales de explotación de huéspedes animales, a saber: castración parasitaria , parasitismo transmitido directamente (por contacto), parasitismo transmitido tróficamente (al ser ingerido), parasitismo transmitido por vectores , parasitoidismo y microdepredación. Un eje principal de clasificación se refiere a la invasividad: un endoparásito vive dentro del cuerpo del huésped; un ectoparásito vive fuera, en la superficie del huésped.
Al igual que la depredación, el parasitismo es un tipo de interacción entre consumidor y recurso , [3] pero a diferencia de los depredadores , los parásitos, con excepción de los parasitoides, son mucho más pequeños que sus huéspedes, no los matan y a menudo viven en sus huéspedes o sobre ellos durante un período prolongado. Los parásitos de los animales son altamente especializados , cada especie de parásito vive en una especie animal determinada y se reproduce a un ritmo más rápido que sus huéspedes. Los ejemplos clásicos incluyen interacciones entre huéspedes vertebrados y tenias , duelas y aquellas entre las especies de Plasmodium causantes de malaria y pulgas .
Los parásitos reducen la aptitud del hospedador mediante una patología general o especializada , que va desde la castración parasitaria hasta la modificación del comportamiento del hospedador . Los parásitos aumentan su propia aptitud explotando a los hospedadores para obtener los recursos necesarios para su supervivencia, en particular alimentándose de ellos y utilizando hospedadores intermediarios (secundarios) para ayudar en su transmisión de un hospedador definitivo (primario) a otro. Aunque el parasitismo suele ser inequívoco, es parte de un espectro de interacciones entre especies , que se gradúa a través del parasitoidismo en depredación, a través de la evolución en mutualismo y, en algunos hongos, en ser saprofito .
El conocimiento humano de parásitos como lombrices intestinales y tenias se remonta al antiguo Egipto , Grecia y Roma . En los primeros tiempos modernos , Antonie van Leeuwenhoek observó Giardia lamblia con su microscopio en 1681, mientras que Francesco Redi describió parásitos internos y externos, incluyendo la duela del hígado de las ovejas y las garrapatas . La parasitología moderna se desarrolló en el siglo XIX. En la cultura humana, el parasitismo tiene connotaciones negativas. Estas fueron explotadas con efecto satírico en el poema de Jonathan Swift de 1733 "Sobre la poesía: una rapsodia", comparando a los poetas con "alimañas" hiperparasitarias . En la ficción, la novela de terror gótico de Bram Stoker de 1897 Drácula y sus muchas adaptaciones posteriores presentaron un parásito bebedor de sangre . La película Alien de Ridley Scott de 1979 fue una de las muchas obras de ciencia ficción que presentaron una especie alienígena parásita. [4]
Utilizada por primera vez en inglés en 1539, la palabra parásito proviene del francés medieval parasite , de la forma latinizada parasitus , del griego antiguo παράσιτος [5] (parasitos) 'el que come en la mesa de otro' a su vez de παρά [6] (para) 'al lado, por' y σῖτος (sitos) 'trigo, comida'. [7] El término relacionado parasitismo aparece en inglés a partir de 1611. [8]
El parasitismo es un tipo de simbiosis , una interacción biológica estrecha y persistente a largo plazo entre un parásito y su huésped. A diferencia de los saprótrofos , los parásitos se alimentan de huéspedes vivos, aunque algunos hongos parásitos, por ejemplo, pueden seguir alimentándose de los huéspedes que han matado. A diferencia del comensalismo y el mutualismo , la relación parasitaria daña al huésped, ya sea alimentándose de él o, como en el caso de los parásitos intestinales, consumiendo parte de su alimento. Debido a que los parásitos interactúan con otras especies, pueden actuar fácilmente como vectores de patógenos, causando enfermedades . [9] [10] [11] La depredación , por definición, no es una simbiosis, ya que la interacción es breve, pero el entomólogo EO Wilson ha caracterizado a los parásitos como "depredadores que comen presas en unidades de menos de uno". [2]
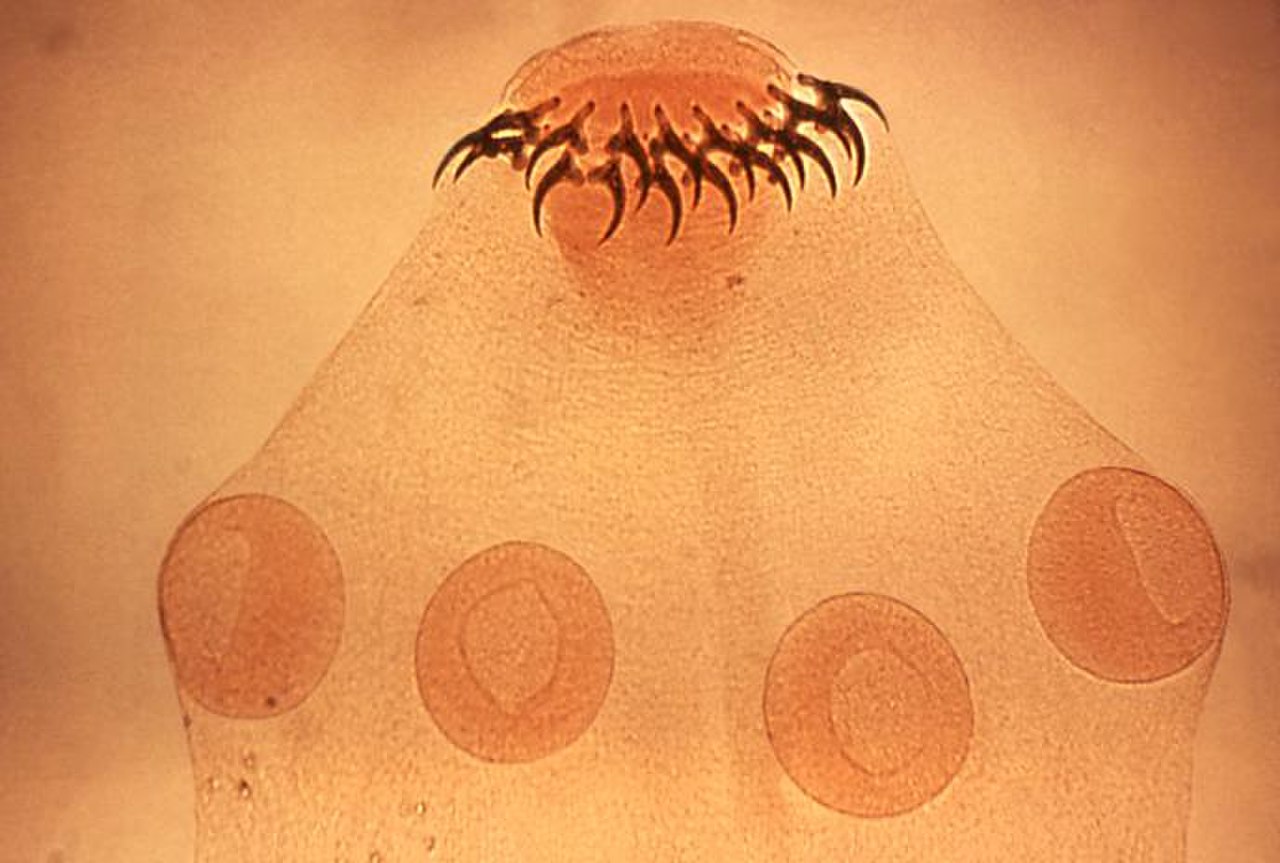
Dentro de ese ámbito hay muchas estrategias posibles. Los taxónomos clasifican a los parásitos en una variedad de esquemas superpuestos, basados en sus interacciones con sus hospedadores y en sus ciclos de vida , que a veces son muy complejos. Un parásito obligado depende completamente del hospedador para completar su ciclo de vida, mientras que un parásito facultativo no. Los ciclos de vida de los parásitos que involucran solo a un hospedador se denominan "directos"; aquellos con un hospedador definitivo (donde el parásito se reproduce sexualmente) y al menos un hospedador intermediario se denominan "indirectos". [12] [13] Un endoparásito vive dentro del cuerpo del hospedador; un ectoparásito vive afuera, en la superficie del hospedador. [14] Los mesoparásitos, como algunos copépodos , por ejemplo, ingresan a una abertura en el cuerpo del hospedador y permanecen parcialmente incrustados allí. [15] Algunos parásitos pueden ser generalistas y alimentarse de una amplia gama de hospedadores, pero muchos parásitos, y la mayoría de los protozoos y helmintos que parasitan animales, son especialistas y extremadamente específicos de hospedadores. [14] Una primera división básica y funcional de los parásitos distinguía a los microparásitos y los macroparásitos. A cada uno de ellos se le asignaba un modelo matemático para analizar los movimientos poblacionales de los grupos hospedador-parásito. [16] Los microorganismos y virus que pueden reproducirse y completar su ciclo de vida dentro del hospedador se conocen como microparásitos. Los macroparásitos son los organismos multicelulares que se reproducen y completan su ciclo de vida fuera del hospedador o en el cuerpo de este. [16] [17]
Gran parte del pensamiento sobre los tipos de parasitismo se ha centrado en los parásitos de animales terrestres, como los helmintos. Aquellos en otros entornos y con otros huéspedes a menudo tienen estrategias análogas. Por ejemplo, la anguila chata es probablemente un endoparásito facultativo (es decir, es semiparásito) que oportunistamente excava y se come a los peces enfermos y moribundos. [18] Los insectos herbívoros como las cochinillas , los pulgones y las orugas se parecen mucho a los ectoparásitos, atacando plantas mucho más grandes; sirven como vectores de bacterias, hongos y virus que causan enfermedades de las plantas . Como las cochinillas hembras no pueden moverse, son parásitos obligados, permanentemente adheridos a sus huéspedes. [16]
Las señales sensoriales que utiliza un parásito para identificar y acercarse a un huésped potencial se conocen como "señales del huésped". Estas señales pueden incluir, por ejemplo, vibración, [19] dióxido de carbono exhalado , olores de la piel, señales visuales y térmicas y humedad. [20] Las plantas parásitas pueden utilizar, por ejemplo, la luz, la fisioquímica del huésped y los compuestos volátiles para reconocer a los huéspedes potenciales. [21]
Existen seis estrategias parasitarias principales , a saber: castración parasitaria ; parasitismo transmitido directamente; parasitismo transmitido tróficamente; parasitismo transmitido por vectores ; parasitoidismo ; y microdepredación. Estas se aplican a parásitos cuyos huéspedes son plantas y animales. [16] [22] Estas estrategias representan picos adaptativos ; son posibles estrategias intermedias, pero los organismos de muchos grupos diferentes han convergido consistentemente en estas seis, que son evolutivamente estables. [22]
Se puede obtener una perspectiva de las opciones evolutivas considerando cuatro cuestiones clave: el efecto sobre la aptitud de los hospedadores de un parásito; el número de hospedadores que tiene por etapa de vida; si se impide que el hospedador se reproduzca; y si el efecto depende de la intensidad (número de parásitos por hospedador). De este análisis, surgen las principales estrategias evolutivas del parasitismo, junto con la depredación. [23]

Los castradores parásitos destruyen parcial o totalmente la capacidad de su anfitrión para reproducirse, desviando la energía que se habría destinado a la reproducción hacia el crecimiento del anfitrión y del parásito, a veces causando gigantismo en el anfitrión. Los otros sistemas del anfitrión permanecen intactos, lo que le permite sobrevivir y sostener al parásito. [22] [24] Los crustáceos parásitos como los del género especializado Sacculina , los percebes , causan específicamente daños a las gónadas de sus muchas especies [25] de cangrejos anfitriones . En el caso de Sacculina , los testículos de más de dos tercios de sus cangrejos anfitriones se degeneran lo suficiente para que estos cangrejos machos desarrollen características sexuales secundarias femeninas , como abdómenes más anchos, pinzas más pequeñas y apéndices para agarrar huevos. Varias especies de helmintos castran a sus anfitriones (como insectos y caracoles). Esto puede suceder directamente, ya sea mecánicamente alimentándose de sus gónadas o secretando una sustancia química que destruye las células reproductivas; o indirectamente, ya sea secretando una hormona o desviando nutrientes. Por ejemplo, el trematodo Zoogonus lasius , cuyos esporocistos carecen de boca, castra químicamente al caracol marino intermareal Tritia obsoleta , desarrollándose en su gónada y matando sus células reproductivas. [24] [26]

Los parásitos de transmisión directa, que no requieren un vector para llegar a sus huéspedes, incluyen parásitos de vertebrados terrestres como piojos y ácaros; parásitos marinos como copépodos y anfípodos cíamidas ; monogeneos ; y muchas especies de nematodos, hongos, protozoos, bacterias y virus. Ya sean endoparásitos o ectoparásitos, cada uno tiene una sola especie huésped. Dentro de esa especie, la mayoría de los individuos están libres o casi libres de parásitos, mientras que una minoría porta una gran cantidad de parásitos; esto se conoce como distribución agregada . [22]

Los parásitos de transmisión trófica se transmiten al ser ingeridos por un huésped. Incluyen trematodos (todos excepto esquistosomas ), cestodos , acantocéfalos , pentastómidos , muchos gusanos redondos y muchos protozoos como Toxoplasma . [22] Tienen ciclos de vida complejos que involucran huéspedes de dos o más especies. En sus etapas juveniles infectan y a menudo se enquistan en el huésped intermediario. Cuando el animal huésped intermediario es ingerido por un depredador, el huésped definitivo, el parásito sobrevive al proceso de digestión y madura hasta convertirse en un adulto; algunos viven como parásitos intestinales . Muchos parásitos de transmisión trófica modifican el comportamiento de sus huéspedes intermediarios, aumentando sus posibilidades de ser ingeridos por un depredador. Al igual que con los parásitos de transmisión directa, la distribución de parásitos de transmisión trófica entre individuos huéspedes es agregada. [22] La coinfección por múltiples parásitos es común. [27] La autoinfección , en la que (por excepción) todo el ciclo de vida del parásito tiene lugar en un único huésped primario, a veces puede ocurrir en helmintos como Strongyloides stercoralis . [28]

Los parásitos transmitidos por vectores dependen de un tercero, un huésped intermediario, donde el parásito no se reproduce sexualmente, [14] para llevarlos de un huésped definitivo a otro. [22] Estos parásitos son microorganismos, a saber, protozoos , bacterias o virus , a menudo patógenos intracelulares (causantes de enfermedades). [22] Sus vectores son en su mayoría artrópodos hematófagos como pulgas, piojos, garrapatas y mosquitos. [22] [29] Por ejemplo, la garrapata del ciervo Ixodes scapularis actúa como vector de enfermedades como la enfermedad de Lyme , la babesiosis y la anaplasmosis . [30] Los endoparásitos protozoarios, como los parásitos de la malaria del género Plasmodium y los parásitos de la enfermedad del sueño del género Trypanosoma , tienen estadios infecciosos en la sangre del huésped que son transportados a nuevos huéspedes por insectos que pican. [31]
Los parasitoides son insectos que tarde o temprano matan a sus hospedadores, lo que hace que su relación sea cercana a la depredación. [32] La mayoría de los parasitoides son avispas parasitoides u otros himenópteros ; otros incluyen dípteros como las moscas fóridas . Se pueden dividir en dos grupos, idiobiontes y koinobiontes, que difieren en su trato con sus hospedadores. [33]
Los parasitoides idiobiontes pican a sus presas, que suelen ser de gran tamaño, y las matan o las paralizan inmediatamente. La presa inmovilizada es luego llevada a un nido, a veces junto con otras presas si no es lo suficientemente grande como para sostener al parasitoide durante su desarrollo. Ponen un huevo sobre la presa y luego se sella el nido. El parasitoide se desarrolla rápidamente a través de sus etapas de larva y pupa, alimentándose de las provisiones que le dejan. [33]
Los parasitoides koinobiontes , que incluyen moscas y avispas, ponen sus huevos dentro de huéspedes jóvenes, generalmente larvas. A estas se les permite seguir creciendo, por lo que el huésped y el parasitoide se desarrollan juntos durante un período prolongado, que termina cuando los parasitoides emergen como adultos, dejando a la presa muerta, comida desde adentro. Algunos koinobiontes regulan el desarrollo de su huésped, por ejemplo, impidiendo que se transforme en pupa o haciendo que mude cuando el parasitoide esté listo para mudar. Pueden hacerlo produciendo hormonas que imitan las hormonas de muda del huésped ( ecdiesteroides ), o regulando el sistema endocrino del huésped. [33]


Un microdepredador ataca a más de un huésped, reduciendo la aptitud de cada huésped al menos en una pequeña cantidad, y solo está en contacto con cualquier huésped de manera intermitente. Este comportamiento hace que los microdepredadores sean adecuados como vectores, ya que pueden pasar parásitos más pequeños de un huésped a otro. [22] [34] [23] La mayoría de los microdepredadores son hematófagos y se alimentan de sangre. Incluyen anélidos como sanguijuelas , crustáceos como branquiuros e isópodos gnátidos , varios dípteros como mosquitos y moscas tsé-tsé , otros artrópodos como pulgas y garrapatas, vertebrados como lampreas y mamíferos como murciélagos vampiros . [22]

Los parásitos utilizan una variedad de métodos para infectar a los huéspedes animales, incluido el contacto físico, la ruta fecal-oral , etapas infecciosas de vida libre y vectores, que se adaptan a sus diferentes huéspedes, ciclos de vida y contextos ecológicos. [35] En la tabla se dan ejemplos para ilustrar algunas de las muchas combinaciones posibles.
Entre las muchas variaciones de las estrategias parasitarias se encuentran el hiperparasitismo, [37] el parasitismo social, [38] el parasitismo de cría, [39] el cleptoparasitismo, [40] el parasitismo sexual, [41] y el adelfoparasitismo. [42]
Los hiperparásitos se alimentan de otro parásito, como lo ejemplifican los protozoos que viven en parásitos helmintos, [37] o parasitoides facultativos u obligados cuyos huéspedes son parásitos convencionales o parasitoides. [22] [33] También se dan niveles de parasitismo más allá del secundario, especialmente entre los parasitoides facultativos. En los sistemas de agallas de roble , puede haber hasta cinco niveles de parasitismo. [43]
Los hiperparásitos pueden controlar las poblaciones de sus huéspedes y se utilizan para este propósito en la agricultura y, en cierta medida, en la medicina . Los efectos de control se pueden ver en la forma en que el virus CHV1 ayuda a controlar el daño que la plaga del castaño , Cryphonectria parasitica , causa a los castaños americanos , y en la forma en que los bacteriófagos pueden limitar las infecciones bacterianas. Es probable, aunque poco investigado, que la mayoría de los microparásitos patógenos tengan hiperparásitos que pueden resultar muy útiles tanto en la agricultura como en la medicina. [44]
Los parásitos sociales se aprovechan de las interacciones interespecíficas entre miembros de animales eusociales como las hormigas , las termitas y los abejorros . Algunos ejemplos son la gran mariposa azul, Phengaris arion , cuyas larvas emplean el mimetismo de las hormigas para parasitar a ciertas hormigas, [38] Bombus bohemicus , un abejorro que invade las colmenas de otras abejas y se hace cargo de la reproducción mientras sus crías son criadas por obreras anfitrionas, y Melipona scutellaris , una abeja eusocial cuyas reinas vírgenes escapan de las obreras asesinas e invaden otra colonia sin reina. [45] Un ejemplo extremo de parasitismo social interespecífico se encuentra en la hormiga Tetramorium inquilinum , un parásito obligado que vive exclusivamente en las espaldas de otras hormigas Tetramorium . [46] Un mecanismo para la evolución del parasitismo social fue propuesto por primera vez por Carlo Emery en 1909. [47] Ahora conocida como " regla de Emery ", establece que los parásitos sociales tienden a estar estrechamente relacionados con sus huéspedes, y a menudo pertenecen al mismo género. [48] [49] [50]
El parasitismo social intraespecífico se produce en la lactancia parasitaria, en la que algunas crías individuales toman leche de hembras no emparentadas. En los monos capuchinos , las hembras de rango superior a veces toman leche de hembras de rango inferior sin ninguna reciprocidad. [51]
En el parasitismo de cría , los hospedadores actúan como padres mientras crían a las crías como si fueran suyas. Los parásitos de cría incluyen aves de diferentes familias, como los tordos , los urogallos , los cucos y los patos de cabeza negra . Estos no construyen nidos propios, sino que dejan sus huevos en nidos de otras especies . Los huevos de algunos parásitos de cría imitan a los de sus hospedadores, mientras que algunos huevos de tordos tienen cáscaras duras, lo que hace que sea difícil para los hospedadores matarlos perforándolos, ambos mecanismos implican una selección por parte de los hospedadores contra los huevos parásitos. [39] [52] [53] La hembra adulta del cuco europeo imita aún más a un depredador, el gavilán europeo , lo que le da tiempo para poner sus huevos en el nido del hospedador sin ser observada. [54]
En el cleptoparasitismo (del griego κλέπτης ( kleptēs ), "ladrón"), los parásitos roban la comida recogida por el huésped. El parasitismo se da a menudo sobre parientes cercanos, ya sea dentro de la misma especie o entre especies del mismo género o familia. Por ejemplo, los numerosos linajes de abejas cucú ponen sus huevos en las celdas de nido de otras abejas de la misma familia. [40] El cleptoparasitismo es poco común en general, pero es evidente en las aves; algunas, como las págalos, están especializadas en piratear la comida de otras aves marinas, persiguiéndolas implacablemente hasta que regurgitan su captura. [55]
En algunas especies de rape , como Ceratias holboelli , se observa un enfoque único: los machos se reducen a pequeños parásitos sexuales , totalmente dependientes de las hembras de su propia especie para sobrevivir, permanentemente adheridos debajo del cuerpo de la hembra e incapaces de valerse por sí mismos. La hembra nutre al macho y lo protege de los depredadores, mientras que el macho no le devuelve nada excepto el esperma que la hembra necesita para producir la siguiente generación. [41]
El adelfoparasitismo (del griego ἀδελφός ( adelphós ), hermano [56] ), también conocido como parasitismo entre hermanos, ocurre cuando la especie hospedadora está estrechamente relacionada con el parásito, a menudo en la misma familia o género. [42] En el parasitoide de la mosca negra de los cítricos, Encarsia perplexa , las hembras no apareadas pueden poner huevos haploides en las larvas completamente desarrolladas de su propia especie, produciendo crías masculinas, [57] mientras que el gusano marino Bonellia viridis tiene una estrategia reproductiva similar, aunque las larvas son planctónicas. [58]
Se ilustran ejemplos de las principales estrategias variantes.
El parasitismo tiene un rango taxonómico extremadamente amplio, que incluye animales, plantas, hongos, protozoos, bacterias y virus. [59]
El parasitismo está muy extendido en el reino animal, [63] y ha evolucionado independientemente de las formas de vida libre cientos de veces. [22] Muchos tipos de helmintos , incluidos los trematodos y los cestodos, tienen ciclos de vida completos que involucran a dos o más huéspedes. El grupo más grande, con diferencia, es el de las avispas parasitoides de los himenópteros. [22] Los filos y clases con el mayor número de especies parásitas se enumeran en la tabla. Los números son estimaciones mínimas conservadoras. Las columnas de endoparasitismo y ectoparasitismo se refieren al huésped definitivo, como se documenta en las columnas de vertebrados e invertebrados. [60]

Un hemiparásito o parásito parcial como el muérdago obtiene algunos de sus nutrientes de otra planta viva, mientras que un holoparásito como la cuscuta obtiene todos sus nutrientes de otra planta. [64] Las plantas parásitas constituyen alrededor del uno por ciento de las angiospermas y se encuentran en casi todos los biomas del mundo. [65] [66] [67] Todas estas plantas tienen raíces modificadas, haustorios , que penetran en las plantas hospedantes, conectándolas al sistema conductor, ya sea el xilema , el floema o ambos. Esto les proporciona la capacidad de extraer agua y nutrientes del hospedante. Una planta parásita se clasifica dependiendo de dónde se adhiere al hospedante, ya sea el tallo o la raíz, y la cantidad de nutrientes que requiere. Dado que los holoparásitos no tienen clorofila y, por lo tanto, no pueden producir alimento por sí mismos mediante la fotosíntesis , siempre son parásitos obligados, que obtienen todo su alimento de sus hospedantes. [66] Algunas plantas parásitas pueden localizar a sus plantas hospedantes detectando sustancias químicas en el aire o el suelo emitidas por los brotes o raíces hospedantes , respectivamente. Se conocen alrededor de 4.500 especies de plantas parásitas en aproximadamente 20 familias de plantas con flores . [68] [66]
Las especies de la familia Orobanchaceae (nabos) se encuentran entre las plantas más destructivas económicamente. Se estima que las especies de Striga (hierba bruja) cuestan miles de millones de dólares al año en pérdidas de rendimiento de los cultivos, infestando más de 50 millones de hectáreas de tierras cultivadas solo en el África subsahariana. Striga infecta tanto pastos como cereales, incluidos el maíz , el arroz y el sorgo , que se encuentran entre los cultivos alimentarios más importantes del mundo. Orobanche también amenaza una amplia gama de otros cultivos importantes, incluidos los guisantes , los garbanzos , los tomates , las zanahorias y las variedades de repollo . La pérdida de rendimiento causada por Orobanche puede ser total; a pesar de una amplia investigación, ningún método de control ha sido completamente exitoso. [69]
Muchas plantas y hongos intercambian carbono y nutrientes en relaciones micorrízicas mutualistas . Sin embargo, unas 400 especies de plantas micoheterótrofas , principalmente en los trópicos, hacen trampa de manera efectiva al tomar carbono de un hongo en lugar de intercambiarlo por minerales. Tienen raíces muy reducidas, ya que no necesitan absorber agua del suelo; sus tallos son delgados con pocos haces vasculares , y sus hojas se reducen a pequeñas escamas, ya que no realizan la fotosíntesis. Sus semillas son muy pequeñas y numerosas, por lo que parecen depender de ser infectadas por un hongo adecuado poco después de germinar. [70]

Los hongos parásitos obtienen parte o la totalidad de sus requerimientos nutricionales de plantas, otros hongos o animales.
Los hongos fitopatógenos se clasifican en tres categorías según su modo de nutrición: biótrofos, hemibiótrofos y necrótrofos. Los hongos biótrofos obtienen nutrientes de las células vegetales vivas y, durante el curso de la infección, colonizan su planta huésped de tal manera que la mantienen viva durante un tiempo máximo. [71] Un ejemplo bien conocido de un patógeno biótrofo es Ustilago maydis , agente causante de la enfermedad del carbón del maíz. Los patógenos necrotróficos, por otro lado, matan las células huésped y se alimentan de forma saprofita , un ejemplo son los hongos melíferos colonizadores de raíces del género Armillaria . [72] Los patógenos hemibiótrofos comienzan a colonizar sus huéspedes como biótrofos y, posteriormente, matan las células huésped y se alimentan como necrótrofos, un fenómeno denominado cambio de biotrofia a necrotrofia. [73]
Los hongos patógenos son agentes causales bien conocidos de enfermedades en animales y humanos. Se estima que las infecciones fúngicas ( micosis ) matan a 1,6 millones de personas cada año. [74] Un ejemplo de un potente patógeno fúngico animal son los microsporidios , hongos parásitos intracelulares obligados que afectan principalmente a los insectos, pero también pueden afectar a los vertebrados, incluidos los humanos, causando la infección intestinal microsporidiosis . [75]

Los protozoos como Plasmodium , Trypanosoma y Entamoeba [76] son endoparásitos. Causan enfermedades graves en vertebrados, incluidos los humanos (en estos casos, malaria, enfermedad del sueño y disentería amebiana ) y tienen ciclos de vida complejos. [31]
Muchas bacterias son parásitas, aunque generalmente se las considera patógenos que causan enfermedades. [77] Las bacterias parásitas son extremadamente diversas e infectan a sus huéspedes por diversas vías. Por dar algunos ejemplos, Bacillus anthracis , la causa del ántrax , se propaga por contacto con animales domésticos infectados ; sus esporas , que pueden sobrevivir durante años fuera del cuerpo, pueden ingresar al huésped a través de una abrasión o pueden inhalarse. Borrelia , la causa de la enfermedad de Lyme y la fiebre recurrente , se transmite por vectores, garrapatas del género Ixodes , desde los reservorios de las enfermedades en animales como los ciervos . Campylobacter jejuni , una causa de gastroenteritis , se propaga por vía fecal-oral de los animales, o al comer aves de corral insuficientemente cocidas , o por agua contaminada. Haemophilus influenzae , un agente de meningitis bacteriana e infecciones del tracto respiratorio como la gripe y la bronquitis , se transmite por contacto con gotitas. Treponema pallidum , la causa de la sífilis , se transmite a través de la actividad sexual . [78]

Los virus son parásitos intracelulares obligados, caracterizados por una función biológica extremadamente limitada, hasta el punto de que, si bien evidentemente pueden infectar a todos los demás organismos, desde bacterias y arqueas hasta animales, plantas y hongos, no está claro si pueden describirse como vivos. Pueden ser virus de ARN o ADN que consisten en una cadena simple o doble de material genético ( ARN o ADN , respectivamente), cubierto por una capa de proteína y, a veces, una envoltura lipídica . Por lo tanto, carecen de toda la maquinaria habitual de la célula , como las enzimas , y dependen completamente de la capacidad de la célula huésped para replicar el ADN y sintetizar proteínas. La mayoría de los virus son bacteriófagos que infectan bacterias. [79] [80] [81] [82]

El parasitismo es un aspecto importante de la ecología evolutiva; por ejemplo, casi todos los animales que viven en libertad son huéspedes de al menos una especie de parásito. Los vertebrados, el grupo mejor estudiado, son huéspedes de entre 75.000 y 300.000 especies de helmintos y de un número incontable de microorganismos parásitos. En promedio, una especie de mamífero alberga cuatro especies de nematodos, dos de trematodos y dos de cestodos. [83] Los humanos tienen 342 especies de parásitos helmintos y 70 especies de parásitos protozoarios. [84] Aproximadamente tres cuartas partes de los eslabones de las redes alimentarias incluyen un parásito, importante para regular el número de huéspedes. Quizás el 40 por ciento de las especies descritas son parásitas. [83]
El parasitismo es difícil de demostrar a partir del registro fósil , pero los agujeros en las mandíbulas de varios especímenes de Tyrannosaurus pueden haber sido causados por parásitos similares a Trichomonas . [85] Saurophthirus , la pulga del Cretácico Inferior , parasitó a los pterosaurios . [86] [87] Se encontraron huevos que pertenecían a gusanos nematodos y probablemente quistes protozoarios en el coprolito del Triásico Tardío del fitosaurio . Este raro hallazgo en Tailandia revela más sobre la ecología de los parásitos prehistóricos. [88]
A medida que los hospedadores y los parásitos evolucionan juntos, sus relaciones suelen cambiar. Cuando un parásito tiene una relación exclusiva con un hospedador, la selección hace que la relación se vuelva más benigna, incluso mutualista, ya que el parásito puede reproducirse durante más tiempo si su hospedador vive más tiempo. [89] Pero cuando los parásitos compiten, la selección favorece al parásito que se reproduce más rápido, lo que conduce a una mayor virulencia. Por lo tanto, existen diversas posibilidades en la coevolución hospedador-parásito . [90]
La epidemiología evolutiva analiza cómo se propagan y evolucionan los parásitos, mientras que la medicina darwiniana aplica un pensamiento evolutivo similar a enfermedades no parasitarias como el cáncer y las enfermedades autoinmunes . [91]

Las asociaciones a largo plazo pueden dar lugar a una relación relativamente estable que tienda al comensalismo o al mutualismo , ya que, en igualdad de condiciones, es de interés evolutivo del parásito que su anfitrión prospere. Un parásito puede evolucionar para volverse menos dañino para su anfitrión o un anfitrión puede evolucionar para hacer frente a la presencia inevitable de un parásito, hasta el punto de que la ausencia del parásito le cause daño al anfitrión. Por ejemplo, aunque los animales parasitados por gusanos a menudo sufren daños evidentes, dichas infecciones también pueden reducir la prevalencia y los efectos de los trastornos autoinmunes en los huéspedes animales, incluidos los humanos. [89] En un ejemplo más extremo, algunos gusanos nematodos no pueden reproducirse, o incluso sobrevivir, sin la infección por la bacteria Wolbachia . [92]
Lynn Margulis y otros han argumentado, siguiendo el libro de Peter Kropotkin de 1902 Ayuda mutua: un factor de evolución , que la selección natural hace que las relaciones pasen del parasitismo al mutualismo cuando los recursos son limitados. Este proceso puede haber estado involucrado en la simbiogénesis que formó los eucariotas a partir de una relación intracelular entre arqueas y bacterias, aunque la secuencia de eventos sigue siendo en gran parte indefinida. [93] [94]
Se puede esperar que la competencia entre parásitos favorezca a los parásitos que se reproducen más rápido y, por lo tanto, son más virulentos , por selección natural . [90] [95]
.jpg/1280px-Pink_Flamingos_with_Duck_-_Camargue,_France_-_April_2007_(cropped).jpg)
Entre las bacterias parásitas que matan insectos de los géneros Photorhabdus y Xenorhabdus , la virulencia dependía de la potencia relativa de las toxinas antimicrobianas ( bacteriocinas ) producidas por las dos cepas involucradas. Cuando sólo una bacteria podía matar a la otra, la otra cepa era excluida por la competencia. Pero cuando las orugas eran infectadas con bacterias que tenían toxinas capaces de matar a la otra cepa, ninguna cepa era excluida, y su virulencia era menor que cuando el insecto era infectado por una sola cepa. [90]
Un parásito a veces experimenta coespeciación con su huésped, lo que da como resultado el patrón descrito en la regla de Fahrenholz , según el cual las filogenias del huésped y del parásito llegan a reflejarse entre sí. [96]
Un ejemplo es el virus espumoso de los simios (SFV) y sus hospedadores primates. Se descubrió que las filogenias de la polimerasa del SFV y la subunidad II de la citocromo c oxidasa mitocondrial de los primates africanos y asiáticos eran muy congruentes en cuanto al orden de ramificación y los tiempos de divergencia, lo que implica que los virus espumosos de los simios coespeciaron con los primates del Viejo Mundo durante al menos 30 millones de años. [97]
La presunción de una historia evolutiva compartida entre parásitos y hospedadores puede ayudar a dilucidar cómo se relacionan los taxones hospedadores. Por ejemplo, ha habido una disputa sobre si los flamencos están más estrechamente relacionados con las cigüeñas o los patos . El hecho de que los flamencos compartan parásitos con patos y gansos fue tomado inicialmente como evidencia de que estos grupos estaban más estrechamente relacionados entre sí que con las cigüeñas. Sin embargo, eventos evolutivos como la duplicación o la extinción de especies de parásitos (sin eventos similares en la filogenia del hospedador) a menudo erosionan las similitudes entre las filogenias del hospedador y el parásito. En el caso de los flamencos, tienen piojos similares a los de los somormujos . Los flamencos y los somormujos tienen un ancestro común, lo que implica coespeciación de aves y piojos en estos grupos. Los piojos de los flamencos luego cambiaron de hospedadores a patos, creando la situación que había confundido a los biólogos. [98]
.jpg/1280px-Toxoplasma_gondii_(2).jpg)
Los parásitos infectan a los huéspedes simpátricos (aquellos dentro de su misma área geográfica) de manera más efectiva, como se ha demostrado con los trematodos digenéticos que infectan a los caracoles de lago. [99] Esto está en línea con la hipótesis de la Reina Roja , que establece que las interacciones entre especies conducen a una selección natural constante para la coadaptación. Los parásitos rastrean los fenotipos de los huéspedes locales comunes, por lo que los parásitos son menos infecciosos para los huéspedes alopátricos , aquellos de diferentes regiones geográficas. [99]
Algunos parásitos modifican el comportamiento del huésped para aumentar su transmisión entre huéspedes, a menudo en relación con el depredador y la presa ( el parásito aumentó la transmisión trófica ). Por ejemplo, en la marisma costera de California , la duela Euhaplorchis californiensis reduce la capacidad de su huésped killifish para evitar a los depredadores. [100] Este parásito madura en garcetas , que tienen más probabilidades de alimentarse de killifish infectados que de peces no infectados. Otro ejemplo es el protozoo Toxoplasma gondii , un parásito que madura en gatos pero que puede ser transportado por muchos otros mamíferos . Las ratas no infectadas evitan los olores de gato, pero las ratas infectadas con T. gondii se sienten atraídas por este olor, lo que puede aumentar la transmisión a los huéspedes felinos. [101] El parásito de la malaria modifica el olor de la piel de sus huéspedes humanos, aumentando su atractivo para los mosquitos y, por lo tanto, mejorando la posibilidad de que el parásito se transmita. [36] La araña Cyclosa argenteoalba suele tener larvas de avispas parasitoides adheridas a ella, lo que altera su comportamiento de construcción de telarañas. En lugar de producir sus habituales telas pegajosas en forma de espiral, creaban telas simplificadas cuando se les adherían los parásitos. Este comportamiento manipulado duraba más y era más notorio cuanto más tiempo se dejaban los parásitos en las arañas. [102]

Los parásitos pueden explotar a sus hospedadores para llevar a cabo una serie de funciones que de otro modo tendrían que llevar a cabo por sí mismos. Los parásitos que pierden esas funciones tienen una ventaja selectiva, ya que pueden desviar recursos hacia la reproducción. Muchos insectos ectoparásitos, incluidas las chinches , las chinches murciélago , los piojos y las pulgas , han perdido su capacidad de volar y dependen en cambio de sus hospedadores para el transporte. [103] La pérdida de rasgos es más generalizada entre los parásitos. [104] Un ejemplo extremo es el mixospario Henneguya zschokkei , un ectoparásito de los peces y el único animal conocido que ha perdido la capacidad de respirar aeróbicamente: sus células carecen de mitocondrias . [105]
Los huéspedes han desarrollado una variedad de medidas defensivas contra sus parásitos, incluidas barreras físicas como la piel de los vertebrados, [106] el sistema inmunológico de los mamíferos, [107] insectos que eliminan activamente los parásitos, [108] y sustancias químicas defensivas en las plantas. [109]
El biólogo evolutivo WD Hamilton sugirió que la reproducción sexual podría haber evolucionado para ayudar a derrotar a múltiples parásitos al permitir la recombinación genética , la mezcla de genes para crear combinaciones variadas. Hamilton demostró mediante modelos matemáticos que la reproducción sexual sería evolutivamente estable en diferentes situaciones, y que las predicciones de la teoría coincidían con la ecología real de la reproducción sexual. [110] [111] Sin embargo, puede haber una compensación entre la inmunocompetencia y la reproducción de las características sexuales secundarias de los huéspedes vertebrados machos , como el plumaje de los pavos reales y las melenas de los leones . Esto se debe a que la hormona masculina testosterona fomenta el crecimiento de las características sexuales secundarias, favoreciendo a dichos machos en la selección sexual , al precio de reducir sus defensas inmunológicas. [112]
.jpg/1280px-Short_Horned_Lizard_(4457945238).jpg)
La barrera física de la piel dura y a menudo seca e impermeable de los reptiles, aves y mamíferos evita que los microorganismos invasores entren en el cuerpo. La piel humana también secreta sebo , que es tóxico para la mayoría de los microorganismos. [106] Por otro lado, los parásitos más grandes, como los trematodos, detectan las sustancias químicas producidas por la piel para localizar a sus huéspedes cuando entran en el agua. La saliva y las lágrimas de los vertebrados contienen lisozima , una enzima que descompone las paredes celulares de las bacterias invasoras. [106] Si el organismo pasa por la boca, el estómago con su ácido clorhídrico , tóxico para la mayoría de los microorganismos, es la siguiente línea de defensa. [106] Algunos parásitos intestinales tienen una capa exterior gruesa y dura que se digiere lentamente o no se digiere en absoluto, lo que permite que el parásito pase vivo por el estómago, momento en el que ingresa al intestino y comienza la siguiente etapa de su vida. Una vez dentro del cuerpo, los parásitos deben superar las proteínas séricas del sistema inmunológico y los receptores de reconocimiento de patrones , intracelulares y celulares, que activan los linfocitos del sistema inmunológico adaptativo, como las células T y las células B productoras de anticuerpos . Estas tienen receptores que reconocen a los parásitos. [107]

Los insectos a menudo adaptan sus nidos para reducir el parasitismo. Por ejemplo, una de las razones principales por las que la avispa Polistes canadensis anida en múltiples panales , en lugar de construir un solo panal como gran parte del resto de su género, es para evitar la infestación por polillas tineidas . La polilla tineida pone sus huevos dentro de los nidos de las avispas y luego estos huevos eclosionan en larvas que pueden excavar de celda en celda y cazar pupas de avispa. Las avispas adultas intentan eliminar y matar los huevos y larvas de polilla masticando los bordes de las celdas, recubriendo las celdas con una secreción oral que le da al nido una apariencia marrón oscura. [108]
Las plantas responden al ataque de parásitos con una serie de defensas químicas, como la polifenol oxidasa , bajo el control de las vías de señalización insensibles al ácido jasmónico (JA) y al ácido salicílico (SA). [109] [113] Las diferentes vías bioquímicas se activan por diferentes ataques, y las dos vías pueden interactuar positiva o negativamente. En general, las plantas pueden iniciar una respuesta específica o no específica. [114] [113] Las respuestas específicas implican el reconocimiento de un parásito por los receptores celulares de la planta, lo que lleva a una respuesta fuerte pero localizada: se producen químicos defensivos alrededor del área donde se detectó el parásito, bloqueando su propagación y evitando desperdiciar la producción defensiva donde no es necesaria. [114] Las respuestas defensivas no específicas son sistémicas, lo que significa que las respuestas no se limitan a un área de la planta, sino que se extienden por toda la planta, lo que las hace costosas en energía. Estas son efectivas contra una amplia gama de parásitos. [114] Cuando las hojas de plantas como el maíz y el algodón sufren daños, como por ejemplo a causa de orugas lepidópteros , liberan mayores cantidades de sustancias químicas volátiles, como terpenos , que indican que están siendo atacadas; uno de los efectos de esto es atraer avispas parasitoides, que a su vez atacan a las orugas. [115]

El parasitismo y la evolución de los parásitos fueron estudiados hasta el siglo XXI por parasitólogos , en una ciencia dominada por la medicina, en lugar de por ecólogos o biólogos evolutivos . Aunque las interacciones parásito-huésped eran claramente ecológicas e importantes en la evolución, la historia de la parasitología provocó lo que el ecólogo evolutivo Robert Poulin llamó una "toma de posesión del parasitismo por parte de los parasitólogos", lo que llevó a los ecólogos a ignorar el área. Esto fue en su opinión "desafortunado", ya que los parásitos son "agentes omnipresentes de la selección natural" y fuerzas significativas en la evolución y la ecología. [116] En su opinión, la división de larga data entre las ciencias limitó el intercambio de ideas, con conferencias separadas y revistas separadas. Los lenguajes técnicos de la ecología y la parasitología a veces implicaban diferentes significados para las mismas palabras. También hubo diferencias filosóficas: Poulin señala que, influenciados por la medicina, "muchos parasitólogos aceptaron que la evolución condujo a una disminución de la virulencia de los parásitos, mientras que la teoría evolutiva moderna habría predicho una mayor variedad de resultados". [116]
Sus complejas relaciones hacen que sea difícil ubicar a los parásitos en las redes alimentarias: un trematodo con múltiples hospedadores para sus diversas etapas del ciclo de vida ocuparía muchas posiciones en una red alimentaria simultáneamente y establecería bucles de flujo de energía, lo que confundiría el análisis. Además, dado que casi todos los animales tienen (múltiples) parásitos, estos ocuparían los niveles superiores de cada red alimentaria. [84]
Los parásitos pueden desempeñar un papel en la proliferación de especies no nativas. Por ejemplo, los cangrejos verdes invasores se ven mínimamente afectados por los trematodos nativos en la costa este del Atlántico. Esto los ayuda a competir con cangrejos nativos como los cangrejos de roca del Atlántico y los cangrejos Jonah . [117]
La parasitología ecológica puede ser importante para los intentos de control, como ocurrió durante la campaña para erradicar el gusano de Guinea . Aunque el parásito fue erradicado en todos los países excepto cuatro, el gusano comenzó a utilizar ranas como hospedador intermediario antes de infectar a los perros, lo que hizo que el control fuera más difícil de lo que hubiera sido si se hubieran entendido mejor las relaciones. [118]
Aunque se considera que los parásitos son dañinos, la erradicación de todos ellos no sería beneficiosa. Los parásitos representan al menos la mitad de la diversidad de la vida; desempeñan importantes funciones ecológicas; y sin parásitos, los organismos podrían tender a la reproducción asexual, disminuyendo la diversidad de rasgos que genera la reproducción sexual. [119] Los parásitos brindan una oportunidad para la transferencia de material genético entre especies, lo que facilita el cambio evolutivo. [120] Muchos parásitos requieren múltiples hospedadores de diferentes especies para completar sus ciclos de vida y dependen de interacciones depredador-presa u otras interacciones ecológicas estables para llegar de un hospedador a otro. La presencia de parásitos, por lo tanto, indica que un ecosistema es saludable. [121]
Un ectoparásito, el piojo del cóndor de California, Colpocephalum californici , se convirtió en un conocido problema de conservación. En Estados Unidos se llevó a cabo un importante y muy costoso programa de cría en cautiverio para rescatar al cóndor de California . Era el huésped de un piojo que vivía únicamente sobre él. Todos los piojos encontrados fueron "deliberadamente asesinados" durante el programa, para mantener a los cóndores en el mejor estado de salud posible. El resultado fue que una especie, el cóndor, fue salvada y devuelta a la naturaleza, mientras que otra especie, el parásito, se extinguió. [122]
Aunque los parásitos suelen omitirse en las representaciones de las redes alimentarias , suelen ocupar la primera posición. Los parásitos pueden funcionar como especies clave , reduciendo el predominio de competidores superiores y permitiendo que las especies competidoras coexistan. [84] [123] [124]

Una sola especie de parásito suele tener una distribución agregada entre los animales hospedadores, lo que significa que la mayoría de los hospedadores son portadores de pocos parásitos, mientras que unos pocos hospedadores son portadores de la gran mayoría de individuos parásitos. Esto plantea problemas considerables para los estudiantes de ecología de parásitos, ya que invalida las estadísticas paramétricas que suelen utilizar los biólogos. Varios autores recomiendan la transformación logarítmica de los datos antes de aplicar la prueba paramétrica o el uso de estadísticas no paramétricas , pero esto puede dar lugar a más problemas, por lo que la parasitología cuantitativa se basa en métodos bioestadísticos más avanzados. [125]
Los parásitos humanos , incluidos los gusanos redondos, el gusano de Guinea , los oxiuros y las tenias, se mencionan en registros de papiros egipcios desde el año 3000 a. C. en adelante; el papiro de Ebers describe el anquilostoma . En la antigua Grecia , los parásitos, incluido el gusano de la vejiga, se describen en el Corpus hipocrático , mientras que el dramaturgo cómico Aristófanes llamó a las tenias "piedras de granizo". Los médicos romanos Celso y Galeno documentaron los gusanos redondos Ascaris lumbricoides y Enterobius vermicularis . [126]

En su Canon de Medicina , completado en 1025, el médico persa Avicena registró parásitos humanos y animales, incluyendo lombrices intestinales, lombrices intestinales, gusanos de Guinea y tenias. [126]
En su libro de 1397 Traité de l'état, science et pratique de l'art de la Bergerie (Tratado del estado, la ciencia y la práctica del arte del pastoreo), Jehan de Brie escribió la primera descripción de un endoparásito trematodo, la duela del hígado de oveja Fasciola hepatica . [127] [128]
En el período moderno temprano , el libro de Francesco Redi de 1668 Esperienze Intorno alla Generazione degl'Insetti ( Experiencias de la generación de insectos ), describió explícitamente los ectoparásitos y endoparásitos, ilustrando las garrapatas , las larvas de las moscas nasales de los ciervos y la duela del hígado de las ovejas . [129] Redi señaló que los parásitos se desarrollan a partir de huevos, contradiciendo la teoría de la generación espontánea . [130] En su libro de 1684 Osservazioni intorno agli animali viventi che si trovano negli animali viventi ( Observaciones sobre animales vivos encontrados en animales vivos ), Redi describió e ilustró más de 100 parásitos, incluido el gran gusano redondo en humanos que causa ascariasis . [129] Redi fue el primero en nombrar los quistes de Echinococcus granulosus observados en perros y ovejas como parásitos; Un siglo después, en 1760, Peter Simon Pallas sugirió correctamente que se trataba de larvas de tenias. [126]
En 1681, Antonie van Leeuwenhoek observó e ilustró el parásito protozoario Giardia lamblia y lo relacionó con "sus propias heces blandas". Este fue el primer parásito protozoario de humanos que se vio bajo un microscopio. [126] Unos años más tarde, en 1687, los biólogos italianos Giovanni Cosimo Bonomo y Diacinto Cestoni describieron la sarna como causada por el ácaro parásito Sarcoptes scabiei , señalándola como la primera enfermedad de humanos con un agente causal microscópico conocido. [131]

La parasitología moderna se desarrolló en el siglo XIX con observaciones y experimentos precisos por parte de muchos investigadores y médicos; [127] el término se utilizó por primera vez en 1870. [132] En 1828, James Annersley describió la amebiasis , infecciones protozoarias de los intestinos y el hígado, aunque el patógeno, Entamoeba histolytica , no fue descubierto hasta 1873 por Friedrich Lösch. James Paget descubrió el nematodo intestinal Trichinella spiralis en humanos en 1835. James McConnell describió el trematodo hepático humano, Clonorchis sinensis , en 1875. [126] Algernon Thomas y Rudolf Leuckart hicieron de forma independiente el primer descubrimiento del ciclo de vida de un trematodo, el trematodo hepático ovino, mediante un experimento en 1881-1883. [127] En 1877 Patrick Manson descubrió el ciclo de vida de los gusanos filariales que causan la elefantiasis transmitida por mosquitos. Manson predijo además que el parásito de la malaria , Plasmodium , tenía un mosquito vector, y persuadió a Ronald Ross para que investigara. Ross confirmó que la predicción era correcta en 1897-1898. Al mismo tiempo, Giovanni Battista Grassi y otros describieron las etapas del ciclo de vida del parásito de la malaria en los mosquitos Anopheles . Ross recibió el premio Nobel de 1902 por su trabajo, mientras que Grassi no lo recibió. [126] En 1903, David Bruce identificó el parásito protozoario y el vector de la mosca tsé-tsé de la tripanosomiasis africana . [133]
Dada la importancia de la malaria, con unos 220 millones de personas infectadas anualmente, se han hecho muchos intentos para interrumpir su transmisión. Se han probado varios métodos de profilaxis de la malaria , incluido el uso de medicamentos antipalúdicos para matar los parásitos en la sangre, la erradicación de sus mosquitos vectores con organoclorados y otros insecticidas , y el desarrollo de una vacuna contra la malaria . Todos estos han demostrado ser problemáticos, con resistencia a los medicamentos , resistencia a los insecticidas entre los mosquitos y fracaso repetido de las vacunas a medida que el parásito muta. [134] La primera y hasta 2015 la única vacuna autorizada para cualquier enfermedad parasitaria de los humanos es la RTS,S para la malaria por Plasmodium falciparum . [135]

Varios grupos de parásitos, incluidos patógenos microbianos y avispas parasitoides, se han utilizado como agentes de control biológico en la agricultura y la horticultura . [137] [138]
Poulin observa que el uso profiláctico generalizado de fármacos antihelmínticos en ovejas y ganado doméstico constituye un experimento mundial no controlado en la evolución del ciclo de vida de sus parásitos. Los resultados dependen de si los fármacos reducen la probabilidad de que una larva de helminto alcance la edad adulta. Si es así, se puede esperar que la selección natural favorezca la producción de huevos a una edad más temprana. Si, por otro lado, los fármacos afectan principalmente a los gusanos parásitos adultos , la selección podría causar una madurez tardía y un aumento de la virulencia . Tales cambios parecen estar en marcha: el nematodo Teladorsagia circumcincta está cambiando su tamaño adulto y su tasa reproductiva en respuesta a los fármacos. [139]

En la época clásica , el concepto de parásito no era estrictamente peyorativo: el parasitus era un rol aceptado en la sociedad romana , en el que una persona podía vivir de la hospitalidad de otros, a cambio de "adulación, servicios sencillos y una voluntad de soportar la humillación". [140] [141]
El parasitismo tiene un sentido peyorativo en el uso popular. Según el inmunólogo John Playfair, [142]
En el lenguaje cotidiano, el término «parásito» está cargado de significados peyorativos. Un parásito es un parásito, un aprovechador holgazán, una carga para la sociedad. [142]
El clérigo satírico Jonathan Swift alude al hiperparasitismo en su poema de 1733 "Sobre la poesía: una rapsodia", comparando a los poetas con "alimañas" que "molestan y pellizcan a sus enemigos": [143]
Las alimañas sólo molestan y pellizcan
a sus enemigos superiores por una pulgada.
Así observan los naturalistas: una pulga
tiene pulgas más pequeñas que la atacan;
y estas tienen pulgas más pequeñas que las pican.
Y así continúa hasta el infinito .
Así, cada poeta, en su especie,
es mordido por el que viene detrás:
Un estudio de 2022 examinó los nombres de unas 3000 especies de parásitos descubiertas en las dos décadas anteriores. De las que llevaban el nombre de científicos, más del 80 % llevaban el nombre de hombres, mientras que aproximadamente un tercio de los autores de artículos sobre parásitos eran mujeres. El estudio descubrió que el porcentaje de especies de parásitos que llevaban el nombre de familiares o amigos del autor había aumentado considerablemente en el mismo período. [144]

En la novela de terror gótico de Bram Stoker de 1897, Drácula , y sus numerosas adaptaciones cinematográficas , el epónimo Conde Drácula es un parásito bebedor de sangre (un vampiro). La crítica Laura Otis sostiene que, como "ladrón, seductor, creador e imitador, Drácula es el parásito definitivo. El objetivo del vampirismo es chupar la sangre de otras personas, vivir a expensas de otras personas". [145]
Las especies alienígenas parásitas repugnantes y aterradoras están muy extendidas en la ciencia ficción , [146] [147] como por ejemplo en la película Alien de Ridley Scott de 1979. [ 148] [149] En una escena, un xenomorfo sale del pecho de un hombre muerto, con sangre saliendo a chorros bajo alta presión asistida por petardos explosivos . Se utilizaron órganos animales para reforzar el efecto de choque. La escena se filmó en una sola toma, y la reacción de sorpresa de los actores fue genuina. [4] [150]
para asegurar su propia supervivencia y reproducción, pero al mismo tiempo con el mínimo dolor y costo para el huésped.
se reconocen mutuamente, en última instancia a nivel molecular, y los tejidos del huésped se estimulan para que reaccionen de alguna manera. Esto explica por qué el parasitismo puede provocar enfermedades, pero no siempre.
La depredación, la herbivoría y el parasitismo existen a lo largo de un continuo de gravedad en términos del grado en que afectan negativamente la aptitud de un organismo. ... En la mayoría de las situaciones, los parásitos no matan a sus huéspedes. Sin embargo, una excepción ocurre con los parasitoides, que difuminan la línea entre parasitismo y depredación.
{{cite journal}}: CS1 maint: varios nombres: lista de autores ( enlace )Se podría pensar que el siglo XIX es el momento de la génesis de la parasitología moderna.
su comida de su patrón, y para esto la personalidad prefabricada del parásito resultó extremadamente útil.
![Las avispas parasitoides idiobiontes paralizan inmediatamente a sus huéspedes para que sus larvas (Pimplinae, en la foto) se las coman.[22]](http://upload.wikimedia.org/wikipedia/commons/thumb/6/65/Live_Tetragnatha_montana_(RMNH.ARA.14127)_parasitized_by_Acrodactyla_quadrisculpta_larva_(RMNH.INS.593867)_-_BDJ.1.e992.jpg/1280px-Live_Tetragnatha_montana_(RMNH.ARA.14127)_parasitized_by_Acrodactyla_quadrisculpta_larva_(RMNH.INS.593867)_-_BDJ.1.e992.jpg)




.jpg/1280px-Great_Skua_(cropped).jpg)
.jpg/1280px-%D0%A1%D0%B5%D0%B2%D0%B5%D1%80%D0%BD%D0%B0%D1%8F_%D1%86%D0%B5%D1%80%D0%B0%D0%BF%D0%B8%D1%8F_(cropped).jpg)
