Transportador ABC
Algunas proteínas ABC no participan en el transporte de moléculas, sino en la traducción y reparación del ADN.
En los seres humanos se han encontrado 48 genes y mutaciones en ellos causan o contribuyen a diversas enfermedades,[5] como la fibrosis quística, la adrenoleucodistrofia, la enfermedad de Stargardt, los tumores resistentes a la quimioterapia, el síndrome de Dubin-Johnson, la colestasis intrahepática familiar progresiva, la anemia sideroblástica, la ataxia, y la hipoglucemia hiperinsuliménica.
Entre los sustratos incorporados a la célula por estos transportadores se cuentan iones, aminoácidos, péptidos, azúcares y otras moléculas hidrofílicas.
La porción transmembranal del transportador ABC aísla los sustratos de los lípidos presentes en la membrana mientras la atraviesan.
Otro ejemplo es producto del gen chvE-gguAB en Agrobacterium tumefaciens, un transportador de glucosa y galactosa igualmente asociado con la virulencia.
[12] Las proteínas ABC se han detectado en la placenta, donde podrían proteger al feto contra sustancias xenobióticas.
Puede transportar colesterol, análogos de fosfatidilcolina (PC), fosfatidiletanolamina (PE), fosfatidilserina (PS), esfingomielina (SM), y glucosilceramida (GlcCer).
Los dominos transmembrana presentan una gran variedad en su estructura y secuencias, consistente con la diversidad de sustratos que pueden transportar.
La secuencia de los dominios ABC, en cambio, está muy bien conservada, con muchas similitudes entre transportadores diferentes.
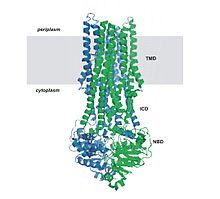
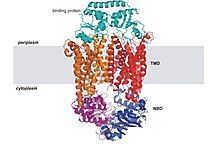
En la mayoría de los ABC secretores o exportadores, los cuatro subdominios forman una sola cadena polipeptídica con una disposición TM-NB-TM-NB.
El subdominio menor contiene tres o cuatro hélices y el llamado motivo característico ABC, motivo LSGGQ, péptido de enlace o motivo C. El dominio NB también tiene una glutamina en un bucle flexible conocido como bucle Q, tapadera o conmutador de fosfato-gamma, que conecta con el dominio transmembrana; se supone que el bucle Q participa en la interacción entre los dominios NB y TM y tiene que ver con la relación entre la hidrólisis del nucleótido y los cambios en conformación que experimenta el dominio transmembrana durante la traslocación del sustrato.
Una última zona, conocida como el motivo H or región de conmutación contiene una histidina muy conservada que desempeña un papel importante en la interacción del dominio NB con el ATP.
La denominación del dominio asociado al nucleótido como ABC o ATP-binding cassette proviene de la forma de caja o casete que adoptan los motivos que lo integran cuando se produce la hidrólisis del ATP.
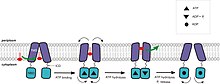
[3][10][18] La dimerización del dominio ABC o NB de los transportadores requiere la asociación con ATP.
[18] Se han elucidado varias estructuras del dominio ABC de varior importadores, como el HisP,[23] GlcV,[24] MJ1267,[25] E. coli MalK,[26] T. litoralis MalK,[27] y de secretores como TAP,[28] HlyB,[29] MJ0796,[30][31] Sav1866,[32] y MsbA.
[18] Esta conformación es consistente con resultados de experimentos bioquímicos que demuestran que el ATP se encuentra próximo a los aminoácidos del bucle P y el motivo LSGGQ durante el proceso de catálisis.
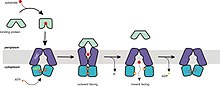
Al haber en esta posición menor afinidad entre el transportador y el sustrato, este queda libre.
No hay acuerdo sobre qué parte del ciclo proporciona la energía inicial para la función de los transportadores ABC.
Tras la hidrólisis del ATP, el dímero ABC se abre y el sustrato pasa al citoplasma.
[3][10][18][39][41] Los secretores o exportadores ABC procariotas abundan mucho y cuentan con homólogos cercanos en los organismos eucariotas.
[14] En las bacterias gram-negativas, los transportadores ABC secretan los sustratos a través de las membranas interior y exterior sin contacto con el periplasma.
[10] En el estado de reposo, los secretores adoptan la orientación hacia dentro, y los dominios transmembrana (TM) y de unión al nucleótido (ABC) se encuentran a una distancia relativamente amplia para poder acomodar sustratos anfifílicos o hidrófobos.
[44] No está claro exactamente cómo estas proteína pueden trasladar una variedad tan amplia de fármacos.
[4] En el ser humano, la subfamilia ABCA se compone de doce transportadores divididos a su vez en dos subgrupos.
Las proteínas MRP se encuentran en todos los seres vivos y median muchas funciones importantes.
La ABCD1 está asociada a una forma de la adrenoleucodistrofia (ALD) ligada al cromosoma X , enfermedad caracterizada por la neurodegeneración y la deficiencia suprarrenal que generalmente se inicia en la infancia tardía.
De la subfamilia ABCE solo se conoce un miembro, OABP or ABCE1, que reconoce algunos oligodendrocitos producidos en respuesta a ciertas infecciones virales.
[5] La subfamilia ABCG está compuesta por seis transportadores con sitios de unión al ATP en el extremo aminoterminal y dominios transmembrana en el extremo carboxilo, al contrario que las demás subfamilias de transportadores ABC.
ABCG2 es capaz de exportar antraciclinas usadas contra el cáncer, así como topotecán, mitoxantrona, o doxorubicina.