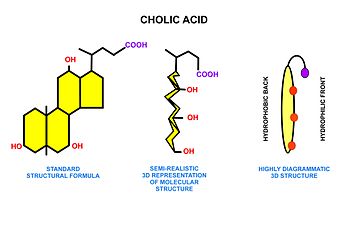
Metabolismo de los ácidos grasos
Las prostaglandinas, formadas a partir del ácido araquidónico almacenado en la membrana celular, probablemente sean el grupo mejor conocido de estas hormonas locales.
[10][11] Este es el destino del acetil-CoA donde sea que la β-oxidación de los ácidos grasos ocurra, excepto bajo ciertas circunstancias en el hígado.
[7] El acetoacetato, el beta-hidroxibutirato y su producto de degradación espontáneo, la acetona, se conocen frecuentemente (aunque un tanto equivocadamente) como cuerpos cetónicos (ya que no son "cuerpos" para nada, sino sustancias químicas solubles en agua).
[7] La situación donde aparecen niveles elevados de cuerpos cetónicos en la sangre durante el ayuno prolongado, dieta baja en carbohidratos, ejercicio pesado prolongado y diabetes tipo 1 descontrolada, se conoce como cetosis, y, en su forma extrema, como aparece en la diabetes tipo 1 sin control, se denomina cetoacidosis.
Las dos vías son diferentes, no solo en el lugar donde ocurren, sino en las reacciones implicadas, y en los sustratos empleados.
[1] Debido a esto no puede haber una conversión neta de ácidos grasos hacia glucosa.
El glicerol liberado a la circulación sanguínea durante la lipólisis de los triglicéridos en el tejido adiposos solamente puede ser tomado por el hígado.
Estos fosfolípidos pueden ser escindidos en diacilglicerol (DAG) e inositol trifosfato (IP3) a través de la hidrólisis de los fosfolípidos, y fosfatidilinositol 4,5-bisfosfato (PIP2), por la enzima fosfolipasa C (PLC) que se encuentra unida a la membrana.
Aunque el inositol trifosfato (IP3), difunde hacia el citosol, el diacilglicerol (DAG) permanece en el interior de la membrana plasmática debido a sus propiedades hidrofóbicas.
Se han encontrado prostaglandinas en casi todos los tejidos humanos y de otros animales.
Originalmente se pensaba que las prostaglandinas abandonaban la célula por difusión pasiva, debido a su alta lipofilicidad.
La capacidad de una determinada prostaglandina para estimular una reacción en un tejido e inhibir la misma reacción en un tejido diferente se encuentra determinada por el tipo de receptor al cual se une la prostaglandina.
Las prostaglandinas actúan como factores autócrinos o parácrinos estando sus células diana presentes en la inmediata vecindad del sitio de secreción.
Las prostaciclinas son poderosos vasodilatadores de acción local, e inhiben la agregación plaquetaria.
[16][17] Sino que son degradados a mono y diglicéridos más ácidos grasos líbres (aunque no glicerol libre) por la lipasa pancreática, la cual forma un complejo 1:1 con una proteína llamada colipasa (también constituyente del jugo pancreático), la cual es necesaria para su actividad.
[21] Los quilomicrones circulan a través del cuerpo, otorgándole al plasma sanguíneo un aspecto lechoso, o cremoso luego de una comida rica en grasas.
Los ácidos grasos son absorbidos por los adipocitos, pero el glicerol y quilomicrones remanentes permanecen en el plasma sanguíneo, siendo al final sacados de la circulación por el hígado.
Estos ácidos grasos se combinan con glicerol para formar triglicéridos que son empacados en gotitas muy similares a los quilomicrones, pero conocidas como lipoproteínas de muy baja densidad (VLDL por sus siglas en inglés).
[22] El tejido adiposo y la glándula mamaria lactante, también toman glucosa de la sangre para su conversión en triglicéridos.
[24] Estas reacciones son llevadas a cabo por la ácido graso sintasa II (FASII), la cual por lo general contiene múltiples enzimas que actúan como un complejo.
La FASII se encuentra presente en procariotas, plantas, hongos, y parásitos, como así también en las mitocondrias.
[26] En los animales, como así también en algunos hongos tales como la levadura, estas mismas reacciones ocurren en el complejo ácido graso sintasa I (FASI), una enorme proteína dimérica que posee todas las actividades enzimáticas necesarias para producir un ácido graso.
La elongación comenzando con el estearato (18:0), ocurre principalmente en el retículo endoplasmático por varias enzimas unidas a membrana.
Los pasos enzimáticos involucrados en este proceso de elongación son básicamente los mismos que los que lleva a cabo la FAS, pero los cuatro pasos sucesivos principales de la elongación son llevados a cabo por proteínas individuales, las cuales se pueden encontrar asociadas físicamente.
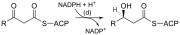
[27][28] Abreviaciones: ACP – Proteína transportadora de acilos, CoA – Coenzima A, NADP – Nicotinamida adenina dinucleótido fosfato.
[29] Sin embargo, sigue siendo desconocido como hacen los ácidos grasos esenciales, que los mamíferos no pueden sintetizar por sí mismos; para alcanzar a estas células, ya que son componentes muy importantes de estas membranas celulares, y necesarios para la síntesis de hormonas parácrinas (como se describe un poco más arriba).
El piruvato producido por la glucólisis es un intermediario importante en la conversión de carbohidratos a ácidos grasos y colesterol.
Sin embargo, este acetil-CoA necesita ser transportado al citosol donde ocurre la síntesis de ácidos grasos y colesterol.
Para obtener acetil-CoA, se quita citrato (producido por la condensación de acetil-CoA con oxaoloacetato) del ciclo del ácido cítrico, se lo transporta a través de la membrana mitocondrial interna hacia el citosol.
El control alostérico ocurre por medio de la retroalimentación ya sea inhibiéndose por el palmitoil-CoA o activándose por el citrato.





55 H
98 O
6


2 para formar IP
3 y DAG. El IP
3 inicia la liberación de calcio intracelular, mientras que el DAG activa una PKC ( proteína quinasa C ). Nota: La PLC ( fosfolipasa C ) no es un intermediario, como posiblemente podría sugerir este diagrama, sino que es la enzima que cataliza la separación de IP
3 /DAG. [ 13 ]