[1] Fue propuesta por la matemática inglesa Dorothy Wrinch a finales de los años 1930 basándose en tres premisas.Como clarificó Linderstrøm-Lang,[14] estos datos experimentales mostraban que las proteínas desnaturalizadas eran polipéptidos, pero no aportaban información acerca de la estructura o el plegamiento de estas macromoléculas, pues la desnaturalización podía provocar una transformación química que convirtiese en polipéptidos sencillos a las proteínas.[22] En una conferencia de Astbury en 1933 a la Oxford Junior Scientific Society, el físico Frederick Frank sugirió que la alfa-queratina, una escleroproteína, podría estabilizarse gracias a un mecanismo alternativo, la interacción covalente entre los enlaces peptídicos según la reacción del ciclol.[36] Dorothy Maud Wrinch propuso un modelo completo para explicar la estructura de las proteínas basado en la hipótesis del ciclol.Su éxito radica en proponer un modelo bien definido y examinable, de tal manera que se podía utilizar para predecir resultados experimentales.Sin embargo, los experimentos realizados en los años siguientes demostraron que la hipótesis del ciclol no era válida para las proteínas globulares.La acetamida resultante es descrita a la vez por las dos formas resonantes, colaborando la primera en un 62 % y la segunda en un 28 %.[45][46] Los primeros estudios mostraron que las proteínas desnaturalizadas por presión se encontraban frecuentemente en un estado diferente a las mismas proteínas desnaturalizadas por alta temperatura, lo que fue interpretado como una confirmación del modelo del ciclol.Este fue el tema del cuarto artículo de Wrinch sobre el modelo del ciclol (1936),[51] escrito conjuntamente con Dorothy Jordan Lloyd, que propuso en un principio que las moléculas globulares se estabilizaban por enlaces de hidrógeno.[52] Wrinch también escribió un artículo con William Astbury, presentando la posibilidad de que la tautomería ceto-enólica del >CαHα y el carbonilo de un grupo amida >C=O, formaran un enlace mixto >Cα-C(OHα)< dejando libre un oxígeno para el grupo hidroxilo.Más generalmente, el modelo del ciclol para proteínas globulares era consistente con los datos experimentales obtenidos por ultracentrifugación por Theodor Svedberg, lo que sugería que el peso molecular de las proteínas podía asimilarse a unas pocas clases de números enteros.[2][3] El modelo del ciclol se mostró consistente con las propiedades características que presentan las proteínas plegadas.En tercer lugar, algunas proteínas, como la hemoglobina, pueden formar enlaces organometálicos; como los metales de transición presentan geometrías de enlace específicas y regulares (por ejemplo octaédricas), se veía razonable que la proteína globalmente tuviera una estructura geométrica similar.En cuarto lugar, la hipótesis del ciclol proporciona una explicación químicamente simple para la desnaturalización y la dificultad de romper proteínas plegadas mediante proteasas.Wrinch finalmente resumió su modelo y los datos experimentales que parecían confirmarlo en tres artículos.[57][58][59] Tras haber propuesto un modelo verosímil para el plegamiento de las proteínas globulares, Wrinch investigó si era consistente con los datos estructurales disponibles en aquel tiempo.[80] Bergmann y Niemann proporcionaron más argumentos químicos en contra de la hipótesis del ciclol,[81] como hizo también Neuberger.[82][83] Los datos obtenidos mediante espectroscopia de infrarrojo mostraban que el número de grupos carbonilo presentes en una proteína no cambiaban tras una hidrólisis,[84] y que las proteínas plegadas e intactas contenían grupos amida-carbonilo complementarios;[85] ambas observaciones contradicen la hipótesis del ciclol de que los carbonilos se transforman en hidroxilos en las proteínas plegadas.Respecto a los impedimentos estéricos por excesiva cercanía de los átomos, propuso que pequeñas deformaciones en los ángulos y longitudes de enlace podían aliviar los impedimentos o al menos reducirlos a un nivel razonable.[113][114][115][116][117] También publicó dos libros en los que describió de nuevo la hipótesis del ciclol y los pequeños péptidos en general.En este caso la hipótesis central era que la forma de ciclol del grupo peptídico estaba favorecida frente a la amida.El estudio experimental debía demostrar si los grupos carbonilo de la proteína se transformaban masivamente en hidroxilos.[96] El modelo del ciclol también predecía una alta densidad lateral de aminoácidos en las proteínas plegadas y en película, que no se observó experimentalmente.[38] Sin embargo, la opinión general es que, si bien estos defectos la hacían poco verosímil, no la convertían en imposible.Curiosamente, el modelo del ciclol es un ejemplo de cómo una teoría científica de gran simetría y belleza, dos cualidades que suelen caracterizar a las teorías científicas «obviamente ciertas», se revela errónea.Por ejemplo, la doble hélice de Watson y Crick como modelo para el ADN[127] es un modelo «obvio» por sus enlaces de hidrógeno y simetría, mientras que otras estructuras mucho menos simétricas están favorecidas en condiciones diferentes.[128] También la teoría de la relatividad general fue considerada una hipótesis tan «elegante» que para Albert Einstein no parecía necesitar verificación experimental, aunque tuvo que revisarse para hacerla consistente con la teoría cuántica de campos.[129] El ejemplo de la hipótesis del ciclol ilustra cómo todas las teorías científicas, incluso las más elegantes, deben ser verificadas empíricamente y nunca ser consideradas ciertas a priori.
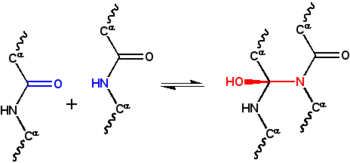
Reacción del ciclol clásica. Dos grupos peptídicos se unen por enlace N-C', convirtiendo el oxígeno carbonílico en una grupo hidroxilo. Si bien esta reacción ocurre en unos pocos péptidos cíclicos, es desfavorable por la
energía libre de Gibbs
, principalmente porque elimina la
estabilización por resonancia
del
enlace peptídico
. Esta reacción es la base de la
hipótesis
del ciclol.
Tránsito de la estructura primaria a la estructura terciaria de una proteína según el modelo de cadena polipeptídica.
La estructura de ciclol propuesta por
Dorothy Wrinch
para una proteína compuesta por seis moléculas de
alanina
. Se trata de una hexapéptido en la que tres
grupos peptídicos
están enlazados al anillo central por reacciones del ciclol. Los tres grupos externos, no enlazados, no son planos, sino que forman un
ángulo diedro
ω=60°. Los tres átomos de color rojo del anillo central representan los
grupos hidroxilo
formados por la reacción del ciclol, mientras que los exteriores corresponden a los
oxígenos
de los
carbonilos
. Los átomos de oxígeno interiores están separados por solo 2.45
Å
, una distancia extremadamente corta incluso para átomos unidos por
enlace de hidrógeno
. Esta molécula hipotética no ha sido observada en la naturaleza.
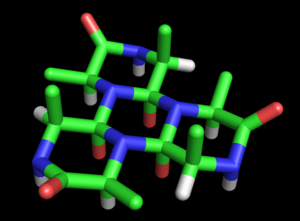
Modelo de varillas de la estructura de cicloles de
alanina
propuesta por
Dorothy Wrinch
. Conceptualmente es similar a una
lámina beta
, pero más uniforme y
densa
. La estructura presenta huecos en su patrón hexagonal, en el que tres átomos
C
β
(en verde) y tres átomos
H
α
(en blanco) convergen en un espacio relativamente vacío. Las dos caras de la estructura no son equivalentes: todos los C
β
emergen del mismo lado, situado en la parte superior de la figura. Los átomos en rojo representan
hidroxilos
(no
carbonilos
) y emergen, en grupos de tres, de las dos caras de la estructura. Finalmente, los átomos azules representan
nitrógeno
. Esta molécula hipotética no ha sido observada en la naturaleza.
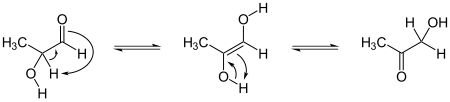
Mecanismo general de la tautomería ceto-enólica.
Modelo de varillas de la
estructura proteica
tipo ciclol C
1
propuesta por
Dorothy Wrinch
. Globalmente, la
macromolécula
es un
tetraedro
truncado compuesto por cuatro subestructuras de ciclol planas, cada una de ellas rodeando un espacio vacío y unidas entre sí par a par por cuatro residuos en cada lado (dos en cada vértice). Así, la molécula está formada por 72
aminoácidos
enlazados, en este caso de
alanina
. La estructura se asemeja a una
jaula
, permitiendo la entrada de molécula menores dentro de su estructura. Esta molécula hipotética no ha sido observada en la naturaleza.
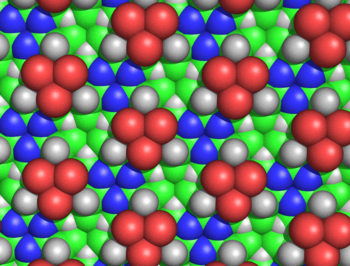
Diagrama de relleno del espacio de la estructura de cicloles de
alanina
, vista desde la cara sin
C
β
emergentes. Se puede observar la simetría de la estructura y su extraordinaria densidad. Por ejemplo, en el espacio libre en el que convergen tres átomos C
β
(en verde) y tres
H
α
(representados como triángulos blancos), los carbonos e hidrógenos están separados por solo 1.68
Å
, una longitud menor del
radio de van der Waals
medio del carbono, de 1.70 Å. Las esferas verdes representan los átomos C
β
. Los C
α
no son visibles desde esta perspectiva, excepto los pequeños triángulos cercanos a los
nitrógenos
(representados en azul). Los átomos rojos representan
hidroxilos
, no
carbonilos
.
Un
azaciclol
típico (rojo) en rápido
equilibrio
con su forma macrocíclica de
bilactama
(azul). Los grupos
amida
de la forma de bilactama se intercambian en el ciclol, dando lugar a dos
tautómeros
con similar estabilidad, proporcionando una
constante de equilibrio
cercana a 1. Sin embargo, la forma abierta (en negro) es inestable y no ha sido observada.
[
100
]