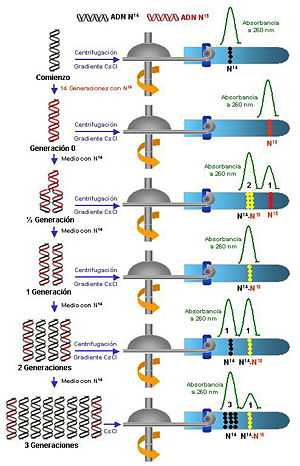
Una vez conseguido el primer objetivo, las células fueron transferidas a un medio que contenía nitrógeno-14, es decir, un medio más ligero, donde continuaron su crecimiento (división celular, que requiere la replicación del ADN).Se purificó el ADN y se analizó mediante una centrifugación en gradiente de cloruro de cesio, en donde hay más densidad en el fondo del tubo que en la parte media del mismo.En la primera generación (figura 2.b) se obtuvo una única banda de ADN con densidad intermedia.En la tercera generación se obtuvieron dos bandas, una ligera (con una abundancia del 75 %) y otra intermedia (con el 25 % restante).La banda intermedia o híbrida representa una molécula de ADN que contiene una cadena pesada (original) y otra ligera (recién sintetizada).Si fuera conservadora, aparecería siempre una banda pesada y el resto ligeras (figuras 1.a, 1.b, 1.c) .Si fuera dispersante solo aparecerían bandas híbridas de densidad intermedia en todas las generaciones.[2] Los experimentos realizados por Cairns (1963) con bacterias Escherichia coli permitieron determinar la existencia de ese punto fijo u origen de replicación a partir del cual el genoma empezaba a replicarse.Los experimentos consistían en mantener un cultivo de E. coli creciendo en un medio que contenía timidina tritiada (timina marcada con tritio), de forma que el ADN quedara marcado radiactivamente pudiendo efectuarse una autorradiografía.Los resultados indicaban que la replicación en E. coli se iniciaba en un punto concreto (OriC).Como los primeros genes en replicarse en la bacteria donadora serían los primeros en transferirse, el experimento permitió demostrar, a partir de las frecuencias relativas de los diferentes genes en las bacterias receptoras, que la replicación sigue un orden (es secuencial).No obstante, la replicación se puede considerar, de forma general, bidireccional.Estas proteínas se unen de forma cooperativa, por lo que su unión al ADN conforme avanza la helicasa es rápida.Por otro lado, conforme las helicasas van avanzando se van generando superenrollamientos en la doble cadena de ADN por delante de la horquilla y si éstos no fueran eliminados, llegado a un punto el replisoma ya no podría seguir avanzando.[2] En el siguiente paso, el ADN Pol III cataliza la síntesis de las nuevas cadenas añadiendo nucleótidos sobre el molde.Esta síntesis se da bidireccionalmente desde cada origen, con dos horquillas de replicación que avanzan en sentido opuesto.En la eliminación del cebador (también denominado iniciador o partidor) intervienen dos enzimas: por un lado el ADN Pol I, que va eliminando el ARN con su actividad exonucleasa 3' → 5' y simultáneamente rellenando con ADN mediante su actividad polimerasa 5' → 3' (proceso denominado nick-traslation).
Replicación de ADN. La doble hélice es desenrollada y cada hebra hace de plantilla para la síntesis de la nueva cadena. El ADN polimerasa añade los nucleótidos complementarios a los de la cadena original.
Tres posibles modelos de replicación. a) Conservadora, b) Dispersora, c) Semiconservadora (mecanismo real).
Distinción entre la replicación unidireccional y la bidireccional mediante el recuento de copias de
genes marcadores
. O es el
origen de replicación
y A, B, C, D, E son genes marcadores.
En la mayoría de los casos la replicación es bidireccional.