Triceratops
Aún no se ha encontrado un esqueleto completo de Triceratops,[4] aunque la criatura está bien documentada por numerosos restos parciales recogidos desde la introducción del género en 1887.
Aunque se vean tradicionalmente como armas defensivas contra depredadores, las últimas teorías proponen que es más probable que estas características fueron utilizadas durante el cortejo sexual y exhibiciones de dominancia, como la cornamenta de los modernos reno, la cabra blanca, o el escarabajo rinoceronte.
En los casmosaurinos, los premaxilares se reunían en su línea media en una placa ósea compleja, cuyo borde posterior estaba reforzado por el "puntal narial".
Estos eran procesos triangulares bajos en el borde del volante, que representaban osificaciones de piel separadas u osteodermos.
Algunos paleontólogos incluso han llegado a sugerir que una lesión en dicha gola puede haber resultado en extremo nociva para el animal, dada su alta irrigación sanguínea.
La piel del animal era gruesa y presentaba unas pequeñas protuberancias que se diseminaban por su cuerpo de manera irregular.
Interpretaciones posteriores revivieron una vieja hipótesis de John Bell Hatcher que muy al frente se puede observar un vestigio del atlas real, el sincervical entonces compuesto por cuatro vértebras.
Sin embargo, las icnitas (huellas de pisadas) atribuidas a Triceratops y las reconstrucciones recientes de esqueletos tanto reales como digitales, parecen demostrar que Triceratops mantuvo una postura vertical durante la locomoción normal, con los codos levemente arqueados hacia fuera, en una posición intermedia entre completamente vertical y completamente arqueado, como en el rinoceronte moderno.
[32] El vaquero Edmund B. Wilson se había sobresaltado al ver una calavera monstruosa que sobresalía del costado de un barranco.
En la primera tentativa de entender las muchas especies, Lull diferenció dos grupos, aunque no describió los caracteres en que se basó.
Estos resultados, sin embargo, fueron discutidos algunos años más tarde por Catherine Forster, quien reanalizó el material de Triceratops más exhaustivamente y concluyó que los restos encajaban en dos especies, T. horridus y T. prorsus, aunque el cráneo distintivo de T. hatcheri se diferenciaba lo suficiente como para justificar un género separado, hoy Nedoceratops.
El género Torosaurus coincide con Triceratops en edad geológica, distribución, anatomía y tamaño, siendo reconocido como su pariente más cercano.
Farke concluyó que Nedoceratops hatcheri corresponde a individuos adultos de su propio género, relacionado estrechamente con Triceratops.
Andrew Farke había subrayado en 2006 que, aparte del volante, no se podían encontrar diferencias sistemáticas entre Torosaurus y Triceratops.
[49] Sin embargo, cuestionó la conclusión de Scannella al argumentar en 2011 que los cambios morfológicos propuestos necesarios para "envejecer" un Triceratops en un Torosaurus no tendrían precedentes entre los ceratópsidos.
[51] Longrich estuvo de acuerdo con Scannella sobre Nedoceratops e hizo una sugerencia adicional, que Ojoceratops recientemente descrito también era un sinónimo.
En 1922, Henry Fairfield Osborn interpretó al recientemente descubierto Protoceratops como su antepasado,[61] pero pasaron muchas décadas antes de que nueva información saliera a luz.
Los dinosaurios se han definido como todos los descendientes del antepasado común más reciente de Triceratops y Neornithes, es decir los pájaros modernos.
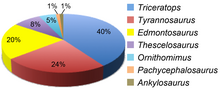
Mercuriceratops Judiceratops Chasmosaurus Mojoceratops Agujaceratops Pentaceratops aquilonius Casmosaurino de Williams Fork Pentaceratops sternbergii Utahceratops Kosmoceratops Anchiceratops Casmosaurino de la formación Almond Bravoceratops Coahuilaceratops Arrhinoceratops Titanoceratops Torosaurus Triceratops Aunque comúnmente se retrate a los Triceratops en manadas, como animales gregarios, realmente hay pocas pruebas.
Sus oídos estaban en sintonía con los sonidos de baja frecuencia dadas las longitudes cocleares cortas registradas en un análisis realizado por Sakagami et al.
Las dos teorías principales han girado alrededor de uso en combate, o exhibición en el cortejo, con esta última idea considerada como la función primaria más probable.
En su lugar se sugieren como causas la resorción ósea no patológica, u otras enfermedades de los huesos desconocidas.
[97] Una teoría similar fue propuesta para las placas del estegosaurio,[98] aunque este uso solamente no explicara la variación extraña y extravagante vista en diversos miembros de Ceratopsidae.
[101] En 2006, se publicó el primer estudio ontogenético extenso de Triceratops en la revista Proceedings of the Royal Society.
[102] Los ojos grandes y las características acortadas, un sello de "lindo" en los mamíferos bebé, también sugieren que los padres Triceratops pueden haber cuidado a sus jóvenes.
[107] Los tiranosáuridos de la formación son Nanotyrannus y Tyrannosaurus, aunque el primero podría ser un sinónimo menor del segundo.
Entre los ornitomimidos se encuentran los géneros Struthiomimus y Ornithomimus;[103] un animal no descrito llamado "Orcomimus" podría ser de la formación.
Se conocen tres anquilosaurianos, Ankylosaurus, Denversaurus y posiblemente una especie de Edmontonia o un género no descrito relacionado.
Entre ellos se encuentran los paquicefalosáuridos derivados Stygimoloch,[103] Dracorex,[113] Pachycephalosaurus,[103] Sphaerotholus, y un espécimen no descrito de Dakota del Norte.
[114] El aspecto distintivo del Triceratops ha llevado a que sean representados con frecuencia en películas, juegos de computadoras y documentales.