Neurociencia molecular
Los químicos transmitidos, conocidos como neurotransmisores, regulan una fracción significativa de las funciones vitales del cuerpo.
La dopamina, una catecolamina, fue identificada en el nematodo C. elegans por medio de esta técnica.
La presencia de neurotransmisores (aunque no sea necesariamente en su ubicación regular) puede ser observado por medio de inmunocitoquímica ligada a enzimas o por medio de un ensayo por inmunoabsorción ligado a enzimas en el que la unión del sustrato en los ensayos enzimáticos puede inducir [precipitado]]s, fluorocromos, o quimioluminiscencia.
[1] Las células excitables en los organismos vivos tienen canales iónicos dependientes de voltaje.
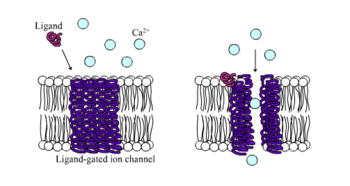
Los receptores GABAA regulan las rápidas respuestas de inhibición en el sistema nervioso central (SNC) y son encontrados en las neuronas, células gliales y células de la médula adrenal.
Los receptores GABA pueden también interaccionar con los ligandos no endógenos para influenciar en su actividad.
Por ejemplo, el compuesto diazepam (comercializado como Valium) es un agonista alostérico que incrementa la afinidad del receptor GABA.
El mecanismo de acción antagónico para este compuesto no es directamente sobre el receptor GABA, pero hay otros compuestos que son capaces de una inactivación alostérica, entre ellos está el T-butilbicicloforotionato (TBPS) y el pentilenetetrazol (PZT).
Estos receptores llevan el nombre de los agonistas que facilitan la actividad del glutamato.
Los receptores de NMDA son notables por sus mecanismos excitatorios para afectar la plasticidad neuronal en el aprendizaje y memoria, así como en neuropatologías como un accidente cerebrovascular y epilepsia.
Los receptores NMDA tienen múltiples sitios de unión como los receptores ionotrópicos GABA y pueden ser influenciados por co-agonistas como el neurotransmisor glicina o fenciclidina (PCP por sus siglas en inglés).
[5] El receptor nicotínico une el neurotransmisor acetilcolina para producir un flujo de canal catiónico no selectivo que genera respuestas excitatorias postsinápticas.
Normalmente estas respuestas lentas son caracterizadas por cambios bioquímicos intracelulares más elaborados.
Estas quantas han sido identificadas por microscopía electrónica como vesículas sinápticas.
Los dimorforfismos sexuales pueden estar significativamente influenciados por la expresión genética en el cerebro basada en el sexo la cual varía de una especie a otra.
Los modelos animales como los roedores, Drosophila melanogaster, y Caenorhabditis elegans, han sido usados para observar los orígenes y/o comparar las vías sexuales en el cerebro contra la producción de hormonas por las gónadas de un animal.
En los estudios con D. menlanogaster se encontró que gran parte de la expresión de las vías sexuales del cerebro ocurren inclusive después de que las gónadas son removidas, sugiriendo que las vías sexuales pueden ser independiente del control hormonal en algunos aspectos.
[9] Observar los genes sexuales parciales tienen un potencial para un significado clínico al observar la fisiología del cerebro y la relación potencial con los desórdenes neurológicos (ya sea directo o indirecto).
[9] Muchas funciones cerebrales pueden ser influenciadas a un nivel celular y molecular por las alteraciones y cambios en la expresión genética, sin alterar la secuencia de DNA en el organismo.
Estos cambios se han encontrado que son fuertemente influyentes en la incidencia de enfermedades cerebrales, desórdenes mentales, y adicción.
[10] El control epigenético se ha demostrado que está involucrado en los altos niveles de plasticidad en el desarrollo temprano, de ese modo se define du importancia en el periodo crítico del organismo.
Puede ser causado por la prolongada transmisión excitatoria sináptica en la cual los altos niveles del neurotransmisor glutamato provocan una excesiva activación en la neurona postsináptica que puede derivar en la muerte de dicha neurona.