Exocitosis
Todas las células utilizan la exocitosis y su contraparte, la endocitosis, porque la mayoría de las sustancias importantes para ellas son moléculas polares grandes, que no pueden atravesar la porción hidrofóbica de la membrana celular mediante el transporte pasivo.
Esta secreción es posible porque la vesícula se fusiona transitoriamente con la membrana plasmática.
[3] La exocitosis en las sinapsis químicas neuronales se desencadena por Ca2+ y sirve para la señalización interneuronal.
La sinaptotagmina ha sido reconocida como el sensor principal para la exocitosis desencadenada por Ca2+ en animales.
[4] Sin embargo, las proteínas sinaptotagmina están ausentes en plantas y eucariotas unicelulares.
Otros posibles sensores de calcio para la exocitosis son proteínas de mano EF (por ejemplo, calmodulina) y dominio C2 (por ejemplo, Ferlins, E-synaptotagmin, Doc2b) que contienen proteínas.
Todavía no está claro si la maquinaria entre la secreción constitutiva y regulada es diferente.
Dos complejos de anclaje están asociados con exocitosis constitutiva en mamíferos, ELKS y Exocyst.
El periplasma se pellizca como vesículas de la membrana externa bacteriana (OMV) para translocar señales bioquímicas microbianas en células huésped eucariotas[7] u otros microbios ubicados cerca,[8] logrando el control del microbio secretor en su entorno, incluida la invasión del huésped, endotoxemia, compitiendo con otros microbios por la nutrición, etc.
Por ejemplo, es probable que las vesículas que transportan proteínas desde el aparato de Golgi al área de la superficie celular utilicen proteínas motoras y una pista del citoesqueleto para acercarse a su objetivo.
Esto sugirió que durante el proceso secretor, solo una parte del contenido vesicular puede salir de la célula.
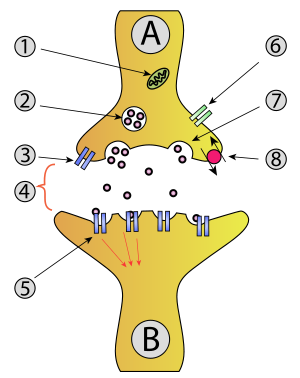
1. Mitocondria ,
2. Vesícula sináptica con neurotransmisores ,
3. Autoreceptor,
4. Sinapsis con neurotransmisor liberado ( serotonina ),
5. Receptores postsinápticos activados por neurotransmisores (inducción de un potencial postsináptico),
6. Canal de calcio ,
7. Exocitosis de una vesícula y
8. Neurotransmisor recapturado


