siRNA
En 1998, Andrew Fire, Craig C. Mello y colaboradores publicaron un trabajo en Nature,[8] en el cual indicaban que, si inyectaban solo ARN de hebra simple en el gusano, tanto sense como antisense, no se observaba ningún efecto.
Sin embargo, si inyectaban ARN doble hebra (dsRNA), observaban la supresión específica del gen objetivo.
Asimismo, en el mismo reporte, Fire y Mello establecieron que la interferencia de la expresión del gen objetivo se produce a nivel del ARNm: cuando inyectaron en gusanos dsRNA contra el gen mex-3, detectaron que el ARNm de mex-3 desaparece, y por tanto los gusanos no son viables.
Con experimentos similares se demostró que plantas infectadas con virus también contienen pequeños ARNs con secuencias virales.
En 2000, Phil Zamore, Tom Tuschl y colaboradores[11] desarrollaron un sistema in vitro para estudiar embriones de Drosophila.
Zamore, Tuschl y colaboradores fueron los primeros en mostrar que el ARNm incubado en un extracto se degrada en presencia de dsRNA.
[8] En otro experimento, Zamore, Tuschl y colaboradores[11] comprobaron si los dsRNAs son procesados en pequeños fragmentos de ~25 nt (como ocurre en plantas).
Este experimento mostraba además que ambas hebras del dsRNA (la sense y la antisense) se cortan en pequeños fragmentos, porque los fragmentos aparecen tanto cuando los dsRNAs han sido marcados solo en la hebra sense, como cuando has sido marcados sólo en la antisense.
Este experimento muestra que los dsRNAs incubados en el extracto son procesados en pequeños fragmentos de ~21-22 nts.
[13] Cuando se introducen en un organismo (por ejemplo, en plantas tras la infección con un virus de ARN, o simplemente inyectando dsRNA en plantas o nemátodos), el ARN exógeno es reconocido por la célula y unido a una enzima llamada Dicer, que contiene diferentes dominios.
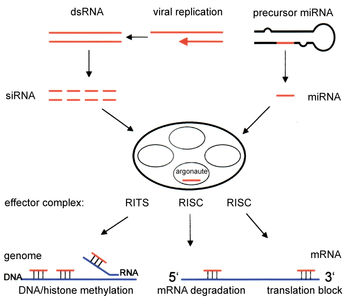
Estos pequeños dsRNAs (siRNAs) se incorporan en un complejo con múltiples componentes, denominado RISC (RNA-induced silencing complex).
A continuación, RISC* promueve el corte y posterior destrucción del ARNm diana, provocando la supresión de la expresión del gen (en la imagen de la derecha, el mecanismo que utilizan los siRNAs corresponde a la parte central e izquierda).
La siguiente cuestión que necesitaba resolverse era identificar la enzima ribonucleasa (ARNasa) involucrada en la fragmentación de los dsRNAs.
Como control negativo se utilizó la proteína "homeless", para probar si el dominio helicasa de Dicer podría estar implicado en la reacción.
[14] Los siRNAs generados por Dicer se ensamblan en el complejo RISC (siglas en inglés de RNA-induced silencing complex) que contiene diferentes factores celulares.
[20][21] La hebra seleccionada como guía tiende a ser aquella cuyo extremo 5' se aparea menos con su complementaria,[22] aunque la selección de la hebra guía no resulta afectada por la dirección en la que Dicer corta el dsRNA antes de su incorporación al complejo RISC.
[26] Todavía no está claro cómo el complejo RISC activado localiza los ARNm complementarios en el interior celular.
Un complejo de silenciamiento inducido por ARN (el complejo RISC) selecciona entre ambas hebras del siRNA: la hebra sense (pasajera) se degrada, mientras que la antisense (guía) se utiliza para localizar el ARNm complementario, y puede tener distintos destinos según el organismo.
En Drosophila y en mamíferos, la hebra antisense se incorpora directamente en el complejo RISC para identificar el mRNA complementario, que será destruido.
También en este caso, el ARNm unido a siRNAs es destruido, produciendo el silenciamiento (knock-down) del gen correspondiente.
En este momento, el material inyectado se diluye hasta solamente algunas moléculas de ARN doble hebra por célula.
Este complejo identifica el ARNm homólogo del siRNA, cortándolo en dos mitades que son después degradadas por la maquinaria celular.
Schizosaccharomyces pombe (la levadura de fisión) es una levadura ampliamente utilizada en estudios genéticos como organismo modelo (por ejemplo, es el organismo en el que Paul Nurse realizó los estudios sobre el ciclo celular que le permitieron obtener el Premio Nobel de Fisiología o Medicina en 2001).
Los elementos repetitivos que rodean al centrómero (cen) son transcritos en S. pombe para producir cenRNAs no-codificantes.
La proteína kinasa dependiente de dsRNA (PKR) es un importante "detector" antiviral, que se activa al unirse a dsRNAs largos, como los producidos por los virus.
PKR también activa las enzimas 2',5'-OligoA sintetasa y ARNasa L, que son componentes centrales de una importante vía antiviral.
Es interesante destacar que la ruta del RNAi se puede modular extracelularmente utilizando moléculas químicas pequeñas.
Aunque la administración es más difícil en otros organismos, también se están haciendo esfuerzos para realizar estudios genómicos a gran escala en células de mamíferos.
Todas estas son consideraciones que están bajo intenso estudio en la actualidad, para reducir su impacto en la utilización terapéutica del RNAi.
[82] Esfuerzos similares se han dirigido hacia la reducción del producto natural cianogénico linamarino en plantas de yuca.