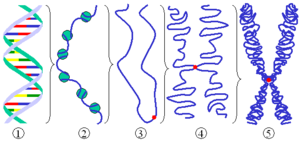
Cromatina
La cromatina[1] es la forma en la que se hace observable el material genético en el núcleo celular.
En 1879 Walther Flemming fue el primero en describir la cromatina; le otorgó este nombre debido a su afinidad por los colorantes.
[2] En 1869 Miescher había aislado una sustancia molecular fosforada a la que llamó nucleína.
En la eucromatina inactiva, esta fibra se enrolla sobre sí misma gracias a las histonas H1 para formar el solenoide.
Es una forma inactiva condensada localizada sobre todo en la periferia del núcleo celular, que se tiñe fuertemente con las coloraciones de histología.
En 1928, Emil Heitz, basándose en observaciones histológicas, definió la heterocromatina como los segmentos cromosómicos que aparecían muy condensados y oscuros en el núcleo en interfase.
Por ejemplo, en Schizosaccharomyces pombe, la heterocromatina se forma en el centrómero, telómeros y en el loci mating-type.
Durante mucho tiempo el papel concreto de la heterocromatina ha sido un misterio, ya que su polimorfismo no parecía tener ningún efecto funcional o fenotípico.
La cromatina es una estructura dinámica que adapta su estado de compactación y empaquetamiento para optimizar los procesos de replicación, transcripción y reparación del ADN, juega un rol regulatorio fundamental en la expresión génica.
Las distintas histonas pueden sufrir modificaciones post-traduccionales, como ser la metilación, acetilación, fosforilación, generalmente dada en residuos lisina o arginina.
La acetilación está asociada con activación de la trascripción, ya que al acetilarse una lisina, disminuye la carga positiva global de la histona por lo cual tiene una menor afinidad por el ADN (que está cargado negativamente).
En consecuencia, el ADN se encuentra unido menos fuertemente lo que permite el acceso de la maquinaria transcripcional.
Por el contrario, la metilación está asociada con la represión transcripcional y una unión ADN-histona más fuerte (si bien no siempre esto se cumple).
La carga de mutaciones es mayor en aquellas zonas que presentan cromatina reprimida y en regiones donde la replicación es tardía, lo cual se ha observado también en cáncer humano.
Estos métodos cuantifican el número de interacciones de loci genómicos que están cerca en le estructura 3D, pero pueden encontrase lejos respecto a la secuencia nucleotídica.Se han desarrollado muchos métodos para estudiar estos contactos genómicos y todos comparten unos pasos iniciales:[7] 1.
En concreto, se modifica el paso de ligación con biotina del método Hi-C.
Ésta postula que dos haplotipo homólogos deben tener diferentes patrones de contacto y que, por tanto, los haplotipos desconocidos de un cromosoma deben contactar en una región cromosómica cercana o vecina.
De este modo, se consigue establecer qué haplotipo celular está implicado en cada contacto cromosómico.
El estudio llevado con este método confirma que detecta más contactos entre los cromosomas y un menor número de falsos positivos.