Durante mucho tiempo se pensó que el calamar gigante ( Architeuthis dux , en la foto) era el cefalópodo más grande que existe. Ahora se sabe que el calamar colosal ( Mesonychoteuthis hamiltoni ) alcanza un tamaño máximo aún mayor. El calamar gigante que se ve aquí medía 9,24 m (30,3 pies) de longitud total y tenía una longitud de manto de 1,79 m (5,9 pies).
Los cefalópodos , que incluyen calamares y pulpos, varían enormemente en tamaño. Los más pequeños miden sólo alrededor de 1 centímetro (0,39 pulgadas) de largo y pesan menos de 1 gramo (0,035 oz) en la madurez, mientras que el calamar gigante puede superar los 10 metros (33 pies) de largo y el calamar colosal pesa cerca de media tonelada ( 1.100 libras), lo que los convierte en los invertebrados vivos más grandes . Las especies vivas varían en masa más de tres mil millones de veces, [nb 1] o nueve órdenes de magnitud , desde las crías más ligeras hasta los adultos más pesados. [4] Ciertas especies de cefalópodos también se caracterizan por tener partes individuales del cuerpo de tamaño excepcional. [5]
Los cefalópodos fueron en algún momento los organismos más grandes de la Tierra, [6] y a partir del registro fósil se conocen numerosas especies de tamaño comparable al de los calamares más grandes de la actualidad , incluidos enormes ejemplos de amonoides , belemnoides , nautiloideos , ortoceratoides , teutidos y vampiromorfidos . En términos de masa, los cefalópodos más grandes conocidos probablemente fueron los amonoides gigantes con caparazón y los nautiloideos endoceridos , [7] aunque quizás todavía estén en segundo lugar después de los cefalópodos vivos más grandes cuando se considera solo la masa de tejido . [8]
En varias ocasiones se han postulado cefalópodos mucho más grandes que los calamares gigantes o colosales. Uno de ellos fue el Monstruo de San Agustín , un gran cadáver que pesaba varias toneladas y que llegó a la costa de Estados Unidos cerca de San Agustín , Florida , en 1896. Se realizaron nuevos análisis en 1995 y 2004 de las muestras de tejido originales, junto con las de otros cadáveres similares demostraron de manera concluyente que todos eran masas de la matriz colagenosa de la grasa de ballena . [9]
Los cefalópodos gigantes han fascinado a la humanidad durante siglos. Los registros más antiguos que se conservan son quizás los de Aristóteles y Plinio el Viejo , quienes describieron calamares de gran tamaño. [10] Los cuentos sobre calamares gigantes han sido comunes entre los marineros desde la antigüedad y pueden haber inspirado el monstruoso kraken de la leyenda nórdica , del que se dice que es tan grande como una isla y capaz de hundir y hundir cualquier barco. [11] Se conocen monstruos marinos con tentáculos similares en otras partes del mundo, incluidos los Akkorokamui de Japón y Te Wheke-a-Muturangi de Nueva Zelanda . La Lusca del Caribe y Scylla en la mitología griega también pueden derivar de avistamientos de calamares gigantes, [12] al igual que relatos de testigos presenciales de otros monstruos marinos como las serpientes marinas . [13]
Medidas estándar para cefalópodos con calamares y pulpos como ejemplos
Longitud del manto
Los científicos miden el ancho del manto de un gran calamar gigante hembra de c. 2 m (6,6 pies) ML
La longitud del manto (ML) es la medida de tamaño estándar para los cefalópodos coleoides (el diámetro de la concha es más común en los nautilos ) y se informa casi universalmente en la literatura científica. El manto es el "cuerpo" del cefalópodo, situado detrás de la cabeza y que encierra la masa visceral y la cavidad del manto , siendo esta última utilizada para la locomoción mediante propulsión a chorro . A menos que se indique lo contrario, la longitud del manto se mide dorsalmente sobre la línea media del manto (a veces especificada como longitud del manto dorsal , LMD). Es una medida recta, no medida sobre la curva del cuerpo. En los Decapodiformes (cefalópodos de diez extremidades), la longitud del manto se mide desde el borde anterior del manto (cerca de la cabeza), hasta el extremo posterior del manto o el ápice de las aletas unidas , el que sea más largo. En Octopodiformes (cefalópodos de ocho extremidades), el borde anterior del manto no está claramente delimitado dorsalmente debido a la avanzada fusión cabeza-manto y, por lo tanto, la longitud del manto se toma desde el punto medio entre los ojos hasta el extremo posterior del manto. Cuando se refiere a longitud del manto ventral en lugar de dorsal, esto siempre se especifica como tal y se abrevia VML. [18]
Como indicación del tamaño total, la longitud del manto generalmente se considera más confiable que la longitud total porque las extremidades de los cefalópodos pueden estirarse fácilmente más allá de su longitud natural y, a menudo, están dañadas o faltan en los especímenes preservados (esto es particularmente cierto en el caso de los largos tentáculos de muchas especies de calamares). ). [19] Sin embargo, la longitud del manto no es igualmente aplicable a todas las especies. Ciertos pulpos bentónicos como Callistoctopus ornatus son capaces de alargar y retraer su manto y, por tanto, las medidas de la longitud del manto, incluso cuando se toman de un espécimen vivo, pueden variar considerablemente. Otro caso problemático es el de los cirroteútidos gelatinosos , cuyos mantos débilmente musculosos son propensos a contraerse sustancialmente durante la conservación. La distancia interocular puede ser un estándar más fiable para este grupo. [20]
Largo total
La longitud total (TL) se mide a lo largo de la línea media dorsal con las extremidades extendidas y en línea con el eje del cuerpo. Es la mayor extensión mensurable de un espécimen desde el extremo posterior del manto o las aletas (o cola , si está presente [21] ) hasta el ápice de la extremidad más larga. [18] Se recomienda medir los brazos y los tentáculos en un estado relajado para no exagerar su longitud, pero históricamente esta práctica no siempre se siguió y algunas de las mediciones más extremas publicadas de calamares gigantes se han atribuido al alargamiento artificial de la tentáculos. [22] Aunque la longitud total se menciona a menudo en relación con las especies de cefalópodos más grandes, rara vez se utiliza en teutología. [23] Al igual que con la longitud del manto, es una medida en línea recta.
La longitud total no debe confundirse con la extensión de los brazos, también conocida como extensión de los brazos , extensión radial o extensión radial , que puede ser mucho mayor y a menudo se informa en el caso de los pulpos (para los cuales los brazos suelen constituir la gran mayoría de la longitud). En los calamares, la longitud total incluye los tentáculos alimentarios, que en algunas especies pueden ser más largos que el manto, la cabeza y los brazos combinados ( los quiroteutidos como Asperoteuthis acanthoderma son un buen ejemplo).
Una medida relacionada es la longitud estándar (SL), que es la longitud combinada del manto, la cabeza y los brazos, excluyendo los tentáculos alimentarios, a menudo largos. [24] Esta medida es especialmente útil para especies como el calamar gigante, donde casi toda la masa del animal ocupa menos de la mitad de su longitud total.
Masa
La masa (a menudo abreviada WT para "peso") se informa con mucha menos frecuencia que el manto o la longitud total, y no existen registros precisos para todas las especies de cefalópodos grandes. También puede variar ampliamente según el estado de la muestra en el momento del pesaje (por ejemplo, si se midió viva o muerta, húmeda o seca, congelada o descongelada, antes o después de la fijación, con o sin masa de huevo, etcétera).
Pico de calamar gigante y músculos asociados con mano para escalar
Métodos de determinación del tamaño.
A diferencia de la gran mayoría de los cefalópodos actuales, que son totalmente de cuerpo blando, la determinación del tamaño de las pocas especies con caparazón supervivientes (en términos de diámetro del caparazón) es comparativamente sencilla y puede lograrse con un alto nivel de precisión. Cualquiera que sea el tipo de cefalópodo, a falta de especímenes completos, el tamaño a menudo puede estimarse sólo a partir de restos parciales. Por ejemplo, los picos de los cefalópodos se pueden utilizar para estimar la longitud del manto, la longitud total y la masa corporal, [25] y este método se ha utilizado en particular para estimar el tamaño máximo del calamar colosal . La longitud rostral inferior (LRL) del pico se utiliza a menudo para este propósito. La longitud rostral de los picos superior e inferior es la medida estándar del tamaño del pico en Decapodiformes ; Se prefiere la longitud de la capucha para los Octopodiformes . [26]
La longitud del manto se ha estimado a partir de grabaciones de vídeo de calamares en estado salvaje. [27]
Etapas tempranas de la vida
Crías
Hilo de huevos y embriones de Haliphron atlanticus , recolectados al norte de las islas de Cabo Verde . Los huevos miden alrededor de 8 mm (0,31 pulgadas) a lo largo del eje longitudinal.
Las crías de Idiosepius thailandicus , posiblemente la especie de cefalópodo existente más pequeña en la madurez, tienen una longitud de manto de alrededor de 1 mm (0,039 pulgadas). [28] El Idiosepius pygmaeus, estrechamente relacionado, pesa sólo 0,00033 g (1,2 × 10 −5 oz) al nacer y aumenta de peso a 0,175 g (0,0062 oz) a medida que alcanza la madurez en 50 días. [2] Aún más pequeñas son las crías del Illex illecebrosus , de importancia comercial , con una masa de 0,00015 g (5,3 × 10 −6 oz). [29] Las crías del pulpo gigante del Pacífico ( Enteroctopus dofleini ), una de las dos especies de pulpo más grandes, pesan 0,0253 g (0,00089 oz) en promedio. [30]
En el otro extremo se encuentran los nautilos , que al nacer suelen tener un diámetro de caparazón de 25 mm (1 pulgada) o más (dependiendo de la especie), el mayor tamaño de cría entre los invertebrados existentes. [31] Se estima que las crías de Nautilus belauensis , una de las especies más grandes, pesan del orden de 5,9 g (0,21 oz) [nb 2] y maduran alrededor de 1,2 kg (2,6 lb) después de casi 4000 días, o alrededor de 11 años. [2]
Idiosepius paradoxus , una pequeña especie de calamar que crece hasta 16 mm (0,63 pulgadas) de longitud del manto
Tamaño adulto más pequeño
El tamaño adulto más pequeño entre los cefalópodos vivos lo alcanzan los llamados calamares pigmeos, Idiosepius , [32] y ciertas especies diminutas del género Octopus , los cuales pesan menos de 1 gramo (0,035 oz) en la madurez. [33] Idiosepius thailandicus es quizás el más pequeño de todos, con las hembras con un promedio de 10,4 mm (0,41 pulgadas) de longitud del manto y los machos con 5,9 mm (0,23 pulgadas). [3] Los pesos húmedos promedio son de alrededor de 0,20 y 0,02 g (0,00705 y 0,00071 oz), respectivamente. [3]
La superfamilia de octópodos Argonautoidea se caracteriza por tener machos marcadamente enanos. [34] Los cuatro géneros existentes del grupo son Argonauta , Haliphron , Ocythoe y Tremoctopus , todos los cuales son exclusivamente pelágicos . La mayor disparidad en el tamaño de los sexos se observa en los pulpos manta del género Tremoctopus . Norman et al. (2002) informaron sobre un Tremoctopus violaceus macho completamente maduro que medía 2,4 cm (0,94 pulgadas) de longitud total y pesaba apenas 0,25 g (0,0088 oz). En comparación, las hembras grandes de esta especie alcanzan una longitud total de 2 m (6,6 pies) y probablemente unos 10 kg (22 lb) de peso. Este es el dimorfismo de tamaño sexual más extremo conocido entre animales no microscópicos, [nb 3], siendo las hembras maduras al menos 10.000 veces más pesadas que los machos, y probablemente hasta 40.000 veces más pesadas. [35] Los géneros relacionados Argonauta y Ocythoe tienen machos igualmente pequeños, pero las hembras no son tan grandes como las de Tremoctopus y, por lo tanto, el dimorfismo de tamaño es menos pronunciado. Las hembras del cuarto género argonautoide, Haliphron , son las más grandes de todos, con una longitud de 3,5 m (11 pies) y una masa de 75 kg (165 lb) (y posiblemente los pulpos más grandes de cualquier tipo), pero los machos también son mucho más grande, de hasta 30 cm (12 pulgadas). Por tanto, el dimorfismo de tamaño es el menos pronunciado de todos. [36]
Taxones extintos
Se conocen numerosas especies de las denominadas amonitas micromórficas. [37] Maximitas del Carbonífero Superior es el ammonoide más pequeño conocido. Los especímenes adultos alcanzaron sólo 10 mm (0,39 pulgadas) de diámetro de concha. [38]
Los calamares son los cefalópodos vivos más grandes en términos de longitud del manto , longitud total y masa, siendo la especie más grande en al menos dos de estas medidas el calamar colosal , Mesonychoteuthis hamiltoni . Con una longitud estimada del manto de 3 m (9,8 pies) y 10 m (33 pies) de longitud total, y un peso de hasta 495 kg (1091 lb), esta especie es también la más grande de todos los invertebrados existentes . [39] El único otro calamar que se acerca a estas dimensiones es el calamar gigante del género Architeuthis , con hembras de hasta 275 kg (606 lb), 2,4 m (7,9 pies) de longitud del manto y posiblemente hasta 15 m (49 ft) de longitud total, lo que lo convierte probablemente en el más largo de todos los cefalópodos. [40] Las dos especies de pulpo más grandes , Enteroctopus dofleini y Haliphron atlanticus , pueden superar los 70 kg (150 lb), y la primera tiene una longitud total máxima de más de 6 m (20 pies). Los octópodos cirrados (con aletas) también pueden alcanzar un gran tamaño; el espécimen capturado más grande probablemente sea un Cirrothauma magna de 1,7 m (5,6 pies) de longitud total y 33 cm (13 pulgadas) de longitud del manto, [41] aunque las observaciones desde sumergibles sugieren que Los miembros de este grupo pueden superar los 4 m (13 pies) de longitud total. [42]
Los miembros de los otros grupos de cefalópodos son sustancialmente más pequeños, aunque la sepia más grande puede superar los 10 kg (22 lb) de peso y los 50 cm (1,6 pies) de longitud del manto. Se conocen cefalópodos de tamaño comparable al calamar más grande de la actualidad a partir de restos fósiles, incluidos enormes ejemplos de amonoides , belemnoides , nautiloideos , ortoceratoides , teutidos y vampiromorfidos . [43]
Calamar colosal ( Mesonychoteuthis hamiltoni )
Pico de un calamar colosal, que tiene el pico más grande entre los cefalópodos vivos.
Aunque se ha registrado un número sustancial de restos de calamar colosal ( Mesonychoteuthis hamiltoni ) (Xavier et al., 1999 recopilaron 188 posiciones geográficas para especímenes totales o parciales capturados en pesquerías comerciales y científicas), se han documentado muy pocos animales adultos o subadultos. lo que dificulta estimar el tamaño máximo de la especie. McClain y cols. (2015) afirmaron que solo se conocían 12 especímenes "completos". El espécimen completo más grande conocido de calamar colosal fue una hembra madura capturada en el Mar de Ross en febrero de 2007. Su peso se estimó inicialmente en 450 kg (990 lb), la longitud de su manto en 4 m (13 pies) y su longitud total. a 8 a 10 m (26 a 33 pies). [44] Una vez completamente descongelado, se descubrió que el espécimen pesaba 495 kg (1091 lb), pero medía solo 2,5 m (8,2 pies) de longitud del manto y 4,2 m (14 pies) de longitud total. [45] Es probable que el espécimen, y particularmente sus tentáculos, se redujeran considerablemente post mortem como resultado de la deshidratación, después de haber estado en un congelador durante 14 meses. (Según lo informado por el Museo de Nueva Zelanda Te Papa Tongarewa , los especímenes de Nototodarus sloanii , el calamar flecha de Nueva Zelanda, pueden encogerse hasta en un 22 % cuando se deshidratan con soluciones alcohólicas. [45] ) El colosal espécimen de calamar se contrajo por un 5% después de varios años en líquido conservante (primero formalina y luego propilenglicol ). [46] Las aletas del espécimen del Mar de Ross de 2007 medían alrededor de 1,2 m (3,9 pies) de ancho y tenía un ancho de manto de 98,2 cm (3,22 pies). [47] Los brazos tenían una longitud de 0,85 a 1,15 m (2,8 a 3,8 pies), mientras que los dos tentáculos medían alrededor de 2,1 m (6,9 pies) de largo. [48]
Los picos recuperados de los estómagos de los cachalotes indican la existencia de animales que superan incluso al espécimen del Mar de Ross de 2007. Ese espécimen tenía una longitud rostral inferior (LRL) de 42,5 mm (1,67 pulgadas) y pesaba 495 kg (1091 lb), [49] mientras que la hembra submadura de 300 kg (660 lb) de 2003 tenía un LRL de 37 mm (1,5 en). [50] En comparación, el pico de calamar colosal más grande conocido de un estómago de cachalote medía 49 mm (1,9 pulgadas) en LRL. [22] Aunque el número de especímenes de calamar grandes y colosales conocidos por la ciencia es demasiado pequeño para tener una buena idea de la relación entre el tamaño del pico y el tamaño corporal general, un pico de tal enormidad indica un animal verdaderamente masivo que pesa quizás tanto como 600– 700 kg (1300 a 1500 libras). [49] Sin embargo, la relación de escala para esta especie muestra una latitud considerable, como lo demuestra un pico de 40 mm (1,6 pulgadas) LRL extraído de un animal que pesa solo 160 kg (350 lb). [49]
Calamar gigante ( Architeuthis dux )
Un ejemplar de calamar gigante; Los tentáculos alimenticios extremadamente largos del calamar gigante lo convierten en el cefalópodo más largo conocido, del pasado o del presente.
El tamaño máximo del calamar gigante ( Architeuthis dux ) ha sido durante mucho tiempo un tema tanto de debate popular como de investigación académica. [51] A diferencia del calamar colosal, el calamar gigante se conoce a partir de un número sustancial de especímenes maduros. El número total de especímenes registrados (en todas las etapas de desarrollo) se acerca a mil, con aproximadamente 700 documentados en 2015 [update], de los cuales alrededor de 460 habían sido medidos de alguna manera. [52] Desde entonces, este número ha aumentado sustancialmente, con 57 especímenes registrados en aguas japonesas durante un período excepcional de 15 meses entre 2014 y 2015. [53]
Un calamar gigante que mide 7 m (23 pies) de longitud total y pesa 250 kg (550 lb) exhibido en un bloque de hielo de 3,5 toneladas (7700 lb) en el Acuario de Melbourne [54]
Basado en un conjunto de datos de 40 años de más de 50 especímenes de calamar gigante, Roper & Shea (2013:114) sugieren una longitud total promedio en la madurez de 11 m (36 pies) y una "longitud máxima rara vez encontrada" de 14 a 15 m (46 a 49 pies). De los casi 100 especímenes examinados por Clyde Roper , el más grande medía "46 pies (14 m) de largo". [55] O'Shea y Bolstad (2008) dan una longitud total máxima de 13 m (43 pies) para las hembras basándose en el examen de más de 130 especímenes, medidos post mortem y relajados, así como picos recuperados de cachalotes ( que no superan el tamaño de los encontrados en los ejemplares completos de mayor tamaño). Steve O'Shea estimó la longitud total máxima de los machos en 10 m (33 pies). [56] Los registros más antiguos de 18 m (59 pies) o más probablemente fueron exagerados por el estiramiento de los largos tentáculos de alimentación o fueron el resultado de métodos de medición inadecuados, como el ritmo. [57] O'Shea ha declarado que, dada la evidencia disponible, el límite superior más alto que consideraría plausible para la longitud total del calamar gigante sería 15 m (49 pies), y que la probabilidad de que existan calamares gigantes de 20 metros es "tan extremadamente remoto que no se podría justificar el esfuerzo de escribir sobre ello". [58]
Incluyendo la cabeza y los brazos pero excluyendo los tentáculos (longitud estándar, SL), la especie rara vez supera los 5 m (16 pies) según O'Shea y Bolstad (2008). Paxton (2016a) considera que 9,45 m (31,0 pies) es el mayor SL medido de manera confiable, basándose en un espécimen reportado por Verrill (1880a:192), y considera que los especímenes de 10 m (33 pies) SL o más son "muy probables". ", pero estas conclusiones han sido criticadas por expertos en calamares gigantes. [59] O'Shea (2003a) estimó el peso máximo de las hembras de calamar gigante en 275 kg (606 lb), basándose en el examen de unos 105 especímenes, así como en los picos recuperados de cachalotes (que no superan el tamaño de aquellos encontrado en los especímenes completos más grandes). Los calamares gigantes tienen dimorfismo sexual en tamaño , con un peso máximo para los machos estimado en 150 kg (330 lb), [56] aunque ocasionalmente se han reportado especímenes más pesados (como un espécimen de 190 kg (420 lb) [60] y un ejemplar de 163 kg (359 lb) espécimen [61] ). Roper y Jereb (2010a:121) dan un peso máximo de hasta 500 kg (1100 lb), y "posiblemente mayor". Pesos desacreditados de hasta una tonelada (2200 libras) o más no son infrecuentes en la literatura antigua (ver más abajo). [22]
Otros taxones de calamar
Calamar pulpo dana
La tercera especie de calamar más pesada que existe es Taningia danae , también conocida como calamar pulpo Dana. El espécimen más grande y bien documentado es una hembra madura de 160 cm ML reportada por Roper y Vecchione (1993) en el Atlántico Norte. El artículo original dio la masa de este espécimen como 61,4 kg (135 lb), pero según Roper & Jereb (2010h:266) esta cifra es incorrecta y se debe a un error tipográfico, siendo la masa correcta 161,4 kg (356 lb). Sin embargo, Roper y Vecchione (1993) fueron consistentes en el uso de la cifra de 61,4 kg. Otro espécimen igualmente grande, una hembra que pesaba 124 kg (273 lb), fue reportado en aguas del norte de España por González et al. (2003:297) (ver también informes iniciales [62] ). En julio de 2010, un cachalote fue fotografiado frente a la isla de Faial , en las Azores, con un calamar grande, probablemente T. danae , en la boca. El ancho máximo del espécimen, de punta a punta de aleta, se estimó en 1,5 a 2 m (4,9 a 6,6 pies); esto se aproximaría a la longitud de su manto. [63]
Onykia robusta crece hasta una longitud de manto de al menos 2 m (6,6 pies)
Onykia robusta , anteriormente conocida como Moroteuthis robusta [64] y a veces llamada calamar garfio robusto, tiene una longitud de manto de hasta 200 cm (6,6 pies). [65] Algunos registros más antiguos superan esto, como el ML de 91,5 pulgadas (232 cm) informado por Verrill (1876:237) de un espécimen con una longitud total de 14 pies (4,3 m) (excluyendo los extremos de los tentáculos, que había sido destruido). Nesis (1987:192) también dio una longitud máxima del manto de 230 cm (7,5 pies), pero Roper y Jereb (2010i:364) escribieron que "este antiguo registro podría estar equivocado", y la especie comúnmente crece hasta 160 cm ( 5,2 pies) ML. Glaubrecht y Salcedo-Vargas (2004:66) proporcionaron una longitud total máxima de 4 a 6 m (13 a 20 pies). Las fuentes bibliográficas dan un peso máximo de 50 kg (110 lb). [66] Existen numerosos registros publicados de individuos grandes de esta especie. [67]
Pescador sacando un calamar de Humboldt frente a las costas de Chile . Esta especie es el calamar más grande que los humanos encuentran comúnmente. [68]
El calamar de Humboldt ( Dosidicus gigas ), también conocido como 'calamar gigante', crece hasta una longitud máxima del manto de al menos 120 cm (3,9 pies), [69] si no 150 cm (4,9 pies). [70] Los animales más grandes se encuentran frente a la costa occidental de América del Sur ; las poblaciones del norte alcanzan los 100 cm (3,3 pies) de ML y, en general, 50 a 80 cm (1,6 a 2,6 pies) de ML es más típico para la especie. [71] Las poblaciones del sur pueden tener una longitud total cercana a los 2,5 m (8,2 pies), [71] y posiblemente hasta 3,7 m (12 pies). [72] Nuevamente, los especímenes del hemisferio norte son mucho más pequeños, y los de la costa de California alcanzan longitudes totales de menos de 1,7 m (5,6 pies). [73] El calamar de Humboldt comúnmente alcanza un peso de alrededor de 20 a 30 kg (44 a 66 lb) [71] y puede alcanzar un máximo de 50 kg (110 lb). [74] Hay informes anecdóticos de animales individuales mucho más grandes, incluido el del buzo Scott Cassell , que ha buceado con calamares de Humboldt más de 300 veces (según se informa, más que cualquier otra persona). [75] [nota 4]
Kondakovia longimana , a veces conocido como calamar verrugoso gigante, es una especie poco conocida con una distribución circunantártica en el Océano Austral . El espécimen completo más grande, encontrado flotando en la superficie frente a las Islas Orcadas del Sur , tenía una longitud de manto de 108 cm (3,54 pies), [76] pero un espécimen femenino dañado con una longitud de manto estimada de alrededor de 150 cm (4,9 pies) es conocido. [77] El espécimen completo más grande tenía un peso húmedo de 29 kg (64 lb). [76] El peso máximo de la especie se ha estimado en 50 kg (110 lb). [78]
Octópodos más grandes
El pulpo gigante del Pacífico es una de las dos especies de pulpos más grandes.
El pulpo gigante del Pacífico ( Enteroctopus dofleini ) crece hasta más de 6,1 m (20 pies) de longitud total [79] y al menos 60 cm (2,0 pies) de longitud del manto. [80] Cosgrove (1987) y Cosgrove & McDaniel (2009:69) dieron un peso máximo confirmado de 71 kg (157 lb) para un espécimen vivo recolectado a mediados de la década de 1960. [40] Norman et al. (2014:124) aceptan un peso máximo de al menos 180 kg (400 lb), que se aproxima a los 182,3 kg (402 lb) reportados para un espécimen capturado en Santa Bárbara , California , en 1945, del cual sobrevive evidencia fotográfica. [81] No se han reportado especímenes que se acerquen a este tamaño desde mediados del siglo XX, y los especímenes recientes rara vez superan los 50 kg (110 lb). [82] Es posible que el tamaño máximo de la especie haya disminuido durante este período, tal vez debido a la bioacumulación de tóxicos (ver más abajo). [83]
En 2002, un espécimen gigante de Haliphron atlanticus , el pulpo de siete brazos, fue capturado por pescadores que pescaban con redes de arrastre a una profundidad de 920 m (3020 pies) frente a la zona oriental de Chatham Rise , Nueva Zelanda . Este ejemplar, el más grande de esta especie y posiblemente de todos los pulpos, fue el primer registro validado de Haliphron del Pacífico Sur . Tenía una longitud de manto de 0,69 m (2,3 pies), una longitud total de 2,90 m (9,5 pies) y un peso de 61,0 kg (134,5 libras), aunque estaba incompleto. [84] La longitud total del espécimen, una vez completo, se ha estimado en 4 m (13 pies) y su peso en 75 kg (165 lb). [85]
Taxones extintos
El espécimen más grande conocido de Parapuzosia seppenradensis , con su autor que lo describió, Hermann Landois , quien reconstruyó su cámara desaparecida con una combinación de estructura de alambre y papel. [86] Landois asumió que la cámara viva constituía un cuarto del verticilo exterior , pero Teichert y Kummel (1960:6) estimaron que continuaba durante tres cuartos a un verticilo completo, lo que daría un diámetro de concha original de alrededor de 3,5 m (11 pies).
Ciertos cefalópodos extintos rivalizaban o incluso superaban el tamaño de las especies vivas más grandes. [87] En particular, se sabe que la subclase Ammonoidea ha incluido un número considerable de especies que pueden considerarse "gigantes" (definidas por Stevens, 1988 como aquellas que superan 1 m (3,3 pies) de diámetro de concha). La amonita más grande confirmada , un espécimen de Parapuzosia seppenradensis descubierto en una cantera alemana en 1895, mide 1,742 m (5,72 pies) de diámetro, [88] aunque su cámara habitable falta en gran medida. El diámetro de la concha completa se ha estimado en 2,55 m (8,4 pies), suponiendo que la cámara habitable ocupara una cuarta parte del verticilo exterior . [89] Teichert y Kummel (1960:6) sugirieron un diámetro de concha original aún mayor, de alrededor de 3,5 m (11 pies) para este espécimen, asumiendo que la cámara del cuerpo se extendía desde tres cuartos hasta un verticilo completo. En 1971, se informó que se encontró una porción de una amonita que posiblemente superaba a este espécimen en una fábrica de ladrillos en Bottrop , Alemania occidental. [90] Se decía que un espécimen encontrado por Jim Rockwood, del Triásico Tardío cerca del lago Williston , Columbia Británica , medía más de 8 pies (2,4 m) de ancho, pero luego se determinó que era una concreción . [91]
Se sabe que los amonites heteromorfos también excedían 1 m (3,3 pies) de longitud, pero como sus caparazones estaban desenrollados en diversos grados, en general eran mucho más pequeños que los no heteromorfos más grandes. Las mayores longitudes de todas las alcanzaron los ortoconos de nautiloideos endocéridos como Endoceras , que pueden haber excedido los 8 m (26 pies), aunque su tamaño máximo es incierto; Si bien el fósil de endocérido más grande y bien documentado es probablemente el fragmento de concha de 3 metros de largo (9,8 pies) que se encuentra en el Museo de Zoología Comparada de la Universidad de Harvard , hay informes publicados de especímenes aún más grandes. Teichert (1927) mencionó especímenes de hasta 5 m (16 pies) de largo de la piedra caliza del Ordovícico Medio de Estonia [92] y Frey (1995:72) dio una longitud máxima de concha de 6 m (20 pies) para el grupo. Sobre el tema del tamaño de las endocéridas, el especialista en nautiloide Rousseau H. Flower escribió:
No todos son grandes, de ninguna manera, pero se han recolectado especímenes de 3,7 m [12 pies] de largo, y los fragmentos de mayor diámetro indican una longitud máxima mucho mayor. No estoy del todo dispuesto a desacreditar un informe sobre un endoceroide encontrado en una cantera cerca de Watertown, Nueva York , que se midió antes de romperlo y se encontró que alcanzaba una longitud de 30 pies [9,1 m]. [93]
Sin embargo, la longitud desenrollada de los amonites más grandes supera con creces la de incluso estos endocéridos gigantes. Parapuzosia seppenradensis , la especie de amonites más grande conocida, tenía una longitud máxima estimada de caparazón desenrollado de alrededor de 18 m (60 pies). También fue posiblemente el más pesado de todos los cefalópodos conocidos, pasados o presentes, con una masa viva estimada de 1.456 kg (3.210 lb), de los cuales el caparazón constituiría 705 kg. [94] En comparación, los endocéridos más grandes pueden haber pesado alrededor de 1000 kg (2200 lb). [7] En términos de masa, estos son los invertebrados más grandes conocidos que jamás hayan existido, [95] aunque quizás todavía estén en segundo lugar después de los cefalópodos vivos más grandes cuando se considera la masa de tejido solo, ya que en las especies con conchas la gran mayoría del tejido vivo está restringido a la cámara del cuerpo , que ocupa sólo una fracción del volumen interno de la cáscara. [8] También podrían ser los animales con conchas más grandes, o al menos más largos, que jamás hayan existido. [8] [nota 5]
El grupo Yezo del Cretácico en Japón produce múltiples taxones de restos fósiles de cefalópodos de gran tamaño. Oegopsid Yezoteuthis giganteus y Haboroteuthis poseidon solo se conocen por los elementos de la mandíbula, cuyo tamaño es cercano al del calamar gigante. Las especies de Nanaimoteuthis , especialmente N. hikidai, serían el vampiromorfo más grande conocido. [96] Allí también se encuentra un fósil de mandíbula superior incompleto y sin nombre que es aproximadamente el doble de grande que el del calamar gigante maduro. [97]
Reclamaciones históricas
Identificaciones erróneas
Fotografía de Kubota de un gran Onykia robusta , que fue identificado erróneamente como el calamar gigante , Architeuthis dux . La fotografía de gran angular exageró el tamaño del calamar [98] de tal manera que parecía increíblemente grande para la especie.
Los tamaños máximos de ciertas especies de cefalópodos, en particular el calamar gigante y el pulpo gigante del Pacífico, a menudo han sido mal informados y exagerados. La literatura sobre el tamaño de los cefalópodos se ha visto aún más confusa por la frecuente atribución errónea de varios especímenes de calamar al género de calamar gigante Architeuthis , a menudo basándose únicamente en su gran tamaño. Sólo en la literatura académica, tales identificaciones erróneas abarcan al menos las familias de oegopsidos Chiroteuthidae , Cranchiidae , Ommastrephidae , Onychoteuthidae y Psychroteuthidae . [99] [nb 6] Esta situación se ve aún más confusa por el uso ocasional del nombre común 'calamar gigante' en referencia a calamares grandes de otros géneros. [100]
Quizás el error de identificación más notable se relaciona con una fotografía tomada algún tiempo antes de 1993 por el buzo H. Kubota frente al sur de Japón . [101] La imagen muestra un individuo grande de Onykia robusta (anteriormente conocido como Moroteuthis robusta ), que parece estar enfermo o moribundo, junto a un buzo en aguas poco profundas. [98] Un vídeo del mismo animal apareció en una película japonesa hecha para televisión. [98] La imagen fue publicada en el libro European Seashells de 1993 por Guido T. Poppe y Yoshihiro Goto, donde fue identificada como Architeuthis dux , el calamar gigante, y se dice que fue tomada en el Atlántico Norte . [102] De ser cierta, esta imagen representaría la primera fotografía conocida de un calamar gigante vivo. En La búsqueda del calamar gigante (1998), Richard Ellis escribió:
Por un momento pensé que alguna oscura fotografía había capturado la imagen más esquiva de la historia natural. Afortunadamente para quienes dedicaron su vida a la búsqueda de Architeuthis , esto fue sólo una aberración, un caso de identidad equivocada. [98]
Pasaría más de una década antes de que Tsunemi Kubodera y Kyoichi Mori tomaran las primeras fotografías reales de un calamar gigante vivo en estado salvaje el 30 de septiembre de 2004 . [103] Posteriormente, Kubodera y su equipo se convirtieron en los primeros en filmar un calamar gigante adulto vivo el 4 de diciembre de 2006, [104] y los primeros en filmar un calamar gigante vivo en su hábitat natural en julio de 2012. [105] Estos hitos fueron precedidos por las primeras imágenes de un calamar gigante vivo ( paralarva ) en 2001, [106] y la primera imagen de un calamar gigante adulto vivo el 15 de enero de 2002. [107] Desde entonces, se han fotografiado y filmado calamares gigantes vivos en varios de ocasiones. [108]
calamar gigante
El "espécimen casi perfecto" que quedó varado vivo en Trinity Bay , Terranova , el 24 de septiembre de 1877. En la década de 1870 se informó de varios calamares gigantes excepcionalmente grandes en Terranova, y estos fueron documentados meticulosamente en una serie de artículos del zoólogo Addison Emery. Verrill . [109]
Están muy extendidos los informes sobre especímenes de calamar gigante ( Architeuthis dux ) que alcanzan o incluso superan los 18 m (59 pies) de longitud total, pero en los últimos tiempos no se ha documentado científicamente ningún animal que se acerque a este tamaño. [110] Esto a pesar de que hay cientos de especímenes disponibles para su estudio ( c. 700 documentados hasta 2015, de los cuales c. 460 medidos de alguna manera [52] ), incluidos numerosos ejemplos recientes, como los 57 especímenes registrados en Japón. aguas durante un período de 15 meses en 2014-2015. [53] Ahora se cree probable que tales longitudes se lograron mediante un gran alargamiento de los dos largos tentáculos de alimentación , de forma análoga al estiramiento de bandas elásticas, o como resultado de métodos de medición inadecuados, como el marcapasos. [57]
Sobre el tema del tamaño máximo frecuentemente citado de 18 metros (o 60 pies), Dery (2013) citó a los expertos en calamares gigantes Steve O'Shea y Clyde Roper :
Si esta cifra [45 pies o 14 m] parece un poco inferior a las afirmaciones brobdingnagianas hechas sobre Architeuthis en la mayoría de las historias de ciencia popular sobre el animal, probablemente se deba a que prácticamente todos los artículos de interés general repiten diligentemente el número mágico de 60 pies.
Steve O'Shea deplora la perpetuación por parte de los medios de comunicación de lo que él cree que es una exageración que pone a prueba la credulidad, basada en la estimación ocular del biólogo del siglo XIX Thomas Kirk sobre la longitud de un espécimen. [nb 7] En un comentario sobre el borrador final de este artículo, O'Shea escribió: "Kirk tomó el ritmo, en sus propias palabras, porque no tenía ninguna regla o medida a mano, y creo que esta tergiversación se ha perpetuado lo suficiente; si eran pie sobre pie, es decir, del talón directamente a la punta, aceptaría 57 (o 58, cualquiera que fuera la cifra exacta), pero creo que perpetuar esto como un hecho no le hace ningún favor a la ciencia".
Roper, en sus comentarios sobre el borrador final de este artículo, fue aún más conservador y escribió: "no hay registros confirmados de calamares gigantes de más de 45 pies [14 m] de longitud total. La mayoría tiene entre 25 y 35 pies [7,6–10,7 m] He examinado especímenes en museos y laboratorios de todo el mundo (quizás unos 100 aproximadamente) y creo que la cifra de 60 pies proviene del miedo, la fantasía y de tirar de los tentáculos altamente elásticos hacia afuera. punto de ruptura cuando se miden en la orilla o en cubierta."
Animales más grandes reportados
Distribución de frecuencia de longitud total, longitud del manto y masa en Architeuthis dux . El valor atípico extremo de 2000 lb (910 kg) proviene de Verrill (1880a) y es poco probable que sea exacto; el siguiente individuo más masivo en el conjunto de datos pesaba sólo 320 kg (700 lb), y el 95% de los especímenes pesaban menos de 550 lb (250 kg). [40]Regresiones lineales log-log para Architeuthis dux : (A) longitud total versus masa; (B) longitud del manto versus longitud total; y (C) longitud del manto versus masa [40]
Paxton (2016a) investigó el tamaño máximo de Architeuthis realizando un análisis estadístico utilizando datos de registros bibliográficos de especímenes de calamar gigante . Seleccionó lo que consideraba los registros de tamaño más grandes para cada longitud del manto (ML), longitud estándar (SL) y longitud total (TL). El estudio de Paxton ha sido criticado por expertos en calamares gigantes, que han puesto en duda la fiabilidad de algunos de los registros bibliográficos seleccionados. [59]
Para la longitud del manto, Paxton (2016a:83) consideró los 3,35 m (11 pies) informados por Dell (1952:98) como el "medido más largo", aunque "de manera más confiable" el espécimen ML de 2,79 m (9 pies 2 pulgadas) de Lyall Bay , Nueva Zelanda , documentado por Kirk (1880:312). [nb 8] Paxton añadió: "Un espécimen de 4,5 m [15 pies] de Mauricio a menudo se cita erróneamente, pero la consulta del artículo principal (Staub, 1993) revela una longitud mal definida que claramente no es ML". El mayor LM medido de un calamar gigante recuperado de un cachalote es los 2,4 m (7,9 pies) informados por Keil (1963:320) (aunque Paxton escribe: "la cuenta es confusa y la cifra de 2,4 m probablemente se refiere a la cabeza y ML combinados") o los 1,98 m (6 pies 6 pulgadas) de un espécimen que había sido tragado entero en las Azores , detallado por Clarke (1955:589) y Clarke (1956:257). El ML "estimado visualmente más largo", según Paxton, es el c. 30 m (100 pies) de un espécimen aparentemente observado en el Atlántico norte frente a Portugal , atribuido a una comunicación personal con T. Lipington. También se proporciona un ML más modesto de 4 m (13 pies), basado en un avistamiento en el Océano Índico procedente del documental de televisión de Lynch (2013).
Para la longitud estándar (excluyendo los tentáculos ), Paxton (2016a:83) citó los 31 pies (9,45 m) del " espécimen de Tres Brazos " documentado por Verrill (1880a:192) como el "medido más largo". Entre los especímenes recuperados de cachalotes, el SL más largo "medido definitivamente" es el de 4,95 m (16 pies 3 pulgadas) informado por Clarke (1956:257) y el SL más largo "estimado visualmente" es el c. 9 m (30 pies) atribuidos a una fotografía de un cachalote con restos de calamar gigante en sus mandíbulas, [111] aunque Paxton admitió que "[n]o está claro cuánto o qué porción del cuerpo se comió". Para los SL "estimados visualmente más largos", supuestos más extremos de c. 175 pies (53 m) y c. 30 m (100 pies) se citan a Starkey (1963) y Ellis (1998a:246), respectivamente (este último es un relato de un testigo presencial de Dennis Braun). Paxton trató estas dos últimas estimaciones de tamaño como SL en lugar de TL porque "los calamares generalmente no dejan sus tentáculos expuestos excepto cuando atrapan a sus presas y este parece ser el caso de Architeuthis ".
En cuanto a la longitud total, Paxton (2016a:83) consideró tres registros como candidatos para la "medida más larga": el espécimen de 19 m (62 pies) de Berzin (1972:199), el espécimen de 55 pies 2 pulgadas (16,81 m) descrito por Kirk (1888) como Architeuthis longimanus , un animal de proporciones extrañas sobre el que se ha comentado mucho [nb 7] , y el "espécimen de cosquillas de dedal " de 55 pies (16,76 m) reportado por Verrill (1880a:191), que a menudo se cita como el calamar gigante más grande jamás registrado. [nb 9] Del último, Paxton escribió: "A veces se cita erróneamente como 17,37 m (57,0 pies), pero la fuente es clara: mide 55 pies de largo". Los dos primeros registros, particularmente el de Berzin, son más cuestionables, como explicó Paxton:
La precisión de los dos TL más largos medidos de 19 y 16,81 m de un espécimen encontrado en el intestino de un cachalote del Océano Índico y del espécimen de Nueva Zelanda en 1887, respectivamente, también debería cuestionarse, pero nuevamente, ciertamente no son imposibles. . El espécimen de Nueva Zelanda (llamado Architeuthis longimanus Kirk, 1888) claramente tiene la proporción más grande de TL a ML jamás conocida en Architeuthis [...] lo que llevó a [O'Shea & Bolstad, 2008] a sugerir que la longitud fue escalonada y /o hubo un extenso estiramiento post-mortem. Sin embargo, una relectura del artículo original sugiere que el espécimen, aunque inicialmente marcado, en realidad fue medido; sin embargo, el TL está en el borde del rango del intervalo de predicción del 99,9% [...] por lo que ciertamente fue un muestra. La afirmación de Berzin (1972) sobre el Océano Índico es sospechosa debido a la redondez de la figura, la falta de medidas detalladas y porque en una fotografía asociada, el manto (cuya longitud no fue dada) no parece muy grande en comparación con los hombres de la imagen. . En consecuencia, la medida, si es exacta, representaría otro animal con tentáculos muy largos. [117]
El tentáculo de 5,8 m (19 pies) que el pescador Theophilus Picot cortó un animal vivo el 26 de octubre de 1873. Picot estimó la longitud total del calamar en 18 m (60 pies). [118]
Sin embargo, como señaló Paxton (2016a:86), el análisis genético de Winkelmann et al. (2013), que concluyó que probablemente existe una única especie de Architeuthis distribuida globalmente , no abarcó estos dos especímenes y, por lo tanto, es posible que exista una segunda especie de calamar gigante, aún sin muestrear, con tentáculos proporcionalmente más largos.
Posteriormente se confirmó que la longitud total de 19 m (62 pies) del espécimen de Berzin era errónea; Según Valentin Yukhov, que participó en el descubrimiento del espécimen, debería haber marcado 9 m (30 pies). [119] El error de imprenta se reprodujo en la traducción al inglés publicada el año siguiente y luego se propagó en varios artículos sobre calamares gigantes. [119] Dado que el espécimen de Berzin no es tan grande como se informó originalmente, el calamar gigante más largo recuperado de un cachalote es el individuo de 34 pies 5 pulgadas (10,49 m) de TL registrado por Clarke (1956:257) (este espécimen también tiene el ML y SL confirmados por más tiempo de cualquier calamar gigante procedente de un cachalote). [120] Paxton consideró que el TL "estimado visualmente más largo" era el de 60 pies (18 m) publicado por Murray (1874:121), a partir de un relato de un testigo ocular del pescador Theophilus Picot, quien afirmó haber golpeado al animal flotante desde su bote. , provocando que ataque . Picot logró cortar uno de sus tentáculos, que posteriormente fue examinado por varios autores. [121]
Quizás el más grande de todos los especímenes de calamar gigante registrados fue el que se encontró flotando en la superficie frente a Saint-Gilles , Reunión , el 4 de marzo de 2016. [119] Aunque debido a su gran tamaño el espécimen no pudo recuperarse en su totalidad, la cabeza y la corona del brazo se salvaron. Fundamentalmente, esto significó que se podía medir el pico para estimar la longitud del manto y la longitud total del espécimen. Utilizando diferentes ecuaciones de escala alométrica , [nb 10] la longitud rostral inferior del pico, de 19,74 mm (0,777 pulgadas), dio una longitud estimada del manto dorsal de 215,3 a 306,0 cm (7,064 a 10,039 pies) y esto, a su vez, fue se utiliza para estimar la longitud total en 11,025 a 15,664 m (36,17 a 51,39 pies). [119]
Afirmaciones más extremas y extravagantes sobre el tamaño del calamar gigante, que pertenecen firmemente al ámbito de la criptozoología , han aparecido en las obras de autores como Bernard Heuvelmans , Willy Ley e Ivan T. Sanderson . [126] La existencia de estos calamares gigantescos a menudo se apoya en referencia a las cicatrices circulares gigantes que a veces se encuentran en los cachalotes , que se supone que fueron infligidas por las ventosas de los calamares gigantes que luchaban. A veces, estas afirmaciones van acompañadas de extrapolaciones del tamaño corporal basadas en la ampliación isométrica de un calamar gigante "típico". [127] Sin embargo, tales cicatrices no son necesariamente de origen de calamar y, en cambio, pueden representar crecimientos de hongos o marcas de mordeduras, siendo las lampreas marinas ( Petromyzon marinus ) una posible fuente. [128] Incluso en el caso de auténticas marcas de chupadores de calamar gigante, es posible que el crecimiento posterior de la piel las haya agrandado mucho más allá de sus dimensiones originales. [129]
Un trozo de piel de cachalote con cicatrices de chupadores de calamar gigantes. En esta imagen ampliamente reproducida, las cicatrices de las ventosas más grandes miden alrededor de una pulgada (2,5 cm) de ancho. [130]
Sin embargo, en la literatura están muy extendidas las afirmaciones de que existen enormes cicatrices en forma de ventosa. Richard Ellis recopiló algunos de "los ejemplos más atroces" en su libro La búsqueda del calamar gigante . [131] Entre ellas se incluye la afirmación de Dozier (1976) de que "un calamar gigante ordinario de 50 pies [15 m] deja marcas de ventosas con dientes que miden entre tres y cuatro pulgadas [7,6 a 10,2 cm] de ancho en una ballena, pero el esperma Se han capturado ballenas con marcas de tentáculos de 46 cm [18 pulgadas] de ancho". El tratamiento monográfico del cachalote de L. Harrison Matthews , publicado en 1938, incluye lo siguiente: "Casi todos los cachalotes machos llevan cicatrices causadas por las ventosas y garras de calamares grandes, cicatrices causadas por ventosas de hasta 10 cm. [3.9 en] de diámetro siendo comunes las marcas de garras toman la forma de rayones de 2 a 3 m [6,6 a 9,8 pies] de largo y parecen ocurrir con más frecuencia que las marcas de ventosas. [132] Ellis (1998a:142) escribió que esta figura de 10 cm es "mucho más grande que cualquier otra dimensión de ventosa registrada que uno sospecha algún tipo de error, ya sea en la medición o en la transcripción".
Wood (1982:192) cubrió el tema con cierto detalle:
Se han conjeturado medidas de 90 pies [27 m], 130 pies [40 m] e incluso 200 pies [61 m] para calamares gigantes a partir del tamaño de las marcas de ventosas encontradas en la piel de los cachalotes capturados, pero es peligroso colocarlos demasiada confianza en esta evidencia. Verrill dice que las ventosas más grandes de los tentáculos de un espécimen de 9,8 m [32 pies] de largo medían 3,2 cm [1 1⁄4 pulgadas ] de diámetro, y las de un espécimen de 16 m [52 pies] aproximadamente 5,1 cm [2 pulgadas]. Daniel (1925), sin embargo, examinó las marcas de ventosas en la cabeza de un cachalote que medía 3,5 pulgadas [ 8,9 cm] de ancho, y se han encontrado otras que medían hasta 5 pulgadas [13 cm] de diámetro en la piel de los espermatozoides. Ballenas capturadas en el Atlántico Norte. Ivan Sanderson (1956) va aún más lejos y afirma que se han encontrado marcas de ventosas de más de 46 cm [18 pulgadas] en las cabezas de los cachalots, ¡pero no explica cómo las pobres ballenas lograron escapar de las garras de tales colosos!
El consenso general es que las marcas de ventosas excepcionalmente grandes, es decir, de más de 5,1 cm de diámetro, son cicatrices antiguas que han aumentado de tamaño a medida que el cachalote crecía.
Uno de los pares de mazas tentaculares de Architeuthis , que muestra las ventosas agrandadas de la manus, que son las más grandes encontradas en cualquiera de las extremidades del calamar gigante.
El biólogo marino Frederick Aldrich , que examinó personalmente más de una docena de especímenes de calamar gigante, escribió que su espécimen más grande de Terranova tenía ventosas tentaculares "de aproximadamente dos pulgadas [5,1 cm] de diámetro", pero que "las chupadoras y su armamento dentado de más de Se han encontrado doce pulgadas [30 cm] de diámetro en el estómago de cachalotes como desechos no digeribles". [134] Esto lo llevó a considerar la idea de un calamar gigante de más de 150 pies (46 m) de largo, [nb 11] e incluso a sugerir un nombre binomial para esta especie de gran tamaño, si alguna vez se descubriera: Architeuthis halpertius . [134]
En comparación, los retoños de calamar gigante normalmente alcanzan un diámetro máximo de sólo unos pocos centímetros. Basándose en un examen detallado de varios especímenes grandes de aguas de Nueva Zelanda, Förch (1998:55) escribió que "[l]as ventosas más grandes [...] en los brazos sésiles son muy constantes entre 21 y 24 milímetros [0,83 –0,94 pulg] de diámetro externo". En el calamar gigante, las ventosas más grandes de todas se encuentran en la porción central de la maza tentacular , llamada manus , y entre los especímenes examinados por Förch (1998:53) éstas alcanzaron un diámetro máximo de 28 a 32 mm (1,10 a 1,26 pulgadas). ). Clarke (1980) escribió: "Aún no he visto pruebas concluyentes que sugieran que las cicatrices de las ventosas midan más de 3,7 centímetros [1,46 pulgadas] de ancho". [138] Según Roper y Boss (1982:97), las ventosas más grandes de las mazas tentaculares alcanzan los 5,2 cm (2,0 pulgadas) de diámetro.
Estimaciones de masa
Una docena de personas intentando mover un calamar gigante que pesa más de 200 kg (440 lb) [139]
Ahora se acepta que el calamar gigante tiene una masa máxima de varios cientos de kilogramos, [140] pero la literatura está plagada de afirmaciones de pesos mucho mayores. Clarke (1966), por ejemplo, estimó la masa de los especímenes de calamar gigante más grandes en alrededor de 1 tonelada (2200 libras). [141] De manera similar, Richard Ellis escribió: "Cuando en realidad se han pesado cadáveres de calamares [gigantes], parece que los más largos, en el rango de 50 pies [15 m], por ejemplo, pesan alrededor de una tonelada [910 kg] ". [142] Se pueden encontrar estimaciones mucho mayores de la masa del calamar gigante, por ejemplo, en Natural History of Marine Animals de MacGinitie & MacGinitie (1949): "se encontraron dos brazos de Architeuthis que medían 42 pies [13 m] de largo, y si se reconstruyó un cuerpo [...] el calamar al que pertenecían estos brazos medía 1,4 m [4,6 pies] de diámetro y 7,3 m [24 pies] de largo, con una medida total de 20 m [66 pies]. alrededor de 42 1 ⁄ 2 toneladas [38,6 toneladas]". Agregaron que un espécimen de 55 pies (17 m), como el reportado en Thimble Tickle , [nb 9] "habría pesado 29 1 ⁄ 4 o 30 toneladas [26,5 o 27,2 toneladas] incluidos los tentáculos, una cifra verdaderamente noble. animal, pesando poco más de una quinta parte del peso de la ballena más grande y mayor que el tiburón ballena y el tiburón peregrino , el mayor de todos los peces". [143] Ellis caracterizó estas estimaciones como "exageraciones infundadas". [142] En la edición revisada de Historia natural de los animales marinos , publicada en 1968, los autores redujeron su estimación a menos de 8 toneladas. [144]
Bernard Heuvelmans creía que "debe haber Architeuthis que pesen más de 5 toneladas, y algunos incluso más grandes que deben pesar entre 2 y 27 toneladas, siendo el peso normal unas 8 toneladas. Hay buenas razones para creer que pueden existir incluso ejemplares dos veces tan largo como el de Thimble Tickle, que, dependiendo de su grosor, podría haber pesado entre 16 y 216 toneladas, pero probablemente alrededor de 64 toneladas." [145] Ellis, que consideró estas estimaciones "completamente ridículas", escribió:
Heuvelmans comete un error fundamental al calcular el peso de algunos de estos monstruos cuando escribe que "la densidad de los seres vivos es sólo ligeramente superior a la del agua... un decímetro de carne viva pesa aproximadamente tanto como un litro de agua. " Esto puede ser cierto para otras criaturas vivientes, pero la carne de Architeuthis , saturada con cloruro de amonio , es más ligera que el agua, y el calamar gigante tiene una flotabilidad neutra . (Se cree que esta es la razón por la que se encuentran calamares muertos o moribundos flotando en la superficie o arrastrados a la playa). Por lo tanto, su suposición de que el calamar Thimble Tickle de 55 pies de largo [17 m] habría "probablemente pesaba cerca de 24 toneladas" es evidentemente erróneo. [146]
Sobre el tema de la masa del espécimen de Thimble Tickle, Wood (1982:190) se refirió al trabajo del zoólogo y escritor soviético Igor Akimushkin :
Según el Dr. Igor Akimushkin (1965), teutólogo ruso, un calamar gigante de 12 m [39 pies] de largo pesará 1 tonelada [2200 lb] si la cabeza, el manto y los brazos combinados constituyen la mitad de la longitud total. Dado que existe una relación cúbica entre las dimensiones lineales de Architeuthis y su volumen o peso, esto significa que el monstruo Thimble Tickle debe haber escalado alrededor de 2,8 toneladas [6.200 libras] (es decir, el peso de un gran hipopótamo macho), aunque 2 toneladas [4.400 libras] lb] es probablemente una cifra más realista.
Pulpo gigante del Pacífico
El tamaño máximo del pulpo gigante del Pacífico ( Enteroctopus dofleini ) ha sido durante mucho tiempo motivo de debate en la comunidad científica, con informes dudosos sobre ejemplares que pesan cientos de kilogramos.
En 1885, al informar sobre el espécimen de pulpo más largo registrado de manera confiable hasta ese momento, el renombrado malacólogo William Healey Dall escribió:
En 1874 atravesé un pulpo en el puerto de Iliuliuk, Unalashka , que luego fue colgado, mediante una cuerda atada alrededor del cuerpo inmediatamente detrás de los brazos, a uno de los pescantes de popa del barco de reconocimiento costero bajo mi mando. Tan pronto como el animal murió y los músculos se relajaron, noté que las puntas de los tentáculos más largos apenas tocaban el agua. Al medir la distancia con una cuerda, encontré que era de dieciséis pies [4,9 m], lo que le daba a la criatura una extensión de punta a punta del par de brazos más largo, de no menos de treinta y dos pies [9,8 m]. Los brazos hacia las puntas eran extremadamente delgados, pero bastante robustos hacia el cuerpo, que medía algo más de 30 cm [un pie] de largo. Los retoños más grandes tenían 6,4 cm [2 pulgadas y media] de diámetro; toda la criatura casi llenaba una gran tina de lavado. Partes de este espécimen se encuentran ahora en el museo nacional de Estados Unidos . [147]
En un artículo para el Servicio Nacional de Pesca Marina que resume los conocimientos sobre el pulpo gigante del Pacífico, High (1976:17-18) escribió:
Se han encontrado y capturado varios pulpos de más de 45 kg [100 libras]. Se han reportado casos mucho más grandes pero, como el monstruo del Lago Ness , generalmente eluden al fotógrafo o científico cuidadoso. La mayoría de los pulpos pesan menos de 32 kg [70 libras] y una longitud estirada de 4,6 m [15 pies] o menos. La longitud total entre los brazos no es una medida adecuada debido a la inusual elasticidad del animal.
A finales de la década de 1950, entrevisté a un buzo comercial canadiense, Jock MacLean de Prince Rupert , Columbia Británica. Informó que había capturado una criatura inmensa que pesaba 272 kg [600 libras] y medía 9,8 m [32 pies] desde la punta del brazo hasta la punta. Lamentablemente, las fotografías de MacLean eran de mala calidad. Ocasionalmente capturaba animales más pequeños, de hasta 181 kg [400 libras], en su actividad comercial de pesca del pulpo.
Hochberg & Fields (1980:436) hizo referencia al mismo espécimen y escribió: "el espécimen más grande registrado con una extensión total de brazo de 9,6 m [31 pies] y un peso de 272 kg [600 lb]". Sin embargo, estas cifras son sólo estimaciones ya que, contrariamente a la cita anterior de High (1976:17-18), parece que este espécimen nunca fue recolectado ni medido. [40] Murray Newman, director del Acuario de Vancouver durante 37 años, citó a Jock MacLean en sus memorias de 1994, Life in a Fishbowl : "El año siguiente [1957] en el mismo lugar, vi uno, tal vez treinta y dos pies [9,8 m] de ancho y seiscientas libras [272 kg]. Sin embargo, no fue por ella, ¡no hay lugar donde guardarla! [148] Sin embargo, la conversión métrica engañosamente precisa de 272 kg (para 600 lb) y la conversión imprecisa de 9,6 m (para 32 pies; empleando ingenuamente un factor de conversión de 0,3 en lugar de 0,3048 ) ganaron amplia aceptación como las dimensiones máximas registradas. del pulpo gigante del Pacífico, y se han repetido mucho. [nota 12]
También se informa que Jock MacLean capturó un animal de 198 kg (437 lb) con una envergadura de brazos de 8,5 m (28 pies) cerca de Port Hardy , Columbia Británica , en marzo de 1956. [150] Otro espécimen gigante fue capturado frente a Santa Bárbara , California , en 1945. Su peso se registró en 182,3 kg (402 lb) y la fotografía superviviente permite estimar su longitud total en más de 3 m (9,8 pies) y su envergadura entre 6 y 6,7 m (20 a 22 pies). ). [81] En un libro dedicado al pulpo gigante del Pacífico, Cosgrove y McDaniel (2009:72) resumieron el conocimiento sobre el tamaño máximo de la especie de la siguiente manera:
El espécimen que William Dall arponeó en 1885 [ sic ] en Iliuliuk tenía la envergadura radial más grande de cualquier pulpo gigante del Pacífico jamás medido. El gigante Port Hardy de 1956 de Jock MacLean fue el más grande jamás pesado. El ejemplar de Santa Bárbara fotografiado en 1945 fue el segundo más pesado. Parecería que pulpos que pesan hasta 272 kg (600 lb) y con envergaduras radiales de más de nueve metros (30 pies) están dentro del ámbito de lo posible, pero en realidad nunca han sido documentados ni midiendo ni pesando.
Posible disminución de tamaño.
No se han reportado especímenes que se acerquen a estos tamaños extremos desde mediados del siglo XX. Esta falta de individuos gigantes es corroborada por los pescadores comerciales de pulpos; Ninguno de los entrevistados por Cosgrove y McDaniel (2009) había capturado un solo animal que pesara más de 57 kg (126 lb) en los 20 años anteriores, entre los muchos miles capturados durante ese período. [82] El especialista en pulpos Roland Anderson, biólogo del Acuario de Seattle durante más de 30 años, había buscado durante mucho tiempo, sin éxito, encontrar un pulpo gigante del Pacífico que pesara más de 45 kg (100 lb). En un intento por criar un espécimen verdaderamente enorme, Anderson alimentó ad libitum a varios machos cautivos . El animal más pesado (apodado 'Grande') alcanzó un peso máximo de 43 kg (95 lb) y sus ventosas más grandes medían 7,9 cm (3,1 pulgadas) de diámetro. [151] Anderson sugirió que la especie ahora podría estar madurando a un tamaño más pequeño como resultado de la bioacumulación de tóxicos , lo que podría explicar la falta de especímenes verdaderamente gigantescos en los últimos tiempos. En particular, se han identificado altas concentraciones de metales pesados y PCB en las glándulas digestivas de los pulpos gigantes salvajes del Pacífico, probablemente procedentes de su presa preferida, el cangrejo rojo de roca ( Cancer productus ). [152] Un estudio preliminar encontró que los animales de acuario alimentados con cantidades iguales de mariscos crudos y C. productus vivo (capturado localmente en Elliott Bay ) maduraron a un tamaño más pequeño, alcanzaron un peso máximo más bajo (27 kg [60 lb] promedio), y tenían concentraciones más altas de la mayoría de los metales pesados que aquellos alimentados únicamente con mariscos crudos (36 kg [79 lb] de media, incluido el espécimen de 43 kg [95 lb] antes mencionado). [153]
Especies más grandes por medida
El tamaño de los cefalópodos se puede cuantificar de varias formas. Algunas de las medidas de tamaño más comunes se tratan a continuación. Las siguientes cuatro tablas enumeran sólo las especies existentes; Los taxones extintos se tratan por separado al final.
La lista de cefalópodos más grandes por longitud del manto está dominada por los calamares, con más de veinte especies superando a los pulpos y sepias de mayor tamaño. El más grande de todos es el calamar colosal ( Mesonychoteuthis hamiltoni ) con una longitud máxima estimada del manto de 3 m (9,8 pies) (Roper & Jereb, 2010c:173). Históricamente se han reportado longitudes de manto aún mayores para el calamar gigante ( Architeuthis dux ), pero estos han sido desacreditados (ver O'Shea y Bolstad, 2008).
Largo total
'Archie', un espécimen de calamar gigante en el Museo de Historia Natural de Londres , mide 8,62 m (28,3 pies) de longitud total (ver Ablett, 2012 para más información sobre este espécimen)Un calamar de aleta grande (cf. Magnapinna ), uno de los cefalópodos más largos conocidos. Este ejemplar fue filmado en octubre de 2000 por DSV Alvin en el Golfo de México , a 1.940 m (6.360 pies) de profundidad.Un espécimen recién capturado de Ommastrephes bartramii en aguas del norte de Hawai . Esta especie crece hasta una longitud total de 2 m (6,6 pies).Los ejemplares grandes de Enteroctopus dofleini pueden superar los 3 m (9,8 pies) de longitud total.
Los ejemplares más largos científicamente documentados pertenecen al calamar gigante , con una longitud total máxima de 14 a 15 m (46 a 49 pies) (Roper & Shea, 2013:114). A pesar de sus tentáculos proporcionalmente más cortos, el calamar colosal puede rivalizar con el calamar gigante en longitud total, pero los límites de tamaño de la especie son inciertos porque sólo se han registrado un puñado de especímenes maduros.
Masa
El calamar colosal de 495 kilogramos (1.091 libras) en exhibición en el Museo de Nueva Zelanda Te Papa Tongarewa . Es el cefalópodo más masivo jamás registrado.El mayor ejemplar documentado de T. danae , conservado en el Museo Nacional de Historia Natural . Originalmente pesaba 161,4 kg (356 lb) y tenía una longitud de manto de 160 cm (5,2 pies).Una hembra madura muy grande de Onykia robsoni con una longitud de manto de 88,5 cm (2,90 pies) y un peso total de 11,1 kg (24 lb).Hembra de Lepidoteuthis grimaldii que mide 61,7 cm (2,02 pies) de largo del manto y pesa 4,07 kg (9,0 lb), de Chatham Rise frente a Nueva Zelanda (obsérvese los prominentes cojines dérmicos , probablemente utilizados para flotar )El peso máximo del pulpo gigante del Pacífico ( Enteroctopus dofleini ) ha sido muy debatido, existiendo numerosos informes de ejemplares que superan los 100 kg (220 lb).Sepia latimanus es la segunda especie de sepia más grande y rivaliza estrechamente con S. apama tanto en longitud como en masa del manto.
El cefalópodo más pesado conocido y el invertebrado vivo más grande es el calamar colosal . El espécimen más grande registrado de esta especie, capturado en el Mar de Ross en 2007, pesaba 495 kg (1091 lb). Sin embargo, su pico no es el más grande conocido de esta especie; Se han recuperado picos de calamar colosales aún más grandes de los estómagos de cachalotes , lo que indica que esta especie puede crecer aún más.
Los nautilos son los únicos cefalópodos existentes con una verdadera capa externa; en otros grupos el caparazón se ha internalizado o se ha perdido por completo. Las conchas internas incluyen las sepias de las sepias, los gladii de los calamares y los calamares vampiro, las conchas aladas de los octópodos cirratos y las conchas espirales de Spirula . Además, las hembras del género de pulpos Argonauta secretan una cáscara de huevo especializada, delgada como el papel, en la que residen, y esto se considera popularmente como una "cáscara", aunque no está adherida al cuerpo del animal (ver Finn, 2013).
El diámetro de la concha de los cefalópodos es de interés tanto para los teutólogos como para los conchólogos . El Registro de conchas de tamaño récord mundial , la publicación más completa sobre el tamaño máximo de concha en moluscos marinos, especifica que los especímenes "deben medirse con calibres tipo vernier y deben reflejar la mayor dimensión mensurable de la concha en cualquier dirección, incluidos los procesos de concha dura". material producido por el animal (es decir, espinas, alas, quillas, canales sifonales, etc.) y sin incluir aditamentos, percebes , algas coralinas o cualquier otro organismo incrustante" (Pisor, 2008:14). A diferencia de la mayoría de las otras medidas del tamaño de los cefalópodos, el diámetro de la concha se puede determinar con un alto grado de precisión y, por lo general, deja poco lugar a la ambigüedad. Por esta razón, generalmente se registra con una precisión de una décima de milímetro (0,0039 pulgadas), como es estándar en conchología .
Cuando el Registro de conchas de tamaño récord mundial cambió de propietario en 2008, se lanzó como una base de datos en línea además de su publicación impresa. Los cambios posteriores en las reglas significaron que todos los registros requerían verificación fotográfica. Con el tiempo, se eliminaron de la base de datos los registros más antiguos de los que no se pudo obtener evidencia fotográfica. Como resultado, algunos registros de ediciones anteriores del registro en realidad exceden el tamaño de los titulares de registros oficiales actuales, a veces por márgenes considerables. Cuando esto ocurre, el tamaño más grande registrado en todas las ediciones se muestra primero y luego se anota cualquier discrepancia o registros en competencia. Cuando un registro bibliográfico confiable supera todos los especímenes incluidos en el registro, se proporciona en su lugar y se anotan los registros del registro a partir de entonces. Pisor (2008) fue la quinta y última edición impresa del registro publicada antes del cambio de regla, y Barbier et al. (Nd) es la base de datos en línea actual y continuamente actualizada. El registro sólo abarca las conchas de nautilos y Spirula y las cáscaras de huevos de Argonauta .
Ojo de calamar gigante preservadoGonatos sp. nadando junto a un sumergible durante una inmersión en el Arco Kermadec frente a Nueva Zelanda . Las gonátidas como ésta tienen ojos proporcionalmente un poco más grandes que los calamares gigantes. [164]
Los calamares gigantes y colosales tienen los ojos más grandes registrados de cualquier animal vivo, con un diámetro máximo de al menos 27 cm (11 pulgadas) y una pupila de 9 cm (3,5 pulgadas) . [5] Esto es tres veces el tamaño de los ojos de pez más grandes (hasta 90 mm (3,5 pulgadas) en el pez espada ) y más del doble del diámetro de los ojos de ballena más grandes (hasta 109 mm (4,3 pulgadas), 61 mm ( 2,4 pulgadas) y 55 mm (2,2 pulgadas) en el cachalote azul , el jorobado y el cachalote , respectivamente, que son los más grandes entre los vertebrados . [5] Un calamar grande y colosal capturado en 2014 y disecado en el Museo de Nueva Zelanda, Te Papa Tongarewa, tenía ojos de entre 35 y 37 cm (14 a 15 pulgadas) de ancho. [165] Hay informes no confirmados del siglo XIX sobre ojos de calamar gigante de hasta 40 cm (1,3 pies) de ancho. [166] Se sabe que sólo los ictiosaurios extintos se acercaron a estas dimensiones, [167] y algunas especies tenían ojos de hasta 35 cm (14 pulgadas) de diámetro. [168]
A pesar de su tamaño, los ojos de los calamares gigantes y colosales no parecen desproporcionadamente grandes; no se desvían significativamente de la relación alométrica observada en otras especies de calamares. [169] Los sepiolidos se caracterizan por tener ojos excepcionalmente grandes, que son mucho más grandes en relación con la longitud de su manto que los del calamar gigante; lo mismo ocurre con las especies de Histioteuthis . [164] Las gonatidas y los loligínidos Loligo y Lolliguncula también tienen ojos proporcionalmente algo más grandes que Architeuthis . [164] Algunas fuentes afirman que el calamar vampiro ( Vampyroteuthis infernalis ) tiene los ojos más grandes de cualquier animal en relación con su tamaño, con un espécimen de 15 cm (5,9 pulgadas) que tiene ojos de alrededor de 2,5 cm (0,98 pulgadas) de diámetro. [170]
Existe cierto debate en la comunidad científica sobre la razón evolutiva detrás de los ojos extremadamente grandes de los calamares gigantes y colosales. [171] Nilsson y otros. (2012) y Nilsson et al. (2013) sostienen que se trata de una adaptación antidepredadores para una mejor detección de cachalotes , en la que los calamares captan la bioluminiscencia del plancton provocada por el movimiento de las ballenas, tal vez desde distancias superiores a 120 m (390 pies). Schmitz y cols. (2013a) y Schmitz et al. (2013b) sostienen que sus ojos son tan grandes debido a un patrón de desarrollo filogenéticamente conservado que gobierna las dimensiones relativas de los calamares y sus ojos, y que cualquier beneficio de aptitud física que su tamaño pueda conferir en términos de evitación de depredadores es el resultado de la exaptación ("pre -adaptación").
Neuronas
Los axones gigantes de los calamares pueden superar 1 mm (0,039 pulgadas) de diámetro: de 100 a 1000 veces el grosor de los axones de los mamíferos . Los axones del calamar de Humboldt ( Dosidicus gigas ) son excepcionales porque pueden alcanzar un diámetro de hasta 1,5 mm (0,059 pulgadas), y los del Loligo forbesii también pueden superar 1 mm. [172] Tal era la importancia del calamar de Humboldt para la investigación de electrofisiología que cuando los animales migraron fuera del alcance de los pescadores chilenos en la década de 1970 "condujeron a la desaparición de un laboratorio de electrofisiología de clase mundial" con sede allí. [173] Los diámetros de los axones gigantes del calamar no necesariamente se correlacionan con el tamaño corporal general; los del calamar gigante ( Architeuthis dux ) tienen sólo 0,137 a 0,21 mm (0,0054 a 0,0083 pulgadas) de espesor. [172]
Diagrama que muestra los tres elementos principales (rojo, verde y amarillo) del sistema neuronal del calamar gigante. Las flechas indican la dirección de transmisión desde el ganglio de la cabeza hacia el manto. El embudo (azul claro) es el lugar de rápida expulsión de agua tras la contracción del manto.
La sinapsis del calamar gigante es la unión química más grande de la naturaleza. Se encuentra en el ganglio estrellado a cada lado de la línea media, en la pared posterior del manto muscular del calamar. La activación de esta sinapsis desencadena una contracción sincrónica de la musculatura del manto, provocando la expulsión contundente de un chorro de agua del manto. Esta propulsión del agua permite al calamar moverse rápidamente a través del agua y, en el caso de los llamados 'calamares voladores', incluso saltar a través de la superficie del agua (rompiendo la barrera aire-agua) para escapar de los depredadores. [174] Muchos elementos esenciales de cómo funcionan todas las sinapsis químicas se descubrieron por primera vez mediante el estudio de la sinapsis del calamar gigante. [175]
fotóforos
Taningia danae , un calamar octopotéutido de gran tamaño , posee fotóforos amarillos "del tamaño de un limón"en las puntas de dos de sus brazos , que son los órganos emisores de luz más grandes conocidos en el reino animal. [176] Imágenes de vídeo filmadas en 2005 en aguas profundas frente a Japón muestran a T. danae emitiendo destellos de luz cegadores desde estos fotóforos mientras ataca a su presa. [177] Un par de párpados musculosos rodean cada fotóforo y es la retirada de estos párpados lo que produce los destellos.Se puede ver claramentea un individuo grande filmado desde un sumergible remoto frente a Hawái en 2015 abriendo las tapas para revelar sus fotóforos. [178] Se cree que este calamar altamente maniobrable utiliza destellos brillantes para desorientar a sus presas potenciales. Los destellos también pueden servir para iluminar a las presas para capturarlas más fácilmente o desempeñar un papel en el cortejo y/o exhibiciones territoriales. [179]
Órganos reproductivos
Un espécimen masculino disecado de Onykia ingens , que muestra un pene erecto de 67 cm (2,20 pies) que se aproxima a la longitud combinada del manto, la cabeza y los brazos.
Se ha observado un alargamiento extremo del pene en el calamar de aguas profundas Onykia ingens . Cuando está erecto, el pene puede ser tan largo como el manto, la cabeza y los brazos combinados. [180] Como tal, los calamares de aguas profundas tienen la mayor longitud de pene conocida en relación con el tamaño corporal de todos los animales móviles, superados en todo el reino animal sólo por ciertos percebes sésiles . [181]
^ El espécimen de cefalópodo científicamente validado más pesado fue un calamar colosal ( Mesonychoteuthis hamiltoni ) que pesaba 495 kg (1091 lb), [1] mientras que las crías de Illex illecebrosus , algunas de las más pequeñas conocidas, tienen una masa de alrededor de 0,00015 g (5,3 × 10 - 6 onzas). [2] La relación de masa entre los dos es . Comparando sólo las hembras adultas, la diferencia entre el calamar colosal y el calamar pigmeo Idiosepius thailandicus , que pesa 0,20 g (0,0071 oz) en su madurez [3] , sería de .
↑ Wood & O'Dor (2000:93) elaboraron esta estimación de masa de la siguiente manera:
No hay [...] pesos publicados de crías de Nautilus spp. El peso de una cría de N. belauensis se estimó utilizando el tamaño de la concha de la cría y un análisis de regresión del diámetro de la concha en cubos versus el peso de siete crías de N. belauensis que pesaban <50 g [1,8 oz] [...] además de un Una sola cría de N. pompilius que fue pesada para el presente estudio el 24 de abril de 1996 en el Acuario de Waikiki . La cría de N. pompilius que pesa 4,33 g [0,153 oz], con un diámetro máximo de concha de 26,25 mm [1,033 in], ajusta una correlación altamente significativa [...] entre el diámetro de la concha al cubo y el peso, lo que indica que una cría de N. belauensis con un diámetro de concha de 30 mm [1,2 pulgadas] [...] pesaría aproximadamente 5,9 g [0,21 oz].
^ Norman y col. (2002:733) escribió: "Los ejemplos más extremos de dimorfismo de tamaño sexual provienen de taxones marinos o parásitos donde las hembras son difíciles de localizar (Ghiselin 1974)".
^ En particular, Scott Cassell mencionó un calamar de Humboldt gigante al que llamó 'Scar' (por las cicatrices en su manto), cuyo tamaño describió así:
Scar mide más de 2,1 m [siete pies] de largo, más grande de lo que nadie ha demostrado jamás. Su cuerpo medía 0,9 m [3 pies] de grosor (es demasiado grande para rodearlo con mis brazos) y estimé que pesaba [ sic ] entre 100 y 110 kg [230 y 250 libras]. [75]
^ Dependiendo de cómo se defina "portador de caparazón", otros contendientes extintos podrían incluir las tortugas acuáticas Archelon y Stupendemys (la primera tiene una estructura esquelética en lugar de un caparazón sólido), la tortuga Megalochelys y el mamífero acorazado Glyptodon y sus parientes.
↑ Iwai (1956:139) informó sobre dos pequeños calamares (92 y 104 mm ML) recuperados del "canal digestivo" de un cachalote , que identificó como pertenecientes al género Architeuthis . Roper y Young (1972:220) demostraron que se trataba ciertamente de una identificación errónea y, en cambio, los atribuyeron a la familia Psychroteuthidae . En un breve resumen de este caso, Ellis (1998a:121) dio una longitud total errónea de "8 pies" (2,4 m) para el mayor de los dos especímenes (aunque en una nota al pie casi idéntica palabra por palabra en Ellis , 1994a:145 afirmó correctamente que la longitud total era "8 pulgadas" [20 cm]). Este error fue repetido por Glaubrecht y Salcedo-Vargas (2004:67), dando lugar a la afirmación de que se trataba de un psicoroteutido inverosímilmente grande "con unos tres metros de longitud total".
^ abBoceto de TW Kirk del espécimen tipo Architeuthis longimanus en aspecto lateral, de Kirk (1888). Tenga en cuenta la longitud extrema de los tentáculos alimentarios en relación con el manto y los brazos.Thomas William Kirk dio el siguiente relato de este espécimen en su descripción formal de Architeuthis longimanus :
[...] A principios del mes pasado, el Sr. Smith, un pescador local, trajo al Museo [Colonial] el pico y la masa bucal de una sepia que había sido encontrada esa mañana en la "Playa Grande" ( Bahía Lyall ), y nos aseguró que la criatura medía sesenta y dos pies [18,9 m] de largo total. Esa tarde me dirigí al lugar e hice un examen cuidadoso, tomé notas, medidas y también obtuve un boceto que, aunque la lluvia terriblemente intensa y el fuerte viento del sur hicieron imposible hacer justicia al tema, confío en que sea posible. transmitirles una idea del perfil general de este pez diablo recién llegado.
Las mediciones mostraron que, aunque el Sr. Smith se excedió al dar la longitud total de 62 pies [18,9 m] (probablemente, al no tener una medida consigo, solo caminó la distancia), esas cifras no estaban tan lejos; porque, aunque el cuerpo era en todos los sentidos más pequeño que el de cualquiera de las especies de Nueva Zelanda descritas hasta entonces, el enorme desarrollo de los muy ligeros brazos tentaculares elevó la longitud total hasta 55 pies y 2 pulgadas [16,81 m], o más de la mitad. nuevamente como la especie más grande registrada hasta ahora en estos mares. La longitud de los brazos tentaculares no es un carácter muy importante, ya que se sabe que son capaces de extenderse o retraerse a voluntad del animal, al menos en una medida considerable.
[...] Este ejemplar era hembra, y a este hecho pueden deberse algunos de los puntos en los que difiere de ocurrencias anteriores; pero, sin embargo, son tan considerables que no tengo ninguna duda de que al menos habrá que crear un nuevo subgénero para su recepción. Mientras tanto lo coloco bajo Architeuthis , con pleno conocimiento, sin embargo, de que no es posible que permanezca allí permanentemente, ya que la forma de los brazos y las aletas por sí solas son suficientes para desvincularlo de ese género. Tan pronto como se presente la oportunidad, espero hacer un estudio más detallado y determinar plenamente sus afinidades. [112]
Kirk (1888:38) proporciona una tabla con un desglose detallado de las diversas medidas del espécimen. Sin embargo, existe una discrepancia entre la longitud total de 684 pulgadas (17,37 m, o exactamente 57 pies) que figura en la tabla, que concuerda con los valores individuales de 71 pulgadas (1,80 m) para el manto, 22 pulgadas (0,56 m) ) para la cabeza y 591 pulgadas (15,01 m) para los tentáculos, y la longitud total de 55 pies 2 pulgadas (16,81 m) proporcionada por Kirk en el cuerpo del artículo.
Wood (1982:191) sugirió que, debido a la naturaleza altamente retráctil de los tentáculos, la longitud total de 62 pies (18,9 m) reportada originalmente por el pescador "puede haber sido correcta en el momento en que encontró el calamar", y que " [e]sto probablemente también explica la discrepancia en las cifras de Kirk". Debido al pequeño tamaño del manto, Wood (1982:191) estimó que "este espécimen probablemente pesaba menos de 300 libras [140 kg]".
O'Shea y Bolstad (2008) opinaron que la longitud total informada de 55 pies 2 pulgadas (16,81 m) "simplemente no puede ser correcta" y la atribuyó a la "imaginación" o al alargamiento artificial de los tentáculos. Agregaron que un calamar gigante hembra con una longitud de manto de 71 pulgadas (180 cm) "medido post mortem y relajado (según los estándares modernos) hoy tendría una longitud total de ≈32 pies [9,8 m]".
↑ ab Kirk (1880:312) escribió sobre este espécimen:
El pasado 23 de mayo, el Ven. El archidiácono Stock, muy amablemente, me envió un mensaje de que tres niños, llamados Edward R. Stock, y Frank y Walter Morrah, habían descubierto esa mañana, en Lyall Bay , lo que tomaron por ser una sepia muy grande, con brazos de varios pies de largo. No perdí tiempo y me dirigí al lugar, decidido, si era posible, a llevarme a casa todo el espécimen; pero juzguen mi sorpresa cuando, al llegar a la bahía, vi un animal del tamaño representado en el dibujo que ahora tienen ante ustedes.* El relato de Víctor Hugo sobre su " pieuvre " me vino vívidamente a la mente, y no pude evitar pensar que un hombre tendría pocas posibilidades si alguna vez estuviera en las garras de un monstruo así.
Mi primer paso después de extender los brazos, fue hacer un boceto aproximado y medidas muy cuidadosas. Luego procedí a extraer el llamado esqueleto , pero descubrí que una o varias personas, que habían visitado el lugar antes que yo, no habían podido resistir la tentación de probar el temple de sus cuchillos en su espalda, y en Como consecuencia cortó el hueso de sepia en varios lugares. Sin embargo, pude no sólo conseguir todas las piezas, sino también el pico , la lengua y algunas ventosas, de las cuales sólo quedaron unas pocas, habiendo sido arrancadas la mayor parte, ya sea en algún encuentro feroz, o durante el mal tiempo que había prevalecido durante algunos días antes. La longitud del cuerpo desde la punta de la cola hasta el margen anterior del manto era de 2,79 m [9 pies 2 pulgadas] y 2,21 m [7 pies 3 pulgadas] de circunferencia; la cabeza desde el margen anterior del manto hasta las raíces de los brazos mide 58 cm [1 pie 11 pulgadas], lo que hace que la longitud total del cuerpo sea 3,38 m [11 pies 1 pulgada]. La cabeza medía 1,2 m [4 pies] de circunferencia. Los brazos sésiles medían 1,30 m [4 pies y 3 pulgadas] de largo y 28 cm [11 pulgadas] de circunferencia; cada uno de estos brazos tenía treinta y seis ventosas, dispuestas en dos filas iguales (como lo muestran las cicatrices) y que medían de una pulgada [ 20,6 a 6,4 mm] de diámetro; cada ventosa estaba reforzada por un anillo óseo armado con de cuarenta a sesenta dientes afilados y curvos. Los brazos tentaculares habían sido arrancados a una longitud de 1,88 m [6 pies y 2 pulgadas], que probablemente era menos de la mitad de su longitud original.
Las aletas eran posteriores, y eran meras expansiones laterales del manto, no se extendían sobre el dorso como en el caso de Onychoteuthis , etc.; cada uno medía 61 cm [24 pulgadas] de largo y 33 cm [13 pulgadas] de ancho.
El hueso de sepia , cuando se extrajo por primera vez, medía 1,91 m [6 pies y 3 pulgadas] de largo y 28 cm [11 pulgadas] de ancho, pero desde entonces se ha reducido considerablemente; era ampliamente lanceolado, con un ápice cónico hueco de 28,6 mm [1 pulgada] de profundidad.
^ ab El espécimen de Thimble Tickle, al que Addison Emery Verrill se refirió como su espécimen número 18, [113] se cita a menudo como el calamar gigante más grande registrado, y durante mucho tiempo ha sido tratado como tal por Guinness . [114]
El calamar Thimble Tickle fue encontrado vivo en alta mar, vivo, el 2 de noviembre de 1878, cerca de la mina de cobre Little Bay, Thimble Tickle , Bahía de Notre Dame , Terranova . Estaba asegurado a un árbol con un rezón y una cuerda y murió cuando la marea bajó. Ninguna parte del animal fue retenida ya que fue cortada para comida para perros. Se decía que la longitud del "cuerpo [...] desde el pico hasta el extremo de la cola" ( es decir, el manto más la cabeza) era de 20 pies (6,1 m), y "uno de los brazos" (presumiblemente un tentáculo) medía 35 pies (10,7 m), para una longitud total de 55 pies (16,8 m). [113] El reverendo Moses Harvey detalló el encuentro en una carta al Boston Traveler fechada el 30 de enero de 1879, que fue reproducida por Verrill (1880a:191-192):
El día 2 del pasado noviembre, Stephen Sherring, un pescador que residía en Thimble Tickle, no lejos de la localidad donde se encontraban los otros peces diablo [espécimen de Verrill No. 19; ver Verrill, 1880a:192], fue arrojado a tierra, estaba en un bote con otros dos hombres; no lejos de la orilla observaron un objeto voluminoso y, suponiendo que pudiera ser parte de un naufragio, remaron hacia él y, para su horror, se encontraron cerca de un pez enorme, de grandes ojos vidriosos, que les hacía desesperarse. esfuerzos por escapar, y batiendo el agua hasta convertirla en espuma con el movimiento de sus inmensos brazos y cola. Estaba encallado y la marea estaba bajando. Del embudo que tenía en la parte posterior de su cabeza expulsaba grandes volúmenes de agua, siendo este su método de retroceder, la fuerza de la corriente, por la reacción del medio circundante, impulsándola en la dirección requerida. A veces el agua del sifón era negra como tinta.
Al encontrar al monstruo parcialmente discapacitado, los pescadores se armaron de valor y se acercaron lo suficiente como para arrojar el rezón de su bote, cuyas aletas afiladas, con puntas de púas, se hundieron en el cuerpo blando. Al rezón le habían atado una cuerda fuerte que habían llevado a la orilla y atada a un árbol, para evitar que los peces se fueran con la marea. Fue una idea feliz, porque el pez diablo se encontró efectivamente amarrado a la orilla. Sus luchas fueron terribles mientras agitaba sus diez brazos en agonía agonizante. Los pescadores tenían cuidado de mantenerse a una distancia respetuosa de los largos tentáculos, que de vez en cuando salían disparados como grandes lenguas de la masa central. Finalmente se agotó y, cuando el agua retrocedió, expiró.
Los pescadores, ¡ay! Sin saber nada mejor, procedió a convertirlo en carne de perro. Era un espécimen espléndido, el más grande capturado hasta ahora, cuyo cuerpo medía 6,1 metros [20 pies] desde el pico hasta el extremo de la cola. Por tanto, era exactamente el doble del tamaño del espécimen de Nueva York [espécimen de Verrill nº 14; ver Verrill, 1880a:189], y cinco pies [1,5 m] más largo que el tomado por [el pescador William] Budgell [No. 19]. No se indica la circunferencia del cuerpo, pero uno de los brazos medía 35 pies [10,7 m]. Esto debe haber sido un tentáculo.
Tal es la fama del espécimen de Thimble Tickle que se ha construido un Centro de Interpretación del Calamar Gigante y una escultura de "tamaño natural" de 55 pies (17 m) cerca del lugar de su captura. [115] La escultura apareció en un sello postal canadiense emitido en 2011. [116]
^Se mide el pico de un calamar gigante. Los picos aislados encontrados en los estómagos de depredadores como los cachalotes pueden usarse para estimar el tamaño del animal original. [122]Romanov et al. (2017) utilizaron las siguientes ecuaciones de escala alométrica para estimar la longitud del manto dorsal (DML) a partir de la longitud rostral inferior (LRL) del pico y la longitud total (TL) a partir de la DML:
(todos los valores en mm) [123]
(todos los valores en m) [124]
(todos los valores en m) [124]
(todos los valores en m) [125]
^ Aldrich reafirmó esta creencia en "Monsters of the Deep", el segundo episodio de la serie de televisión de 1980 Arthur C. Clarke's Mysterious World : "Creo que el calamar gigante alcanza un tamaño máximo aproximado de unos ciento cincuenta pies [46 m ]". [135] Richard Ellis , aparentemente inconsciente de declaraciones similares impresas de Aldrich, comentó: "Es difícil imaginar por qué Aldrich habría hecho una declaración tan irresponsable, a menos que tuviera que ver con estar frente a la cámara". [136] El propio Arthur C. Clarke comentó una vez: "sería ciertamente extraño si el calamar más grande del mundo hubiera estado entre los pocos arrojados a la costa para ser examinados y medidos por naturalistas. Es muy posible que haya especímenes de más de treinta metros [30 longitud mínima." [137]
^ Un ejemplo de la edición de mayo de 2000 de la revista Diver dice: "El récord generalmente aceptado era de proporciones asombrosas, pesaba 272 kg y tenía una envergadura de brazos de 9,6 m". [149]
^ Wood (1982:191) proporcionó los siguientes detalles: "La Dra. Anna M Bidder (com. pers.) del Departamento de Zoología de la Universidad de Cambridge , posee un corte transversal de la pluma de otro Mesonychoteuthis que, a juzgar por su ancho, debe Provienen de un crínquido [ sic ] que mide al menos 5 m [16 pies] de longitud del manto". Bright (1989:146) resume la misma información.
El buque ruso Novoulianovsk , que faenaba en el mar de Ojotsk en 1984, sacó a la luz los restos de un ejemplar gigantesco de Galiteuthis phyllura desde una profundidad de mil a mil trescientos metros (trescientos trescientos a cuatrocientos pies), y Nesis. (1985) dijeron que era "casi tan grande como Mesonychoteuthis hamiltoni (de la misma familia)". Sólo se recogieron un brazo y un tentáculo, pero eran tan grandes (el brazo medía 40 cm de largo [15,6 pulgadas] y el tentáculo 115 cm [45,5 pulgadas]) que Nesis pudo estimar la longitud del manto entre 265 y 275 cm ( 8,69 a 9,02 pies), y la longitud total es de más de 4 metros (más de 13 pies). "Debido a su cuerpo estrecho", escribió Nesis, "concluimos que su masa es consistentemente menor que la de otros calamares grandes".
Roper y Jereb (2010c:165) cuestionaron la validez de este registro y escribieron: "este se considera un registro dudoso que podría referirse a la longitud total; probablemente la longitud máxima del manto sea inferior a 400 a 500 mm". Pero Steve O'Shea comentó:
El fallecido gran Kir Nesis no era alguien que exagerara y debe haber tenido muy buenas razones para citar una posible longitud del manto de 2,7 m [8,9 pies] para Galiteuthis phyllura . Un manto tan monstruoso no sólo convertiría a esta en la 'especie más grande de Galiteuthis ' , sino que también le daría a la especie un manto considerablemente más largo que el de Architeuthis , y marginalmente más largo (0,2 m [0,66 pies]) que el calamar colosal submaduro. ( Mesonychoteuthis ) informamos el año pasado [en 2003]. Lo que me sorprende es que el adulto nunca haya aparecido en la prensa (que yo sepa). Incluso si Galiteuthis carece de ganchos en los brazos, sería un calamar muy aterrador con el que toparse en el abismo. (O'Shea, 2004b)
↑ Este registro se basa en una concha recolectada en 2001 en el mar de Timor frente a Indonesia , que se vendió en 2003 como un espécimen de N. repertus con un diámetro de 268 mm (10,6 pulgadas). Posteriormente se descubrió que medía sólo alrededor de 243 mm (9,6 pulgadas) y la discrepancia se atribuyó a un error de codificación ([Anónimo], 2003a). Se afirmó que otro espécimen de la misma localidad, vendido aproximadamente al mismo tiempo, medía 255 mm (10,0 pulgadas) ([Anónimo], 2003b).
↑ Hay dos lugares que llevan o han llevado el nombre de Joachimsthal: Jáchymov , República Checa y Joachimsthal, Brandeburgo , Alemania. La cita cita un informe inédito y no está claro a cuál se refiere. Si la información es posterior al año 1945, cuando se cambió el nombre oficial de la ciudad checa, lo más probable es que se trate de un lugar en Alemania.
^ Ellis, 1998a:106; Glaubrecht y Salcedo-Vargas, 2004:62
^ Roper y Voss, 1983:55
^ ver Arkhipkin et al., 2015
^ a b C O'Shea y Bolstad, 2008
^ Roper y joven, 1972:205
^ Clarke, 1996:1105; O'Shea y Bolstad, 2008
^ ver Clarke, 1962; Wolff, 1981; 1984; Gröger et al., 2000; Staudinger et al., 2009; Potier et al., 2011
^ Clarke, 1986:11
^ Zeidberg, 2004:4195; Phillips, 2004:ii
^ Nabhitabhata, 1998:32
^ O'Dor y otros, 1986:59; Wood y O'Dor, 2000:93
^ Cosgrove y McDaniel, 2009:88
^ Grulke, 2014:105
^ Rowlett, Joe (6 de octubre de 2017). "Conoce al calamar más pequeño y extraño del mundo, Idiosepius". Arrecifes.com . Consultado el 19 de enero de 2019 .
^ Boletzky, 2003:19
^ Boletzky, 1999:24; 2003:20; Norman et al., 2002:733
^ Norman y otros, 2002:733; Fairbairn, 2007:3
^ Norman y otros, 2002:733
^ ver Kennedy y Cobban, 1990
^ Nishiguchi, MK, R. Mapes (2008). "Cefalópodos" (PDF) . Prensa de la Universidad de California . págs. 163-199.{{cite web}}: CS1 maint: multiple names: authors list (link)
^ Rosa y Seibel, 2010; McClain y otros, 2015; Rosa et al., 2017
^ abcdefg McClain y otros, 2015
^ Collins y otros, 2001; Collins y Villanueva, 2006:294
^ Voss, 1988; Vecchione et al., 2008
^ ver Teichert y Kummel, 1960; Stevens, 1988; Eyden, 2003b; Larson, 2010; Iba y otros, 2015
^ O'Shea y Bolstad, 2008; [Anónimo], Dakota del Norte
^ Nigmatullin y otros, 2001; Roper y otros, 2010b:301
^ Wormuth, 1976:38; normando, 2000:165; Glaubrecht y Salcedo-Vargas, 2004:54
^ abc Roper y otros, 2010b: 301
^ Clarke, 1966:117; Glaubrecht y Salcedo-Vargas, 2004:59
^ Glaubrecht y Salcedo-Vargas, 2004:59
^ Nigmatullin y otros, 2001:10; Roper y otros, 2010b:301
^ ab Cassell, 2005
^ ab Lynnes y Rodhouse, 2002:1087; ver también Carrington, 2000
^ Bolstad, 2008:171
^ Guerra y Segonzac, 2014:65
^ Cosgrove, 1987
^ normando, 2000:214; Norman et al., 2014:124
^ ab Cosgrove y McDaniel, 2009: 67–69
^ ab Cosgrove y McDaniel, 2009:71
^ Anderson, 2003:3; Cosgrove y McDaniel, 2009:71; Yong, 2015
^ O'Shea, 2002:1; 2004a:9; finlandés, 2014a:227
^ O'Shea, 2004a:9
^ Landois, 1895: 108
^ Carnal, 2017
^ Kennedy y Kaplan, 1995:21
^ Landois, 1895: 100
^ Cerveza, 2015
^ [Anónimo], Dakota del Norte; [Anónimo], 2008
^ Teichert y Kummel, 1960:2
^ Flor, 1955:329
^ Landois, 1898:27
^ Grulke, 2014:124
^ ab Tanabe, Kazushige; Misaki, Akihiro; Ubukata, Takao (2014). "Registro del Cretácico tardío de grandes coleoides de cuerpo blando basado en restos de la mandíbula inferior de Hokkaido, Japón". Acta Paleontológica Polonica . doi : 10.4202/app.00052.2013 . ISSN 0567-7920.
^ ab Tanabe, Kazushige; Misaki, Akihiro (20 de junio de 2023). "Registro del Cretácico superior de mandíbulas coleoides no belemnítidas de Hokkaido, Japón, y sus implicaciones evolutivas". Investigación del Cretácico . 151 : 105624. Código bibliográfico : 2023CrRes.15105624T. doi :10.1016/j.cretres.2023.105624. ISSN 0195-6671. S2CID 259228292.
^ abcd Ellis, 1998a: 211
^ ver Ellis, 1998a; Salcedo Vargas, 1999; Glaubrecht y Salcedo-Vargas, 2004
^ Robson, 1933:681; véase, por ejemplo, Mitsukuri & Ikeda, 1895; Manso y Goddard, 1926; Clarke y Robson, 1929; Phillips, 1933; Croker, 1934; Rees, 1950; Smith, 1963; Nesis, 1970
^ ver Wood, 1982:189; Carwardine, 1995:240; Glenday, 2014:62
^ Chupetón, 2009
^ Chupetón, 2010; [Anónimo], Dakota del Norte
^ Paxton, 2016a: 86
^ Murray, 1874: 121
^ abcd Romanov y otros, 2017
^ Paxton, 2016a: 83
^ ver Murray, 1874; Verril, 1875a; Verril, 1875b
^ ver Roeleveld, 2000; Cherel, 2003; Romanov y otros, 2017
^ Roeleveld, 2000:185
^ ab Paxton, 2016a: 85
^ McClain y otros, 2015: Tabla 2
^ ver Sanderson, 1956; Heuvelmans, 1958; Ley, 1959
^ Roper y jefe, 1982:97
^ Madera, 1982:193
^ Roper y jefe, 1982:99; Madera, 1982:192; Haszprunar y Wanninger, 2012:R510
^ Ellis, 1998a: 143
^ ver Ellis, 1998a: 142
^ Mateos, 1938
^ Ley, 1959:210
^ ab Aldrich, 1980:59
^ Ellis, 1998a: 6–7
^ Ellis, 1998a: 7
^ Ellis, 1998a: 6
^ Ellis, 1998a: 142
^ ver Feldman, 1999 para más información sobre este espécimen
^ O'Shea, 2003a; Roper y Jereb, 2010a:121
^ Alejandro, 1998:1233
^ ab Ellis, 1998a: 106
^ MacGinitie y MacGinitie, 1949
^ MacGinitie y MacGinitie, 1968; Madera, 1982:190
^ Heuvelmans, 1958
^ Ellis, 1998a: 107
^ Dall, 1885:432
^ Newman, 1994:66
^ Bavendam, 2000:63–64
^ Newman, 1994:66; Cosgrove y McDaniel, 2009:66–67
^ Anderson, 2003:2; Cosgrove y McDaniel, 2009:71
^ Anderson, 2003:3; Cosgrove y McDaniel, 2009:71; Scheel y Anderson, 2012
^ Anderson, 2003: 2
^ El Loligo forbesi de las Azores (Cephalopoda: Loliginidae) en cautiverio: transporte, manipulación, mantenimiento, etiquetado y supervivencia.
^ El tamaño del cuerpo y la longitud de las aletas como determinantes en la distribución geográfica de los calamares loligínidos.
^ Ifrim, Cristina; Stinnesbeck, Wolfgang; González, Arturo H. González; Schorndorf, Nils; Gale, Andrew S. (10 de noviembre de 2021). "Ontogenia, evolución y distribución paleogeográfica del amonita más grande del mundo, Parapuzosia (P.) seppenradensis (Landois, 1895)". MÁS UNO . 16 (11): e0258510. Código Bib : 2021PLoSO..1658510I. doi : 10.1371/journal.pone.0258510 . ISSN 1932-6203. PMC 8580234 . PMID 34758037.
^ abc Fuchs, Dirk; Stinnesbeck, Wolfgang (1 de noviembre de 2021). "Octobraquios (cefalópodos coleoideos) de gran tamaño con gladius en el plattenkalk turoniano de Vallecillo, México". Investigación del Cretácico . 127 : 104949. Código bibliográfico : 2021CrRes.12704949F. doi :10.1016/j.cretres.2021.104949. ISSN 0195-6671.
^ Kröger, Björn (27 de marzo de 2013). "Los cefalópodos de Boda Limestone, Ordovícico tardío, de Dalarna, Suecia". Revista europea de taxonomía (41). doi : 10.5852/ejt.2013.41 . ISSN 2118-9773.
^ Kroger, Björn; Ebbestad, Jan Ove R. (2014). "Paleoecología y paleogeografía de cefalópodos del Ordovícico tardío (Katian-Hirnantian) de Boda Limestone, distrito de Siljan, Suecia". Lethaia . 47 (1): 15–30. Código Bib : 2014 Letha..47...15K. doi :10.1111/let.12034. ISSN 0024-1164.
^ Fuchs, Dirk; Iba, Yasuhiro; Hola, Alejandro; Iijima, Masaya; Klug, cristiano; Larson, Neal L.; Schweigert, Günter (2020). Brayard, Arnaud (ed.). "Los Muensterelloidea: filogenia y evolución del carácter de los octópodos del tallo mesozoico". Artículos de Paleontología . 6 (1): 31–92. Código Bib : 2020PPal....6...31F. doi :10.1002/spp2.1254. ISSN 2056-2802. S2CID 198256507.
^ Fuchs, Dirk (8 de octubre de 2019). "Eromangateuthis n. Gen., Un nuevo género de un octobraquio gigante con gladius del Albiano tardío (Cephalopoda: Coleoidea)". Aportes paleontológicos . doi : 10.17161/1808.29619 . ISSN 1946-0279.
^ Tanabe, Kazushige; Hikida, Yoshinori (2010). "Fauces de una nueva especie de Nanaimoteuthis (Coleoidea: Vampyromorphida) del Tuironian de Hokkaido, Japón". Investigación Paleontológica . 14 (2): 145-150. doi :10.2517/1342-8144-14.2.145. ISSN 1342-8144. S2CID 128429338.
^ abc Schmitz y otros, 2013a
^ Farquhar, 2014; [Te papá], 2014
^ Tierra y Nilsson, 2012:86
^ Motani y otros, 1999; Humphries y Ruxton, 2002
^ Nilsson y otros, 2012:687
^ Schmitz y otros, 2013a:45
^ Ellis, 1996:177; [Anónimo], 2005a; aunque ver Young et al., 2015
^ Perdiz, 2012
^ ab Adelman y Gilbert, 1990:102
^ Cully, 2008
^ Arata, 1954; Murata, 1988; Maciá et al., 2004; Muramatsu et al., 2013; O'Dor et al., 2013
^ ver Llinás, 1999
^ Ellis, 1998a:149; Barrat, 2015
^ ver Kubodera et al., 2006
^ ver Barrat, 2015
^ Kubodera y otros, 2006:1033
^ Arkhipkin y Laptikhovsky, 2010:299; Caminante, 2010
[Anónimo] (1934). Un calamar gigante. The Scotsman , 27 de enero de 1934. p. 15.
[Anónimo] (2000). Criatura gigante de las profundidades marinas sorprende a los científicos españoles. Reuters, 3 de noviembre de 2000. [Archivado desde el original el 1 de diciembre de 2000.]
[Anónimo] (2002a).巨大イカ現れる!! Sitio web de la prefectura de Kioto, 1 de febrero de 2002. [Archivado desde el original el 12 de junio de 2002] (en japonés)
[Anónimo] (2002b). Sintonizando con un monstruo de las profundidades marinas. CNN , 13 de junio de 2002. [Archivado desde el original el 13 de julio de 2005.]
[Anónimo] (2003a). Nautilus repertus ID:118764. Enciclopedia Shell, Conchology, Inc.
[Anónimo] (2003b). Nautilus repertus ID:118763. Enciclopedia Shell, Conchology, Inc.
[Anónimo] (2003c). Espirula espirula ID:126690. Enciclopedia Shell, Conchology, Inc.
[Anónimo] (2003d). Calamar gigante 'ataca a un barco francés'. BBC News , 15 de enero de 2003.
[Anónimo] (2003e). Una masa gigante desconcierta a los científicos marinos. BBC News , 2 de julio de 2003.
[Anónimo] (2003 y siguientes). La mancha chilena podría ser pulpo. BBC News , 3 de julio de 2003.
[Anónimo] (2005a). La mayor proporción ojo-cuerpo. Récords mundiales Guinness, 19 de enero de 2005.
[Anónimo] (2005b). Calamar gigante muy raro capturado vivo. Boletín informativo de Georgia del Sur , junio de 2005. [Archivado desde el original el 16 de febrero de 2007.]
[Anónimo] (2006). Espirula espirula ID:306458. Enciclopedia Shell, Conchology, Inc.
[Anónimo] (2007). Calamar gigante captado en vídeo por científicos japoneses. Reuters, 21 de enero de 2007.
[Anónimo] (2008). La amonita más grande del mundo. Página 4. Foros en línea de la revista Octopus News.
[Anónimo] (2010). El Proyecto Undersea Voyager - Junta Directiva. El proyecto Voyager submarino. [Archivado desde el original el 24 de diciembre de 2011.]
[Anónimo] (2013). ¡El calamar gigante, captado por la cámara en su hábitat natural por primera vez! NHK, 9 de enero de 2013. [Archivado desde el original el 23 de enero de 2013.]
[Anónimo] (2015a). Amonitas - Moutoniceras moutonianum y Gassendiceras quelquejeui - 148 cm. Catawiki , 20 de septiembre de 2015.
[Anónimo] (2015b). Calamar de dos metros capturado en la costa de Kona. KHON2 , 1 de octubre de 2015.
[Anónimo] (2016). Un monumento en Nueva York rinde homenaje a las víctimas del ataque de un pulpo gigante que nunca ocurrió. El guardián , 1 de octubre de 2016.
[Anónimo] ( nd ). Amonitas [ enlace muerto ] . Minerales y réplicas de fósiles de Hanman. [Archivado desde el original el 10 de febrero de 2003.]
[Anónimo] ( nd ). Calamar de Humboldt, Calamar Gigante, Calamar Gigante de Humboldt, Calamar Gigante, Rojo Diablo (Dosidicus gigas). MexFish.com.
[Anónimo] ( nd ). Capitán Scott Cassell. Explora el verde. [Archivado desde el original el 27 de abril de 2015.]
[Anónimo] ( nd ). ¿Qué tamaño tiene el colosal calamar? Museo de Nueva Zelanda Te Papa Tongarewa.
[Anónimo] ( nd ). El cuerpo del colosal calamar. Museo de Nueva Zelanda Te Papa Tongarewa.
[Anónimo] ( nd ). El pico del calamar colosal. Museo de Nueva Zelanda Te Papa Tongarewa.
[Anónimo] ( nd ). Los brazos y tentáculos. Museo de Nueva Zelanda Te Papa Tongarewa.
[Anónimo] ( nd ). El calamar gigante. Sellos postales canadienses.
Ablett, J. (2012). El calamar gigante, Architeuthis dux Steenstrup, 1857 (Mollusca: Cephalopoda): la creación de un espécimen icónico. Noticias NatSCA no. 23: 16-20.
Adelman, WJ y DL Gilbert (1990). Electrofisiología y Biofísica del Axón Gigante del Calamar. En: DL Gilbert, WJ Adelman y JM Arnold (eds.) Calamares como animales experimentales. Plenum Press, Nueva York. págs. 93-132. ISBN 0306435136 .
Akimushkin, II (1963). Головоногие моллюски морей СССР. [ Golovonogie Mollyuski Morei SSSR .] Academia de Ciencias de la URSS, Instituto de Oceanología, Moscú. 235 págs. (en ruso) [Traducción al inglés de A. Mercado: Akimushkin, II (1965). Cefalópodos de los mares del Programa de Traducciones Científicas de Israel de la URSS, Jerusalén. 223 págs.]
Aldrich, FA (1980). La fuente del obispo marino. [páginas. 55–64] En: KS Goldstein & NV Rosenburg (eds.) Estudios de folclore en honor a Herbert Halpert: A Festschrift. Memorial University de Terranova, St. John's. ix+395 págs. ISBN 0-88901-019-6 .
Aldrich, FA (1991). Algunos aspectos de la sistemática y biología de los calamares del género Architeuthis basados en un estudio de especímenes de aguas de Terranova. Boletín de Ciencias Marinas 49 (1–2): 457–481.
Alejandro, RM (1998). Gigantes de todos los tiempos: los animales más grandes y sus problemas. Paleontología 41 (6): 1231-1245.
Anderson, RC (1996a). Encuentro de pulpos en Slant Rock. De mar y costa 18 (4): 203–204.
Anderson, RC (1996b). ¿Qué come un pulpo gigante del Pacífico? De mar y costa 18 (4): 221–222.
Anderson, RC (2003). Un informe preliminar sobre la bioacumulación en pulpos (Enteroctopus dofleini). En: TW Droscher y DA Fraser (eds.) Actas de la Conferencia de investigación de la Cuenca de Georgia/Puget Sound de 2003. Asociación de Puget Sound, Olympia. 5 págs.
Anderson, RC y RL Shimek (2014). Los pulpos se alimentan de aves. Boletín malacológico americano 32 (2): 220–222. doi :10.4003/006.032.0213
Anderton, J. (2007). El calamar más grande del mundo aterrizó en Nueva Zelanda. El sitio web oficial del gobierno de Nueva Zelanda. [Archivado desde el original el 2 de mayo de 2008.]
Arata, GF (1954). Una nota sobre el comportamiento de vuelo de ciertos calamares. El Nautilus 68 (1): 1–3.
Arkhipkin, AI y VV Laptikhovsky (2010). Observación del alargamiento del pene en Onykia ingens : implicaciones para la transferencia de espermatóforos en calamares de aguas profundas. Revista de estudios de moluscos 76 (3): 299–300. doi :10.1093/mollus/eyq019
Arkhipkin, A., R. Weis, N. Mariotti y Z. Shcherbich (2015). Cefalópodos de 'cola'. Revista de estudios de moluscos 81 (3): 345–355. doi :10.1093/mollus/eyu094
Baird, G. (2002). Persiguiendo gigantes: tras la pista del calamar gigante. NIWA Science, 28 de febrero de 2002. [Archivado desde el original el 19 de enero de 2008.]
Balini, M., JF Jenks, R. Martin, CA McRoberts, MJ Orchard y NJ Silberling (2015). La sucesión de límites Carniano/Noriano en el Parque Estatal Berlín-Ictiosaurio (Triásico Superior, Nevada central, EE.UU.). Paläontologische Zeitschrift 89 (3): 399–433. doi :10.1007/s12542-014-0244-2
Barbier, JP, P. Quiquandon y O. Santini ( ND ). Registro de proyectiles de tamaño récord mundial. Pasión de conchas. [Actualizado continuamente; base de datos consultada el 25 de marzo de 2020.]
Barrat, J. (2015). El raro calamar T. danae capturado en un nuevo video. Noticias científicas del Smithsonian, 9 de diciembre de 2015.
Barrère, F. (2017). Une espèce animale à l'épreuve de l'image: Essai sur le calmar géant. Nouvelle édition revue et augmentée. Édiciones L'Harmattan, París. 300 págs. ISBN 2-14-004283-2 . (en francés)
Bavendam, F. (2000). Luchando con el pez diablo. Buzo , mayo de 2000. págs. 62–67.
Beale, T. (1839). La historia natural del cachalote. A lo que se añade un boceto de un viaje ballenero por los mares del Sur; abarcando una descripción de la extensión, así como de las aventuras y accidentes ocurridos durante el viaje en el que se vio involucrado personalmente el autor. John Van Voorst, Londres. xii + 393 págs. doi :10.5962/bhl.title.110064
Cerveza, M. (2015). Koloss aus der Kreidezeit: Der größte Ammonit der Welt wurde vor 120 Jahren in Seppenrade entdeckt. Westfälische Nachrichten , 22 de febrero de 2015. (en alemán)
Bert, D., S. Bersac, J. Juárez-Ruiz y Z. Hughes (2017). Reducción de tamaño y oscilación ornamental dentro de un linaje barremiano de amonites heteromórficos gigantes (Cretácico temprano, margen noroeste de Tethyan). Cretaceous Research , publicado en línea el 3 de febrero de 2017. doi :10.1016/j.cretres.2017.02.001
Berzin, AA (1971). Кашалот . [ Kashalot .] Instituto de Investigaciones Científicas de Pesca y Oceanografía del Pacífico, Moscú. 368 págs. (en ruso) [traducción al inglés de E. Hoz y Z. Blake: Berzin, AA (1972). El cachalote. Programa de Traducción Científica de Israel, Jerusalén. v+394 págs. ISBN 0-7065-1262-6 .]
Bille, MA (1995). ¿Réquiem por el pulpo gigante? Zoología exótica: revista bimestral de criptozoología 2 (4): 3–4.
Biró-Bagóczky, L. (1976). Titanitas chilensis n. sp. en la Formación Lo Valdés, Titoniano–Neocominao, Provincia de Santiago, Chile. [páginas. L11–L19] En: [Departamento de Geología, Universidad de Chile] Actas Primer Congreso Geológico Chileno: 2–7 Agosto de 1976, Santiago, Chile. Tomo III. Universidad de Chile, Santiago. (en español)
Bittel, J. (2016). Qué tan grande es grande y otros misterios sobre el glorioso calamar gigante de sangre azul. Red de noticias Earth Touch , 20 de junio de 2016.
Boletzky, Sv (1987). Sobre las dimensiones de la cápsula de huevo en Loligo forbesi (Mollusca: Cephalopoda): una nota. Vie et Milieu 37 (3–4): 187–192.
Boletzky, Sv (1997). Limitaciones del desarrollo y heterocronía: una nueva mirada al tamaño de la descendencia en moluscos cefalópodos. Geobios 30 (suplemento 2): 267–275. doi :10.1016/S0016-6995(97)80102-7
Boletzky, Sv (1999). Cefalópodos enanos: condiciones de reproducción en tamaño pequeño. Berichte der Geologischen Bundesanstalt 46 : 24.
Boletzky, Sv (2002). ¿Qué tan pequeño es "muy pequeño" en los cefalópodos coleoides? Berliner Paläobiologische Abhandlungen 1 : 14-15.
Boletzky, Sv (2003). Un límite inferior para el tamaño adulto en cefalópodos coleoides: elementos de una discusión. Berliner Paläobiologische Abhandlungen 3 : 19–28.
Bolstad, KSR (2003). Cefalópodos de aguas profundas: introducción y descripción general. La revista Octopus News en línea.
Bolstad, KSR (2008). Sistemática de Onychoteuthidae Gray, 1847 (Cephalopoda: Oegopsida). Doctor. tesis, Universidad Tecnológica de Auckland, Auckland.
Brillante, M. (1989). Hay Gigantes en el Mar: Monstruos y Misterios de las Profundidades Explorados . Libros Robson, Londres. 224 págs. ISBN 0-86051-481-1 .
Amplio, WJ (2004). ¿Ogro? ¿Pulpo? Los blobólogos resuelven un antiguo misterio. Los New York Times , 27 de julio de 2004.
Buel, JW (1887). Horribles monstruos de las profundidades. [páginas. 68–84] En: Mar y Tierra. Una historia ilustrada de las cosas maravillosas y curiosas de la naturaleza que existieron antes y después del diluvio. Compañía editorial histórica, Filadelfia. 800 págs.
Bullen, pies (1898). [1906]. Lo cual se trata del Kraken. [páginas. 139-148] En: El crucero del Cachalot: la vuelta al mundo detrás de los cachalotes. D. Appleton and Company, Nueva York. xx + 379 págs. doi :10.5962/bhl.title.35756
Caillois, R. (1973). La pieuvre: ensayo sobre la lógica del imaginario. La Table Ronde, París. 231 págs. (en francés)
Carnal, M. (2017). En busca de los antiguos: los cefalópodos gigantes lovecraftianos y el registro fósil. El guardián , 31 de octubre de 2017.
Carr, SM, HD Marshall, KA Johnstone, LM Pynn y GB Stenson (2002). Cómo distinguir un monstruo marino: discriminación molecular de grandes animales marinos del Atlántico Norte. El Boletín Biológico 202 (1): 1–5. doi :10.2307/1543217
Carrington, D. (2000). El calamar grande bate récord. BBC News , 3 de julio de 2000.
Carwardine, M. (1995). El Libro Guinness de los Récords de Animales . Editorial Guinness, Enfield. 256 págs. ISBN 0-85112-658-8 .
Cassell, S. (2005). Bailando con demonios: el calamar gigante de Humboldt. DeeperBlue, 15 de diciembre de 2005.
Cassell, S. (2007). Monster Quest: El calamar gigante encontrado / Blog de Scott Cassell. ScubaBoard, 20 de noviembre de 2007.
Cerullo, MM y CFE Roper (2012). Calamar gigante: en busca de un monstruo marino. Prensa Capstone, North Mankato. 48 págs. ISBN 1429680237 .
Chotiyaputta, C., T. Okutani y S. Chaitiamvong (1991). Una nueva sepia pigmea del Golfo de Tailandia Idiosepius thailandicus n. sp. (Cephalopoda: Idiosepiidae). Venus, Revista japonesa de malacología 50 (3): 165–174. NAID 40000387379
Clarke, A., MR Clarke , LJ Holmes y TD Waters (1985). Valores caloríficos y análisis elemental de once especies de calamares oceánicos (Mollusca: Cephalopoda). Revista de la Asociación de Biología Marina del Reino Unido 65 (4): 983–986. doi :10.1017/S0025315400019457
Clarke, JM (1897). Los cefalópodos del Silúrico inferior de Minnesota. En: EO Ulrich, JM Clarke, WH Scofield y NH Winchell La geología de Minnesota. vol. III, Parte II, del informe final. Paleontología. Harrison y Smith, Minneapolis. págs. 761–812.
Clarke, señor (1962). La identificación de los "picos" de cefalópodos y la relación entre el tamaño del pico y el peso corporal total. Boletín del Museo Británico (Historia Natural), Zoología 8 (10): 419–480.
Clarke, señor (1966). Una revisión de la sistemática y ecología de los calamares oceánicos. Avances en biología marina 4 : 91–300. doi :10.1016/S0065-2881(08)60314-4
Clarke, señor (1980). Cefalópodos en la dieta de los cachalotes del hemisferio sur y su influencia en la biología del cachalote. Informes de descubrimiento 37 : 1–324.
Clarke, señor (1986). Manual para la identificación de picos de cefalópodos . Prensa de la Universidad de Oxford, Oxford. xiii+273 págs. ISBN 0-19-857603-X .
Clarke, MR y N. Goodall (1994). Cefalópodos en las dietas de tres especies de cetáceos odontocetos varados en Tierra del Fuego, Globicephala melaena (Traill, 1809), Hyperoodon planifrons Flower, 1882 y Cephalorhynchus commersonii (Lacepede, 1804). Ciencia Antártica 6 (2): 149–154. doi :10.1017/S0954102094000234
Clarke, R. (1955). Un calamar gigante tragado por un cachalote. Norsk Hvalfangst-Tidende (La Gaceta Ballenera de Noruega) 44 (10): 589–593.
Clarke, R. (1956). Cachalotes de las Azores. Informe de descubrimiento 28 : 237–298.
Clarke, WJ y GC Robson (1929). Notas sobre el varamiento de calamares gigantes en la costa noreste de Inglaterra [ enlace muerto ] . Actas de la Sociedad Malacológica 18 (4): 154–158.
Collins, MA, M. O'Dea y C. Henriques (2001). Un gran Cirroteuthis magna (Cephalopoda: Cirroctopoda) capturado en la Terraza de Cabo Verde (Atlántico Norte). Revista de la Asociación de Biología Marina del Reino Unido 81 (2): 357–358. doi :10.1017/S0025315401003915
Collins, MA y R. Villanueva (2006). Taxonomía, ecología y comportamiento de los octópodos cirrados. [páginas. 277–322] En: RN Gibson, RJA Atkinson y JDM Gordon (eds.) Oceanografía y biología marina: una revisión anual, volumen 44. CRC Press, Boca Raton. 536 págs. ISBN 1-4200-0639-8 .
Colwell, M. (2015). Nuestra profunda necesidad de monstruos que acechan en la oscuridad. BBC News , 23 de junio de 2015.
Cosgrove, JA (1987). Aspectos de la historia natural del Octopus dofleini , el pulpo gigante del Pacífico. Maestría en Ciencias tesis. Departamento de Biología, Universidad de Victoria (Canadá). 101 págs.
Cosgrove, JA y N. McDaniel (2009). Súper chupadores: el pulpo gigante del Pacífico y otros cefalópodos de la costa del Pacífico. Harbour Publishing, Parque de Madeira. ISBN 1550174665 .
Costantino, G. (2014a). ¡Lance el kraken! Blog de la Biblioteca del Patrimonio de la Biodiversidad, 30 de octubre de 2014.
Costantino, G. (2014b). El Pulpo...El Monstruo que No Es. Blog de la Biblioteca del Patrimonio de la Biodiversidad, 30 de octubre de 2014.
Dall, WH (1885). Los brazos del pulpo o pez diablo. Ciencia 6 (145): 432. doi :10.1126/science.ns-6.145.432
Daniel, RJ (1925). Vida animal en el mar . Hodder & Stoughton, Londres. 119 págs.
Deagle, BE, SN Jarman, D. Pemberton y NJ Gales (2005). Detección genética de presas en el contenido intestinal de un calamar gigante ( Architeuthis sp.). Revista de herencia 96 (4): 417–423. doi :10.1093/jhered/esi036
Dell, RK (1952). Los cefalópodos recientes de Nueva Zelanda. Boletín del Museo Dominion 16 : 1–157.
de Montfort, PD (1801). Le Poulpe Colosal. [páginas. 256–412] En: Histoire naturallle, générale et particulière des mollusques, animaux sans vertèbres et a sang blanc. Ouvrage faisant suite aux œuvres de Leclerc de Buffon, et partie du cours complet d'histoire naturelle rédigé par CS Sonnini, miembro de plusieurs sociétés savantes. Tomo Segundo. L'Imprimerie de F. Dufart, París. 424 págs. doi :10.5962/bhl.title.51804 (en francés)
Dery, M. (2013). El Kraken despierta: lo que Architeuthis intenta decirnos. Boing Boing , 28 de enero de 2013.
Dommergues, J.-L., S. Montuire y P. Neige (2002). Patrones de tamaño a través del tiempo: el caso de la radiación de amonitas del Jurásico Temprano. Paleobiología 28 (4): 423–434. JSTOR 3595493
Doukan, G. (1957). Los hábitos del pulpo. [páginas. 67–70] En: Doukan, G. El mundo bajo las olas. John De Graff, Nueva York. 356 págs.
Dozier, TA (1976). Criaturas marinas peligrosas . Televisión Time-Life, Nueva York. 128 págs. ISBN 0-913948-04-7 .
Dunstan, AJ, PD Ward y NJ Marshall (2011). Historia de vida y demografía de Nautilus pompilius en el monte submarino Osprey Reef, Mar del Coral, Australia. MÁS UNO 6 (2): e16312. doi :10.1371/journal.pone.0016312
Ellis, R. (1994b). Architeuthis , el calamar gigante: una verdadera explicación para algunas serpientes marinas. El registro de Mystic Seaport 46 (2): 34–40.
Ellis, R. (1996). Atlántico profundo: vida, muerte y exploración en el abismo. Knopf, Nueva York. 395 págs. ISBN 0679433244 .
Ellis, R. (1997a). Los modelos de Architeuthis . Curador: The Museum Journal 40 (1): 30–55. doi :10.1111/j.2151-6952.1997.tb01120.x
Ellis, R. (1997b). El verdadero Architeuthis , aún no visto, pero un modelo más. Curador: The Museum Journal 40 (3): 176–177. doi :10.1111/j.2151-6952.1997.tb01301.x
Ellis, R. (1998b). ¿Qué tamaño alcanza el calamar gigante? La revisión de criptozoología 3 (1): 11-19.
Evon, D. (2016a). Kraken Rocken: Los informes sobre el posible avistamiento de un kraken cerca de la Antártida en realidad muestran una roca muy grande. Snopes.com , 20 de junio de 2016.
Evon, D. (2016b). Hay una playa: un vídeo que pretende mostrar un calamar de 50 pies de largo varado en Punakaiki, Nueva Zelanda, es un engaño. Snopes.com , 1 de diciembre de 2016.
Evon, D. (2017). ¿Se encontró un cadáver de calamar gigante en Indonesia? Las fotografías que supuestamente muestran un cadáver de calamar gigante varado en una playa de Indonesia en realidad muestran una ballena muerta. Snopes.com , 12 de mayo de 2017.
Eyden, P. (2003a). Belemnitas: una mirada rápida. La revista Octopus News en línea.
Eyden, P. (2003b). Tusoteuthis y los calamares gigantes del Cretácico. La revista Octopus News en línea.
Fairbairn, DJ (2007). Introducción: el enigma del dimorfismo de tamaño sexual. En: DJ Fairbairn, WU Blanckenhorn y T. Székely (eds.) Sexo, tamaño y roles de género: estudios evolutivos del dimorfismo del tamaño sexual. Prensa de la Universidad de Oxford, Oxford. págs. 1–10. ISBN 0199545588 . doi :10.1093/acprof:oso/9780199208784.003.0001
Farquhar, P. (2014). Los científicos encontraron solo el segundo calamar colosal intacto: así es como se ve. Business Insider Australia , 16 de septiembre de 2014.
Feldman, GC (1999). 20 de febrero: Ocho brazos para abrazarte. En: I. Roper, GC Feldman, CFE Roper , B. Brennan, D. Tracey, S. O'Shea y MV deGruy (1999). En busca de calamares gigantes: Nueva Zelanda - 1999: Diarios de expedición. Página de inicio web del Smithsonian de Historia Natural . [Archivado desde el original el 8 de octubre de 1999.]
Finn, JK (2013). Taxonomía y biología de los argonautas (Cephalopoda: Argonautidae) con especial referencia al material australiano. Investigación de moluscos 33 (3): 143–222. doi :10.1080/13235818.2013.824854
Finn, JK (2014a). Familia Alloposidae. En: P. Jereb, CFE Roper , MD Norman y JK Finn (eds.) Cefalópodos del mundo. Un catálogo comentado e ilustrado de especies de cefalópodos conocidas hasta la fecha. Volumen 3. Octópodos y calamares vampiros. Catálogo de especies de la FAO para fines pesqueros. No. 4, vol. 3. FAO, Roma. págs. 225-228.
Finn, JK (2014b). Familia Argonautidae. En: P. Jereb, CFE Roper , MD Norman y JK Finn (eds.) Cefalópodos del mundo. Un catálogo comentado e ilustrado de especies de cefalópodos conocidas hasta la fecha. Volumen 3. Octópodos y calamares vampiros. Catálogo de especies de la FAO para fines pesqueros. No. 4, vol. 3. FAO, Roma. págs. 228-237.
Flor, RH (1955). Estado de la clasificación de endoceroide. Revista de Paleontología 29 (3): 329–371. JSTOR 1300321
Förch, CE (1998). La fauna marina de Nueva Zelanda: Cephalopoda: Oegopsida: Architeuthidae (calamar gigante). Memoria de biodiversidad de NIWA 110 : 1–113. ISBN 0-478-08447-1 .
Francia, RL (2016a). Reinterpretar los relatos del siglo XIX sobre ballenas que luchan contra "serpientes marinas" como una ilusión de enredos tempranos en aparejos de pesca preplásticos o desechos marítimos. La Revista Internacional de Historia Marítima 28 (4): 686–714. doi :10.1177/0843871416667434
Francia, RL (2016b). Historicidad de las tortugas marinas identificadas erróneamente como monstruos marinos: un caso del enredo temprano de quelonios marinos en redes de pesca preplásticas y desechos marítimos. Coriolis 6 (2): 1–25.
Francia, RL (2017). Monstruos marinos imaginarios y amenazas ambientales reales: reconsiderando los famosos avistamientos de objetos marinos no identificados de Osborne, 'Moha-moha', Valhalla y 'Soay Beast'. Revista internacional de historia ambiental 3 (1): 63–100. doi :10.22459/IREH.03.01.2017.07
Frébold, H. (1957). "Género" Titanitas Buckman. En: The Jurassic Fernie Group en las Montañas Rocosas y las estribaciones de Canadá. Memoria del Servicio Geológico de Canadá 287 . Departamento de Minas y Estudios Técnicos de Canadá. págs. 66–67.
Frey, RC (1995). Cefalópodos nautiloides del Ordovícico medio y superior de la región del Arco de Cincinnati en Kentucky, Indiana y Ohio. Servicio Geológico de Estados Unidos, Denver.
Galbreath, GJ (2015). Identificada la 'serpiente enorme' del Dédalo de 1848. Investigador escéptico 39 (5): 42–46.
Gantès, RTF (1979). El misterio de los pies gigantes . Estudios Vivantes, París. 63 págs. ISBN 2-7310-1213-7 . (en francés)
Garcín, P.-Y. Y M. Raynal (2011). Tentáculos: de la ciencia a la ficción . Éditions Gaussen, Marsella. 144 págs. ISBN 2-356-98034-2 . (en francés)
Gennaro, Jr., JF (1971). Una trilogía de pulpos. Parte 2: la criatura revelada. Historia Natural 80 (3): 24, 84.
Gerhardt, MI (1966). Conocimiento en declive: información antigua y medieval sobre los "peces de tinta" y sus hábitos. Vivario 4 (1): 144–175. doi :10.1163/156853466X00079
Ghiselin, MT (1974). La economía de la naturaleza y la evolución del sexo. Prensa de la Universidad de California, Berkeley. 346 págs. ISBN 0-520-02474-5 .
Gibson, J. (1887). Monstruos del mar: legendarios y auténticos. Thomas Nelson and Sons, Londres. 138 págs. doi :10.5962/bhl.title.138418
Glaubrecht, M. y MA Salcedo-Vargas (2004). El calamar de Humboldt Dosidicus gigas (Orbigny, 1835): historia del espécimen de Berlín, con una reevaluación de otros cefalópodos gigantes (batí) pelágicos (Mollusca, Ommastrephidae, Architeuthidae). Mitteilungen aus dem Museum für Naturkunde en Berlín, Zoologische Reihe 80 (1): 53–69. doi :10.1002/mmnz.20040800105
Glenday, C. (ed.) (2014). Principalmente moluscos. [páginas. 62–63] En: Guinness World Records 2015. Guinness World Records. 255 págs. ISBN 1-908843-70-5
González, M., M. Fernández-Casado, MdP Rodríguez, A. Segura & JJ Martín (2000). Primer registro del calamar gigante Architeuthis sp. (Architeuthidae) en el Mar Mediterráneo. Revista de la Asociación de Biología Marina del Reino Unido 80 : 745–746 doi : 10.1017/S0025315400002630
González, Á.F., Á. Guerra y F. Rocha (2003). Nuevos datos sobre la historia de vida y la ecología del calamar anzuelo de aguas profundas Taningia danae. Sarsia 88 (4): 297–301.
Goodrich, SG (1861). Historia natural ilustrada del reino animal, que es una descripción sistemática y popular de los hábitos, estructura y clasificación de los animales desde las formas más altas hasta las más bajas, con sus relaciones con la agricultura, el comercio, las manufacturas y las artes. vol. I. Derby y Jackson, Nueva York. xv + 680 págs. doi :10.5962/bhl.title.118884
Gosse, PH (1861). Asalto de una sepia. [páginas. 235–237] En: El romance de la historia natural. Gould y Lincoln, Boston. xiv + 372 págs. doi :10.5962/bhl.title.3995
Goudsward, D. (2017). El legendario pulpo gigante de Florida: una historia. [sin paginar; 24 págs.] En: Revista de la Sociedad Internacional de Criptozoología: Volumen uno, 2016 . Museo Internacional de Criptozoología , Portland. 112 págs. ISBN 1-52-183755-4 .
Gould, C. (1886). Monstruos míticos. WH Allen & Co., Londres. 407 págs.
Conceder, G. (1896). El monstruo marino que llegó a la costa de Florida. Pennsylvania Grit , 13 de diciembre de 1896.
Greshko, M. (2016). El calamar gigante podría ser más grande que un autobús escolar. National Geographic , 27 de mayo de 2016.
Griggs, K. (2003). Súper calamar emerge en la Antártida. BBC News , 2 de abril de 2003.
Grist, EPM y GD Jackson (2007). ¿Cuánto tiempo tardaría en convertirse en un calamar gigante? Reseñas en Biología de peces y pesca 17 (2–3): 385–399. doi :10.1007/s11160-007-9046-x
Gröger, J., U. Piatkowski y H. Heinemann (2000). Análisis de la longitud del pico del calamar del Océano Austral Psychroteuthis glacialis (Cephalopoda: Psychroteuthidae) y su uso para la estimación de tamaño y biomasa. Biología polar 23 (1): 70–74. doi :10.1007/s003000050009
Grønningsaeter, A. (1946). Sjøormen-blekkspruten. Naturaleza 70 (12): 379–380. (en noruego)
Grulke, W. (2014). Heteromorfo: los amonites fósiles más raros: la naturaleza en su forma más extraña . En One Communications. 224 págs. ISBN 978-0-9929740-0-8 .
Grulke, W. (2016). Nautilus: Hermosa sobreviviente: 500 millones de años de historia evolutiva . En One Communications. 224 págs. ISBN 978-0-9929740-2-2 .
Guerra, Á. & Á.F. González (2009). ¿Qué sabemos de? El calamar gigante. CSIC y Los Libros de la Catarata, Madrid. 116 págs. ISBN 978-84-8319-466-9 . (en español)
Guerra, Á., Á.F. González, S. Pascual & EG Dawe (2011). El calamar gigante Architeuthis : un invertebrado emblemático que puede representar preocupación por la conservación de la biodiversidad marina. Conservación biológica 144 (7): 1989–1997. doi :10.1016/j.biocon.2011.04.021
Guerra, Á., Á.F. González y GJ Pierce (2018). Primer registro del varamiento de un calamar gigante vivo Architeuthis dux fuera de aguas japonesas. Ecología , publicado en línea el 23 de enero de 2018. doi :10.1002/ecy.2073
Guerra, Á., Á.F. González, F. Rocha, J. Gracia & L. Laria (2006). Enigmas de la Ciencia: El Calamar Gigante. CEPESMA, Vigo. 313 págs. ISBN 8460999998 . (en español)
Guerra, Á., AB Rodríguez-Navarro, Á.F. González, CS Romanek, P. Álvarez-Lloret & GJ Pierce (2010). Rasgos de la historia de vida del calamar gigante Architeuthis dux revelados a partir de firmas de isótopos estables registradas en los picos. Revista ICES de Ciencias Marinas 67 (7): 1425–1431. doi :10.1093/icesjms/fsq091
Guerra, Á. Y M. Segonzac (2014). Gigantes de profundidad. Éditions Quæ, Versalles. 143 págs. ISBN 2-7592-2220-9 . (en francés)
Haaker, PL (1985). Observaciones de un pulpo enano, Octopus micropyrsus . Conchas y vida marina 17 (1): 39–40.
Hallam, A. (1975). Aumento evolutivo del tamaño y longevidad en bivalvos y amonites del Jurásico. Naturaleza 258 : 493–496. doi :10.1038/258493a0
Hanlon, RT y JB Messenger (2018). Calamar grande y calamar gigante, Architeuthis. [páginas. 264–268] En: Comportamiento de los cefalópodos. Segunda edicion. Prensa de la Universidad de Cambridge, Cambridge. xv+365 págs. ISBN 1-108-54674-9 .
Hansford, D. (2009). Fotos raras: calamar gigante comido por cachalote. Noticias de National Geographic , 29 de octubre de 2009.
Harasewych, MG y F. Moretzsohn (2010). El libro de las conchas: una guía de tamaño natural para identificar y clasificar seiscientas conchas . A & C Black Publishers, Londres. ISBN 1408125277 .
Hashimoto, Y. (2017). Piratas, piratería y pulpos: ¿de monstruo con múltiples armas a minoría modelo? [páginas. 37–46] En: S. Inaga (ed.) La visión de un pirata de la historia mundial: una percepción invertida del orden de las cosas desde una perspectiva global. Centro Internacional de Investigación de Estudios Japoneses, Kioto. 174 págs.
Haszprunar, G. y A. Wanninger (2012). Moluscos. Biología actual 22 (13): R510 – R514. doi :10.1016/j.cub.2012.05.039
Hecht, J. (1995). El monstruo marino que nunca existió. Nuevo científico no. 1972, 8 de abril de 1995.
Heuvelmans, B. (1958). Dans le sillage des monstres marins: le kraken et le poulpe colossal . Plón, París. OCLC 16321559 (en francés) [Traducción al inglés: Heuvelmans, B. (2003). El Kraken y el pulpo colosal: tras los monstruos marinos. Kegan Paul Internacional, Londres. xv+332 págs. ISBN 0-7103-0870-1 .]
Hewitt, RA y JM Hurst (1977). Cambios de tamaño en amonitas liparoceratidas del Jurásico y su importancia estratigráfica y ecológica. Lethaia 10 (4): 287–301. doi :10.1111/j.1502-3931.1977.tb00623.x
Hickey, S. (2009). Súper calamar: el museo abrirá sus puertas el próximo mes. Anunciante (Grand Falls), 25 de mayo de 2009.
Hickey, S. (2010). Canada Post celebra el calamar gigante. Anunciante (Grand Falls), 17 de junio de 2010.
Alto, WL (1976). El pulpo gigante del Pacífico. Revista de pesca marina 38 (9): 17–22.
Hochberg, FG y WG Fields (1980). Cephalopoda: Los calamares y pulpos. En: RH Morris, DP Abbott y EC Haderlie (eds.) Invertebrados intermareales de California. Prensa de la Universidad de Stanford, Stanford. págs. 429–444. ISBN 0804710457 .
Hofilena, J. (2014). Calamar gigante de 4 metros de largo encontrado atrapado en una red de pesca frente a Japón. The Japan Daily Press, 13 de enero de 2014.
Hogenboom, M. (2014). ¿Son los enormes calamares realmente los monstruos marinos legendarios? BBC Earth , 12 de diciembre de 2014.
Holden, C. (1995). Un monstruo marino caído. Ciencia 268 (5208): 207–208. doi :10.1126/ciencia.268.5208.207-c
Holanda, CH (1987). Los cefalópodos nautiloideos: un extraño éxito. Revista de la Sociedad Geológica de Londres 144 (1): 1–15. doi :10.1144/gsjgs.144.1.0001
Hollenbeck, N. y D. Scheel (2017). Patrones corporales del pulpo gigante del Pacífico con volantes, una nueva especie de pulpo de Prince William Sound, Alaska. Boletín malacológico americano 35 (2): 134–144. doi :10.4003/006.035.0206
Hollenbeck, N., D. Scheel, MC Gravley, GK Sage, R. Toussaint y SL Talbot (2017). Uso de hisopos para muestreo de células epiteliales para análisis de genética molecular en Enteroctopus . Boletín malacológico americano 35 (2): 145–157. doi :10.4003/006.035.0207
Holroyd, J. (2005). Calamares nuevos en el bloque (de hielo). La edad , 21 de diciembre de 2005.
Hoving, HJT y SHD Haddock (2017). El pulpo gigante de aguas profundas Haliphron atlanticus se alimenta de fauna gelatinosa. Informes científicos 7 : 44952. doi : 10.1038/srep44952
Hoving, HJT y BH Robinson (2017). El ritmo de vida de los calamares que habitan en las profundidades. Investigación de aguas profundas, parte I: artículos de investigación oceanográfica 126 : 40–49. doi :10.1016/j.dsr.2017.05.005
Howard, J. (2017). ¿Existen los monstruos marinos? Sí, pero tienen otro nombre... The Guardian , 18 de mayo de 2017.
Humphries, S. y GD Ruxton (2002). ¿Por qué algunos ictiosaurios tenían ojos tan grandes? La Revista de Biología Experimental 205 (4): 439–441.
Iba, Y., S.-I. Sano y M. Goto (2015). Las grandes belemnitas ya eran comunes en el Jurásico Temprano: nueva evidencia del centro de Japón. Investigación paleontológica 19 (1): 21–25. doi :10.2517/2014PR025
Iversen, RTS, PJ Perkins y RD Dionne (1963). Una indicación de la producción de sonido submarino por parte de los calamares. Naturaleza 199 : 250–251. doi :10.1038/199250a0
Iwai, E. (1956). Descripciones de especies no identificadas de cefalópodos dibranquiados. I. Un calamar oegopsiden perteneciente al género Architeuthis . Los informes científicos del Instituto de Investigación de Ballenas 11 : 139–151.
Jackson, GD y ML Domeier (2003). Los efectos de un evento extraordinario de El Niño/La Niña en el tamaño y crecimiento del calamar Loligo opalescens frente al sur de California. Biología Marina 142 (5): 925–935. doi :10.1007/s00227-002-1005-4
Jereb, P. (2005). Familia Nautilidae. En: P. Jereb y CFE Roper , eds. Cefalópodos del mundo. Un catálogo comentado e ilustrado de especies conocidas hasta la fecha. Volumen 1. Nautilos y sepioides con cámara (Nautilidae, Sepiidae, Sepiolidae, Sepiadariidae, Idiosepiidae y Spirulidae) . Catálogo de especies de la FAO para fines pesqueros No. 4, vol. 1. FAO, Roma. págs. 51–55.
Jereb, P., M. Vecchione y CFE Roper (2010). Familia Loliginidae. En: P. Jereb y CFE Roper (eds.) Cefalópodos del mundo. Un catálogo comentado e ilustrado de especies conocidas hasta la fecha. Volumen 2. Calamares miopsidos y oegopsidos . Catálogo de especies de la FAO para fines pesqueros No. 4, vol. 2. FAO, Roma. págs. 38-117.
Johnson, Rhode Island (1989). Taxones de moluscos de Addison Emery Verrill y Katharine Jeannette Bush, incluidos los introducidos por Sanderson Smith y Alpheus Hyatt Verrill. Artículos ocasionales sobre moluscos 5 (67): 1–143. [ Pulpo giganteus en la pág. 39]
Johnson, Rhode Island (1998). El mito de Octopus giganteus Verrill, 1897: una ballena de historia. Artículos ocasionales sobre moluscos 5 (76): 456–457.
Kauffman, EG (1990). Depredación de peces del Cretácico sobre un calamar grande. [páginas. 195–196] En: AJ Boucot (ed.) Paleobiología evolutiva del comportamiento y la coevolución. Elsevier, Ámsterdam. xxiii+725 págs. ISBN 0-444-88034-8 .
Kawahara, M., S. Uye, K. Ohtsu y H. Iizumi (2006). Inusual explosión demográfica de la medusa gigante Nemopilema nomurai (Scyphozoa: Rhizostomeae) en aguas del este de Asia. Serie de progreso de la ecología marina 307 : 161–173. doi :10.3354/meps307161
Quilla, JA (1970). Criaturas extrañas del tiempo y el espacio . Publicaciones Fawcett, Greenwich. 288 págs. [ISBN de la edición Spearman de 1975: ISBN 0-85435-372-0 .]
Keil, A. (1963). Riesentintenfische aus dem Pottwal-Magen. Natur und Museum 93 (8): 319–323. (en alemán)
Kennedy, WJ (1977). [Evolución de amonita:] Cambios de tamaño evolutivo. En: A. Hallam (ed.) Patrones de evolución, ilustrados por el registro fósil. Compañía editorial científica Elsevier, Ámsterdam. págs. 288-290.
Kennedy, WJ y WA Cobban (1990). Amonitas micromórficas cenomanianas del interior occidental de EE. UU. Paleontología 33 (2): 379–422.
Kennedy, WJ y U. Kaplan (1995). Parapuzosia (Parapuzosia) seppenradensis (Landois) und die Ammonitenfauna der Dülmener Schichten, unteres Unter-Campan, Westfalen. Geologie und Paläontologie en Westfalen 33 : 1–127. ISBN 3-924590-44-3 . (en alemán)
Kin, A. y R. Niedźwiedzki (2012). Primer registro del género Pachydesmoceras de amonitas puzosiina del Turoniense medio y superior de Polonia. Investigación del Cretácico 33 (1): 15–20. doi :10.1016/j.cretres.2011.07.003
Kirk, TW (1880). Sobre la aparición de sepias gigantes en la costa de Nueva Zelanda. Transacciones y procedimientos del Instituto de Nueva Zelanda 12 (2): 310–313.
Kirk, TW (1888). Breve descripción de una nueva especie de decápodo grande (Architeuthis longimanus). Transacciones y procedimientos del Instituto de Nueva Zelanda 20 (1): 34–39.
Klug, C., K. De Baets, B. Kröger, MA Bell, D. Korn y JL Payne (2015). ¿Gigantes normales? Cambios temporales y latitudinales del gigantismo de invertebrados marinos paleozoicos y cambio global. Lethaia 48 (2): 267–288. doi :10.1111/let.12104
Knop, D. (1996). Almejas gigantes: una guía completa para la identificación y el cuidado de las almejas tridacnidas. Dähne Verlag, Ettlingen.
Korabelnikov, LV (1959). La dieta de los cachalotes en los mares antárticos. Priroda 3 : 103-104. (en ruso)
Kröger, B. (2003). El tamaño del sifúnculo en la evolución de los cefalópodos. Senckenbergiana Lethaea 83 : 39. doi : 10.1007/BF03043304
Kubodera, T. (2009).海に生きるものたちの掟: 食うか食われるか、命を受け継ぐ食物連鎖の秘密. SoftBank Creative, Tokio. 208 págs. ISBN 4797350717 . (en japonés)
Kubodera, T. (2010). Observaciones de grandes calamares mesopelágicos en estado salvaje utilizando equipos visuales submarinos desarrollados recientemente. [páginas. 22–42] En: S. Uchida (ed.) Actas de un simposio internacional: Hacia lo desconocido, investigando animales misteriosos de las profundidades marinas. 23 y 24 de febrero de 2007, Okinawa, Japón. Acuario Churaumi de Okinawa , Motobu . 179 págs. ISBN 4-9904177-0-4 .
Kubodera, T. (2013a).深海の怪物 ダイオウイカを追え! Popurasha, Tokio. 144 págs. ISBN 4591135152 . (en japonés)
Kubodera, T. (2013b).ダイオウイカ、奇跡の遭遇. Shinchosha, Tokio. 202 págs. ISBN 4103346914 . (en japonés)
Kubodera, T. y H. Horikawa (2005). Fauna de cefalópodos alrededor de las islas Nansei, en el sur de Japón. En: K. Hasegawa, G. Shinohara y M. Takeda (eds.) Fauna y contaminantes de las profundidades marinas en las islas Nansei . Monografías del Museo Nacional de Ciencias N° 29 . Museo Nacional de Ciencias, Tokio. págs. 191-223. NAID 110004677347
Kubodera, T., Y. Koyama y K. Mori (2006). Observaciones del comportamiento de caza salvaje y bioluminiscencia de un gran calamar de ocho brazos de aguas profundas, Taningia danae. Actas de la Royal Society B: Ciencias Biológicas 274 (1613): 1029–1034. doi :10.1098/rspb.2006.0236
Kubodera, T. y K. Mori (2005). Primeras observaciones de un calamar gigante vivo en la naturaleza. Actas de la Royal Society B: Ciencias Biológicas 272 (1581): 2583–2586. doi :10.1098/rspb.2005.3158
Kubodera, T., U. Piatkowski, T. Okutani y MR Clarke (1998). Taxonomía y zoogeografía de la familia Onychoteuthidae (Cephalopoda: Oegopsida). Contribuciones del Smithsonian a la zoología 586 (2): 277–291.
Kubodera, T., T. Wada, M. Higuchi y A. Yatabe (2016). Se encontró una cantidad extraordinaria de calamar gigante, Architeuthis dux , en las aguas costeras japonesas del Mar de Japón desde enero de 2014 hasta marzo de 2015. Marine Biodiversity , publicado en línea el 24 de diciembre de 2016. doi :10.1007/s12526-016-0618-7 [material complementario ]
Kummel, B. (1948). Importancia ambiental de los cefalópodos enanos. Revista de petrología sedimentaria 18 (2): 61–64. doi :10.1306/D4269305-2B26-11D7-8648000102C1865D
Tierra, MF y D.-E. Nilsson (2012). Ojos de animales. Segunda edicion. Prensa de la Universidad de Oxford, Oxford. xiii+271 págs. ISBN 0-19-958113-4 .
Landman, NH, JK Cochran, R. Cerrato, J. Mak, CFE Roper y CC Lu (2004). Hábitat y edad del calamar gigante (Architeuthis sanctipauli) inferidos a partir de análisis isotópicos. Biología Marina 144 (4): 685–691. doi :10.1007/s00227-003-1245-y
Landman, NH y R. Ellis (1998). Architeuthis ... por fin. Curador: The Museum Journal 41 (3): 150–152. doi :10.1111/j.2151-6952.1998.tb00828.x
Landman, NH, SM Klofak y KB Sarg (2003). Variación en el tamaño adulto de amonitas escafítidas de la Formación Pierre Shale y Fox Hills del Cretácico Superior. En: PJ Harries (ed.) Enfoques de alta resolución en paleontología estratigráfica . Saltador. págs. 149-194. doi :10.1007/978-1-4020-9053-0_5
Landois, H. (1895). Die Riesenammoniten von Seppenrade, Pachydiscus Zittel Seppenradensis H. Landois. Jahresbericht des Westfälischen Provinzial-Vereins für Wissenschaft und Kunst 23 : 99–108. (en alemán)
Landois, H. (1898). Gewichtsverhältnisse unserer Riesen-Ammoniten. Jahresbericht des Westfälischen Provinzial-Vereins für Wissenschaft und Kunst 26 : 27–28. (en alemán)
Carril, FW (1957). Reino del pulpo: la historia de vida de los cefalópodos. Editores Jarrolds, Londres. xx + 287 págs.
Laptikhovsky, V. (2006). Tendencias latitudinales y batimétricas en la variación del tamaño de los huevos: una nueva mirada a las reglas de Thorson y Rass. Ecología marina 27 (1): 7–14. doi :10.1111/j.1439-0485.2006.00077.x
Laptikhovsky, V., S. Nikolaeva y M. Rogov (2018). Conchas embrionarias de cefalópodos como herramienta para reconstruir estrategias reproductivas en taxones extintos. Reseñas biológicas 93 (1): 270–283. doi :10.1111/brv.12341
Laptikhovsky, VV, MA Rogov, SV Nikolaeva y AI Arkhipkin (2013). Impacto ambiental en las estrategias reproductivas de cefalópodos ectococleados y la importancia evolutiva del tamaño de los huevos de cefalópodos. Boletín de Geociencias 88 (1): 83–94.
Larson, NL (2010). Enchoteuthididae: cefalópodos gigantes del Cretácico Superior del Interior Occidental. [páginas. 66–78] En: JP Cavigelli (ed.) Tate 2010: 16ª Conferencia Anual de la Tate, 4 al 6 de junio de 2010. Beneath Wyoming Waves: Marine Paleontology. Museo Geológico Tate, Casper. 94 págs.
Larson, NL, SD Jorgensen, RA Farrar y PL Larson (1997). Amonitas y otros cefalópodos de Pierre Seaway . Prensa de geociencias, Tucson. ISBN 0-945005-25-3 .
Larson, RJ (1986). Contenido de agua, contenido orgánico y composición de carbono y nitrógeno de medusas del Pacífico nororiental. Revista de biología y ecología marina experimental 99 (2): 107–120. doi :10.1016/0022-0981(86)90231-5
LeBlond, PH y JR Sibert (1973). Observaciones de grandes animales marinos no identificados en Columbia Británica y aguas adyacentes. Informe manuscrito No. 28. Instituto de Oceanografía, Universidad de Columbia Británica, Vancouver. 64 + A-3 págs. doi :10.13140/RG.2.1.3821.9608
Lee, H. (1875). Sepias gigantescas. [páginas. 99-114] En: El pulpo; o el "pez diablo" de la ficción y de la realidad. Chapman y Hall, Londres. xv + 114 págs. doi :10.5962/bhl.title.10557
Lee, H. (1883). Monstruos marinos desenmascarados. William Clowes and Sons, Londres. vii + 103 págs. doi :10.5962/bhl.title.101985
Lee, JJ (2014). El engaño del calamar gigante de 160 pies: ¿Qué tamaño alcanzan realmente? National Geographic Daily News , 10 de enero de 2014.
Lehn, WH e I. Schroeder (1981). El tritón nórdico como fenómeno óptico. Naturaleza 289 : 362–366. doi :10.1038/289362a0
Leith, EI (1942). Notas sobre el cefalópodo Lambeoceras lambii de Manitoba. Revista de Paleontología 16 (1): 130–132.
Leporati, SC, GT Pecl y JM Semmens (2007). Crecimiento de las crías de cefalópodos: los efectos del tamaño inicial y las temperaturas estacionales. 151 : 1375. doi : 10.1007/s00227-006-0575-y
Lewy, Z. (2002). La función de los márgenes septales estriados de amonita. Revista de Paleontología 76 (1): 63–69. doi :10.1666/0022-3360(2002)076<0063:TFOTAF>2.0.CO;2
Liu, Y.-C., T.-H. Liu, C.-C. Yu, C.-H. Su & C.-C. Chiao (2017). Desajuste entre el ojo y el lóbulo óptico en el calamar gigante. Ciencia Abierta de la Royal Society 4 : 170289. doi : 10.1098/rsos.170289
Lipington, T. (2007-2009). Usuario: Tim Lipington . Wikipedia , 11 de octubre de 2007 - 17 de marzo de 2009.
Lipington, T. (2008). Avistamiento de un posible cefalópodo grande cerca de Portugal. Foros en línea de la revista Octopus News, 3 de abril de 2008.
Llinás, RR (1999). La sinapsis del calamar gigante: un modelo de transmisión química. Prensa de la Universidad de Oxford, Oxford. ISBN 0195116526 .
Lovis, P. (2011). El calamar colosal en exhibición recibe un chequeo. Blog Te Papa, 14 de febrero de 2011.
Lucas, FA (1897). El monstruo de Florida. Ciencia 5 (116): 476. doi :10.1126/science.5.116.476-a
Lucas, FA (1928). Algunos errores de los científicos: errores en la investigación cometidos honestamente y corregidos honestamente. Historia Natural 28 (2): 169–174.
Lynch, M. (2013). La búsqueda del superdepredador del océano. Corporación Australiana de Radiodifusión.
Lynnes, AS y PG Rodhouse (2002). Un gran bocado para los depredadores: el ejemplar más grande registrado de Kondakovia longimana (Cephalopoda: Onychoteuthidae). Boletín de Ciencias Marinas 71 (2): 1087–1090.
MacGinitie, GE y N. MacGinitie (1949). Historia Natural de los Animales Marinos . McGraw-Hill, Nueva York. xii + 473 págs.
MacGinitie, GE y N. MacGinitie (1968). Historia natural de los animales marinos. Segunda edicion. McGraw-Hill, Nueva York. 523 págs.
Maciá, S., MP Robinson, P. Craze, R. Dalton y JD Thomas (2004). Nuevas observaciones sobre la propulsión a chorro (vuelo) en el aire en calamares, con una revisión de informes anteriores. Revista de estudios de moluscos 70 (3): 297–299. doi :10.1093/mollus/70.3.297
Mackal, RP (1980). Pulpo giganteus Verrill. [páginas. 33–49] En: Búsqueda de animales ocultos: una investigación sobre misterios zoológicos . Doubleday, Ciudad Jardín. xxiii+294 págs. ISBN 0-385-14897-6 .
Mackal, RP (1986). Análisis bioquímicos de tejido conservado de Octopus giganteus. Criptozoología: Revista interdisciplinaria de la Sociedad Internacional de Criptozoología 5 : 55–62.
Manger, WL, LK Meeks y DA Stephen (1999). Gigantismo patológico en cefalópodos del Carbonífero medio, medio continente sur, Estados Unidos. En: F. Olóriz & FJ Rodríguez-Tovar (eds.) Avance de la investigación sobre cefalópodos vivos y fósiles . Kluwer Academic/Plenum Publishers, Nueva York. págs. 77–89. doi :10.1007/978-1-4615-4837-9_7
Mangiacopra, GS (1975a). Octopus giganteus Verrill: una nueva especie de cefalópodo. De mar y costa 6 (1): 3–10, 51–52.
Mangiacopra, GS (1975b). Batalla colosal. De Mar y Ribera 6 (4): 222.
Mangiacopra, GS (1976a). Los Grandes: una historia fragmentada del pulpo gigante y colosal. De mar y costa 7 (2): 93–96.
Mangiacopra, GS (1976b). ¿Una medusa monstruosa? De Mar y Costa 7 (3): 169.
Mangiacopra, GS (1977). Más sobre el pulpo giganteus. De Mar y Costa 8 (3): 174, 178.
Mangiacopra, GS (1999). Otra refutación de Octopus giganteus – ¡otra vez! De mar y costa 21 (4): 233–234.
Mangiacopra, GS, MPR Raynal, DG Smith y DF Avery (1994). Actualización sobre Octopus giganteus Verrill Parte 1: más fragmentos olvidados de su historia del siglo XIX. De mar y costa 17 (3): 171–178.
Mangiacopra, GS, MM Raynal, DG Smith y DF Avery (1995a). La historia se repite: Parte II. Ignorado – redescubierto – ¡ignorado de nuevo! – y reivindicación final de Octopus giganteus: 1909-1994. De mar y costa 17 (4): 221–225.
Mangiacopra, GS, MPR Raynal, DG Smith y DF Avery (1995b). Octopus giganteus: ¿sigue vivo y escondido dónde? Parte III: Lusca y cubas del Caribe. Del mar y la costa 18 (1): 5–12.
Mangiacopra, GS, DG Smith, DF Avery, M. Raynal, R. Ellis , JR Greenwell y B. Heuvelmans (1996). Un foro abierto sobre el artículo del Biological Bulletin sobre el análisis de tejidos del Octopus giganteus. De mar y costa 19 (1): 45–50.
Markey, M. (2010). El monstruo de San Agustín. Blog de archivos de la Institución Smithsonian, 18 de agosto de 2010.
Matthews, LH (1938). El cachalote, Physeter catodon . Informes de descubrimiento 17 : 93–168.
McCarey, K. y J. Rubin (directores) (1998). Monstruos marinos: busca el calamar gigante. [película documental] National Geographic Television. 55 minutos.
McClain, CR, MA Balk, MC Benfield, TA Branch, C. Chen, J. Cosgrove, ADM Dove, LC Gaskins, RR Helm, FG Hochberg, FB Lee, A. Marshall, SE McMurray, C. Schanche, SN Stone & AD Thaler (2015). Dimensionamiento de gigantes oceánicos: patrones de variación de tamaño intraespecífico en la megafauna marina. ParJ 3 : e715. doi :10.7717/peerj.715
McDowall, California (1998). ¿Calamares gigantes? The Marine Observer: Revista trimestral de meteorología marítima 68 (339): 39.
McMenamin, MAS (2012). Evidencia de un Kraken del Triásico: disposición inusual de los huesos en el Parque Estatal Ictiosaurio en Nevada. Ciencia y tecnología del siglo XXI 24 (4): 55–58.
McMenamin, MAS (2016). Huesos profundos. En: Paleontología dinámica de MAS McMenamin: uso de la cuantificación y otras herramientas para descifrar la historia de la vida . Springer, Cham. págs. 131-158. doi :10.1007/978-3-319-22777-1_9 ISBN 978-3-319-22776-4
McMenamin, MAS y D. Schulte McMenamin (2011). Kraken del Triásico: el conjunto mortuorio de ictiosaurios de Berlín interpretado como un basurero de cefalópodos gigante. Resúmenes de la Sociedad Geológica de América con programas 43 (5): 310.
McMenamin, MAS y D. Schulte McMenamin (2013). La espalda del Kraken: nueva evidencia sobre la posible disposición de los esqueletos de ictiosaurios en forma de cefalópodos. Resúmenes de la Sociedad Geológica de América con programas 45 (7): 900.
Meek, A. y TR Goddard (1926). Sobre dos ejemplares de calamar gigante varados en la costa de Northumberland. Transacciones de la Sociedad de Historia Natural de Northumberland, Durham y Newcastle-upon-Tyne , nueva serie, 6 (2): 229–237.
Mignot, Y., S. Elmi y J.-L. Dommergues (1993). Croissance et miniaturization de quelques Hildoceras (Cephalopoda) en liaison avec des environnements contraignants de la Téthys toarcienne. Geobios 26 (suplemento 1): 305–312. doi :10.1016/S0016-6995(06)80384-0 (en francés)
Mikkelson, D. (2014). Calamar gigante: ¿Ha llegado un calamar gigante a la costa de California? Snopes.com , 12 de enero de 2014.
Miller, AK y B. Kummel (1944). Algunos cefalópodos grandes y rectos del Ordovícico de Minnesota. Anales del Museo Carnegie 30 (2): 19–38.
Molinero, HW (1957). Niobrarateuthis bonneri , un nuevo género y especie de calamar de la Formación Niobrara de Kansas. Revista de Paleontología 31 (4): 809–811. JSTOR 1300484
Molinero, R. (1997). ¿Calamares o pulpo? Un misterio de 20.000 leguas de viaje submarino . Viajes Extraordinarios 3 (3): 3.
Molinero, R. (1998). 20.000 leguas de viaje submarino : el misterioso calamar gigante, tercera parte. Viajes extraordinarios 4 (2): 1–2.
Miller, WJ y FP Walter (1997). El controvertido calamar gigante de Verne: continuación. Viajes extraordinarios 4 (1): 1–5.
Milne, DH (1995). Vida Marina y el Mar. Compañía editorial Wadsworth, Belmont. xxii+459+A-28 págs. ISBN 0534163149 .
Mitsukuri, K. y S. Ikeda (1895). Notas sobre un cefalópodo gigantesco. Revista Zoológica 7 (77): 39–50. (en japonés)
Miyahara, K., K. Fukui, T. Ota y T. Minami (2006). Observaciones de laboratorio sobre las primeras etapas de vida del calamar diamante Thysanoteuthis rhombus . Revista de estudios de moluscos 72 (2): 199–205. doi :10.1093/mollus/eyi068
Monnet, C., A. Brayard y M. Brosse (2015). Tendencias evolutivas de los amonoides del Triásico. En: C. Klug, D. Korn, K. De Baets, I. Kruta y RH Mapes (eds.) Paleobiología amonoide: de la macroevolución a la paleogeografía . Springer, Dordrecht. págs. 25–50. doi :10.1007/978-94-017-9633-0_2 ISBN 978-94-017-9632-3
Mooney, TA, RT Hanlon, J. Christensen-Dalsgaard, PT Madsen, DR Ketten y PE Nachtigall (2010). Detección de sonido por el calamar de aleta larga ( Loligo pealeii ) estudiada con potenciales evocados auditivos: sensibilidad al movimiento de partículas de baja frecuencia y no a la presión. La Revista de Biología Experimental 213 : 3748–3759. doi :10.1242/jeb.048348
Morten, SD y RJ Twitchett (2009). Fluctuaciones en el tamaño corporal de los invertebrados marinos durante el evento de extinción pliensbachiano-toarciano. Paleogeografía, Paleoclimatología, Paleoecología 284 (1–2): 29–38. doi :10.1016/j.palaeo.2009.08.023
Mosendz, P. (2015). Fotos: raro calamar gigante encontrado frente a la costa de Hawái. Newsweek , 3 de octubre de 2015.
Motani, R., BM Rothschild y W. Wahl (1999). Globos oculares grandes en ictiosaurios buceadores. Naturaleza 402 : 747. doi : 10.1038/45435
Muntz, WRA (1995). Pulpos gigantes y calamares desde Plinio hasta el reverendo Moses Harvey. Archivos de Historia Natural 22 (1): 1–28. doi :10.3366/anh.1995.22.1.1
Muramatsu, K., J. Yamamoto, T. Abe, K. Sekiguchi, N. Hoshi y Y. Sakurai (2013). Los calamares oceánicos vuelan. Biología Marina 160 (5): 1171–1175. doi :10.1007/s00227-013-2169-9
Murata, M. (1988). Sobre el comportamiento de vuelo del calamar volador neón Ommastrephes bartrami observado en el centro y noroeste del Pacífico Norte. Nippon Suisan Gakkaishi 54 (7): 1167–1174. doi :10.2331/suisan.54.1167
Murray, A. (1874). Captura de un calamar gigante en Terranova. El naturalista americano 8 (2): 120–123. [Ver también corrección: Sepias gigantes de Terranova. El naturalista americano 8 (4): 226–227.]
Myers, PZ (2011). ¿Rastros de un Kraken del Triásico? Faringula , 10 de octubre de 2011.
Myers, PZ (2013). El hombre Kraken ha vuelto. Faringula , 1 de noviembre de 2013.
Nabhitabhata, J. (1998). Comportamiento distintivo del calamar pigmeo tailandés, Idiosepius thailandicus Chotiyaputta, Okutani & Chaitiamvong, 1991. Publicación especial del Centro de Biología Marina de Phuket 18 (1): 25–40.
Naish, D. (2017). Krakens, monstruos isleños, calamares gigantes y pulpos gigantes. [páginas. 51–60] En: Cazando monstruos: criptozoología y la realidad detrás de los mitos. Sirius Publishing, Londres. 240 págs. ISBN 1-78428-862-4 .
Nesis, KN (1970). Biología del calamar gigante peruano-chileno, Dosidicus gigas . Okeanologiya 10 (1): 140-152. (en ruso)
Nesis, KN (1982). Clave abreviada de los moluscos cefalópodos de los océanos del mundo . Editorial de la industria ligera y alimentaria, Moscú. 385 + ii págs. (en ruso) [Traducido al inglés por BS Levitov, ed. de LA Burgess (1987). (1987). Cefalópodos del mundo: calamares, sepias, pulpos y aliados . Publicaciones TFH, Neptune City.]
Nesis, KN (1985). Гигантский кальмар в Охотском море. [Un calamar gigante en el mar de Okhotsk.] Priroda [1985](10): 112–113. (en ruso)
Nesis, KN (1987). Cefalópodos del mundo: calamares, sepias, pulpos y aliados . Publicaciones TFH, Neptune City. ISBN 0-86622-051-8 .
Newell, Dakota del Norte (1949). Aumento del tamaño filético, una tendencia importante ilustrada por los invertebrados fósiles. Evolución 3 (2): 103–124. JSTOR 2405545
Newman, M. (1994). La vida en una pecera: confesiones del director de un acuario . Douglas y McIntyre, Vancouver. ix+262 págs. ISBN 1550541250 .
[ NHK ] (2013a).ダイオウイカと深海の生物. Gakken Paburisshingu, Tokio. 65 págs. ISBN 4056100438 . (en japonés)
[NHK] (2013b).深海の超巨大イカ. Shinnihon Shuppansha, Tokio. 32 págs. ISBN 4406057153 . (en japonés)
[NHK] (2013c). NHKスペシャル 世界初撮影!深海の超巨大イカ. [película documental] NHK Enterprises, Tokio. 73 minutos. (en japonés)
[NHK] (2014).伝説のイカ 宿命の闘い. Shinnihon Shuppansha, Tokio. 32 págs. ISBN 4406057331 . (en japonés)
Nicholls, EL y H. Isaak (1987). Importancia estratigráfica y taxonómica de Tusoteuthis longa Logan (Coleoidea, Teuthida) del miembro Pembina, Pierre Shale (Campaniano), de Manitoba. Revista de Paleontología 61 (4): 727–737. doi :10.1017/S0022336000029085
Ruiseñor, A. (2012). ¿Quién quiere almorzar? Un pulpo que se alimenta de gaviotas en Victoria, BC. BirdFellow, 27 de abril de 2012.
Nigmatullin, CM, KN Nesis y AI Arkhipkin (2001). Una revisión de la biología del calamar gigante Dosidicus gigas (Cephalopoda: Ommastrephidae). Investigación pesquera 54 (1): 9–19. doi :10.1016/S0165-7836(01)00371-X
Nilsson, D.-E., EJ Warrant, S. Johnsen, R. Hanlon y N. Shashar (2012). Una ventaja única para los ojos gigantes en los calamares gigantes. Biología actual 22 (8): 683–688. doi :10.1016/j.cub.2012.02.031
Nilsson, D.-E., EJ Warrant, S. Johnsen, RT Hanlon y N. Shashar (2013). Los ojos gigantes de los calamares gigantes son de hecho inesperadamente grandes, pero no si se usan para observar cachalotes. BMC Biología Evolutiva 13 : 187. doi : 10.1186/1471-2148-13-187
Norman, MD y JK Finn (2014). Familia Vampyroteuthidae. En: P. Jereb, CFE Roper , MD Norman y JK Finn (eds.) Cefalópodos del mundo. Un catálogo comentado e ilustrado de especies de cefalópodos conocidas hasta la fecha. Volumen 3. Octópodos y calamares vampiros. Catálogo de especies de la FAO para fines pesqueros. No. 4, vol. 3. FAO, Roma. págs. 268-270.
Norman, MD, JK Finn y FG Hochberg (2014). Familia Octopodidae. En: P. Jereb, CFE Roper , MD Norman y JK Finn (eds.) Cefalópodos del mundo. Un catálogo comentado e ilustrado de especies de cefalópodos conocidas hasta la fecha. Volumen 3. Octópodos y calamares vampiros. Catálogo de especies de la FAO para fines pesqueros. No. 4, vol. 3. FAO, Roma. págs. 36-215.
Norman, MD, D. Paul, J. Finn y T. Tregenza (2002). Primer encuentro con un pulpo manta macho vivo: el animal grande con mayor dimorfismo sexual del mundo. Revista de Investigación Marina y de Agua Dulce de Nueva Zelanda 36 (4): 733–736. doi :10.1080/00288330.2002.9517126
O'Dor, RK, EA Foy, PL Helm y N. Balch (1986). La locomoción y la energía de las crías de calamar, Illex illecebrosus. Boletín malacológico americano 4 (1): 55–60.
O'Dor, RK y JA Hoar (2000). ¿La geometría limita el crecimiento de los calamares? Revista ICES de Ciencias Marinas 57 (1): 8–14. doi :10.1006/jmsc.1999.0502
O'Dor, RK, J. Stewart, W. Gilly, J. Payne, TC Borges y T. Thys (2013). Ciencia espacial de los calamares: cómo los calamares se lanzan al aire. Investigación de aguas profundas, parte II: estudios temáticos en oceanografía 95 : 113–118. doi :10.1016/j.dsr2.2012.07.002
Okutani, T. (1995). Sepias y calamares del mundo en color . Publicación por el 30 aniversario de la fundación de la Asociación Cooperativa Nacional de Procesadores de Calamar. 185 págs.
Okutani, T. (2015a). Megalocranchia maxima Pfeffer, 1884. En: Sepias y calamares del mundo: nueva edición. Asociación Nacional Cooperativa de Procesadores de Calamar. 246 págs. ISBN 4486037340 .
Okutani, T. (2015b). Onykia (Onykia) robusta (Verrill, 1876). En: Sepias y Calamares del Mundo: Nueva Edición. Asociación Nacional Cooperativa de Procesadores de Calamar. 246 págs. ISBN 4486037340 .
Olivero, EB y WJ Zinsmeister (1989). Grandes amonites heteromorfos del Cretácico Superior de la Isla Seymour, Antártida. Revista de Paleontología 63 (5): 626–636. JSTOR 1305622
Omori, M. y M. Kitamura (2004). Revisión taxonómica de tres especies japonesas de medusas comestibles (Scyphozoa: Rhizostomeae). Biología y ecología del plancton 51 (1): 36–51.
O'Shea, S. (2002). Haliphron atlanticus es un pulpo gelatinoso gigante. Actualización de Biodiversidad 5 : 1.
O'Shea, S. (2003a). Hoja informativa sobre el calamar gigante y el calamar colosal. La revista Octopus News en línea. [Archivado desde el original el 7 de diciembre de 2003.]
O'Shea, S. (2003b). Kondakovia longimana. En: Hoja informativa sobre el calamar gigante y el calamar colosal. La revista Octopus News en línea. [Archivado desde el original el 27 de abril de 2006.]
O'Shea, S. (2003c). Re: Necropsia del calamar colosal. Foros en línea de la revista Octopus News, 15 de agosto de 2003.
O'Shea, S. (2003d) Re: Living Architeuthis foto. Foros en línea de la revista Octopus News, 4 de octubre de 2003. [Archivado desde el original el 4 de febrero de 2013.]
O'Shea, S. (2004a). El pulpo gigante Haliphron atlanticus (Mollusca: Octopoda) en aguas de Nueva Zelanda. Revista de Zoología de Nueva Zelanda 31 (1): 7–13. doi :10.1080/03014223.2004.9518353
O'Shea, S. (2004b). Re: Estreno de AFO (Calamar colosal/Cachalote). Foros en línea de la revista Octopus News, 25 de septiembre de 2004. [Archivado desde el original el 15 de diciembre de 2007.]
O'Shea, S. (2006). Estimación de la edad y la tasa de crecimiento en Architeuthis dux. La revista Octopus News en línea.
O'Shea, S. y K. Bolstad (2008). Hoja informativa sobre el calamar gigante y el calamar colosal. La revista Octopus News en línea.
Oudemans, AC (1892). La gran serpiente marina. Un tratado histórico y crítico. EJ Brill, Leiden. xv + 592 págs. doi :10.5962/bhl.title.63980
Owen, R. (1881). Cefalópodos destacables por su gran tamaño. [páginas. 156–168] En: Descripciones de algunos Cephalopoda nuevos y raros. (Parte II.) Transacciones de la Sociedad Zoológica de Londres 11 (5): 131–170. doi :10.1111/j.1096-3642.1881.tb00354.x
Perdiz, JC (2012). Ecología sensorial: ¿ojos gigantes para depredadores gigantes? Biología actual 22 (8): R268 – R270. doi :10.1016/j.cub.2012.03.021
Paxton, CGM (2003). Los peligros de una racionalización excesiva o de calamares gigantes son pistas falsas. El inteligente escéptico 6 : 19-28. [Reimpreso en una forma ligeramente modificada: Paxton, CGM (2004). Los calamares gigantes son pistas falsas: por qué Architeuthis es una fuente poco probable de avistamientos de monstruos marinos. The Cryptozoology Review 4 (2): 10-16.]
Paxton, CGM (2009). El plural de "anécdota" puede ser "datos": análisis estadístico de distancias de visualización en informes de grandes animales marinos no identificados 1758-2000. Revista de Zoología 279 (4): 381–387. doi :10.1111/j.1469-7998.2009.00630.x
Paxton, CGM (2016a). Desatando al Kraken: sobre la longitud máxima en el calamar gigante ( Architeuthis sp.). Revista de Zoología 300 (2): 82–88. doi :10.1111/jzo.12347
Paxton, CGM (2016b). Los calamares gigantes que cada vez son más grandes. [transmisión de radio] World Update , 2 de junio de 2016.
Paxton, CGM y R. Holanda (2005). ¿Tenía razón Steenstrup? Una nueva interpretación del monje marino de Øresund del siglo XVI. Steenstrupiá 29 (1): 39–47.
Paxton, CGM, E. Knatterud y SL Hedley (2005). Cetáceos, sexo y serpientes marinas: un análisis de los relatos de Egede sobre un "monstruo más espantoso" visto frente a la costa de Groenlandia en 1734. Archives of Natural History 32 (1): 1–9. doi :10.3366/anh.2005.32.1.1
Paxton, CGM y AJ Shine (2016). Una lista de hipótesis históricas sobre el monstruo del lago Ness. La Revista de Criptozoología 4 : 21–37.
Payne, JL, AG Boyer, JH Brown, S. Finnegan, M. Kowalewski, RA Krause, Jr., SK Lyons, CR McClain, DW McShea, PM Novack-Gottshall, FA Smith, JA Stempien y SC Wang (2009). El aumento en dos fases del tamaño máximo de la vida a lo largo de 3.500 millones de años refleja la innovación biológica y las oportunidades ambientales. PNAS 106 (1): 24–27. doi :10.1073/pnas.0806314106
[ PBS ] (2012). Dentro de los gigantes de la naturaleza: el calamar gigante. [película documental] Películas inesperadas. 53 minutos.
Peattie, N. (1996). Hydra y Kraken: o la tradición y el atractivo de los monstruos del lago y las serpientes marinas. Prensa regente, Oakland. 56 págs. ISBN 0-916147-99-1 .
Pecl, GT, MA Steer y KE Hodgson (2004). El papel del tamaño de las crías en la generación de la variabilidad intrínseca del tamaño por edad de los cefalópodos: ampliando la hipótesis de Forsythe. Investigación marina y de agua dulce 55 (4): 387–394. doi :10.1071/MF03153
Phillips, JB (1933). Descripción de un calamar gigante capturado en Monterey, con notas sobre otros calamares capturados en la costa de California. Pesca y caza de California 19 (2): 128–136.
Phillips, JB (1961). Dos cefalópodos inusuales tomados cerca de Monterey. Pesca y caza de California 47 (4): 416–417.
Pierce, SK, SE Massey, NE Curtis, GN Smith, C. Olavarría y TK Maugel (2004). Características microscópicas, bioquímicas y moleculares de la Mancha Chilena y una comparación con los restos de otros monstruos marinos: nada más que ballenas. El Boletín Biológico 206 (3): 125-133. doi :10.2307/1543636
Pierce, SK, GN Smith, TK Maugel y E. Clark (1995). Sobre el pulpo gigante (Octopus giganteus) y el Bermuda Blob: homenaje a AE Verrill. El Boletín Biológico 188 (2): 219–230. doi :10.2307/1542087
Pohle, A. y C. Klug (2017). Tamaño corporal de cefalópodos ortocónicos del Silúrico tardío y Devónico del Antiatlas (Marruecos). Lethaia 51 (1): 126–148. doi :10.1111/let.12234
Poppe, GT e Y. Goto (1993). Conchas europeas. vol. II: Scaphopoda, Bivalvia, Cephalopoda. Christa Hemmen, Wiesbaden. 221 págs. ISBN 3-925919-10-4 .
Potier, M., F. Ménard, HD Benivary & R. Sabatié (2011). Estimaciones de longitud y peso a partir de estructuras diagnósticas de partes duras de especies forrajeras de peces, crustáceos y cefalópodos en el Océano Índico occidental. Biología ambiental de los peces 92 (3): 413–423. doi :10.1007/s10641-011-9848-5
Poulton, TP (1989). Amonitas del Jurásico (Oxfordiano) de la Formación Fernie del oeste de Canadá: un peltoceratinido gigante y Cardioceras canadense Whiteaves. Contribuciones a la paleontología canadiense, Servicio Geológico de Canadá, Boletín 396 : 173–179.
Prothero, D. (2013). Krakens y chiflados... otra vez. blog escéptico , 6 de noviembre de 2013.
Puig, R. (2004). Una ballena de cuento. Research.Online , 16 de agosto de 2004. [Archivado desde el original el 24 de noviembre de 2005.]
Raynal, M. (1987). Propiedades del colágeno y naturaleza del Monstruo de Florida. Criptozoología: Revista interdisciplinaria de la Sociedad Internacional de Criptozoología 6 : 129–130.
Raynal, M. (2000). Le "Monstre de Floride" de 1896: ¿cachalot ou pieuvre geante? Institut Virtuel de Cryptozoologie, 1 de mayo de 2000. (en francés)
Raynal, M. (2002). Les pieuvres géantes de Sidmouth: una nouvelle de cryptozoologie-fiction. Institut Virtuel de Cryptozoologie, 9 de marzo de 2002. (en francés)
Raynal, M. (2003a). Le dossier du poulpe colosal. Institut Virtuel de Cryptozoologie, 10 de mayo de 2003. (en francés)
Raynal, M. (2003b). Les poulpes géants du Pacifique. Institut Virtuel de Cryptozoologie, 10 de mayo de 2003. (en francés)
Raynal, M. (2017). El "monstruo de Florida" de 1896: un siglo de controversia. [sin paginar; 17 págs.] En: Revista de la Sociedad Internacional de Criptozoología: Volumen uno, 2016 . Museo Internacional de Criptozoología , Portland. 112 págs. ISBN 1-52-183755-4 .
Raynal, M. y M. Dethier (1991). Le «Monstre de Floride» de 1896: ¿cétacé ou poulpe colosal? Boletín de la Société Neuchâteloise des Sciences Naturelles 114 : 105–115. doi :10.5169/seals-89341 (en francés)
Reboulet, S. (2001). Factores limitantes en el crecimiento de la concha, el modo de vida y la segregación de las poblaciones de ammonoideos de valanginiano: evidencia de variaciones en el tamaño de los adultos. Geobios 34 (4): 423–435. doi :10.1016/S0016-6995(01)80006-1
Rees, WJ (1950). Sobre un calamar gigante, Ommastrephes caroli Furtado, varado en Looe, Cornwall. Boletín del Museo Británico (Historia Natural), Zoología 1 (2): 29–42.
Reid, A. (2005). Familia Spirulidae. En: P. Jereb y CFE Roper , eds. Cefalópodos del mundo. Un catálogo comentado e ilustrado de especies conocidas hasta la fecha. Volumen 1. Nautilos y sepioides con cámara (Nautilidae, Sepiidae, Sepiolidae, Sepiadariidae, Idiosepiidae y Spirulidae) . Catálogo de especies de la FAO para fines pesqueros No. 4, vol. 1. FAO, Roma. págs. 211-212.
Reid, A. y P. Jereb (2005). Familia Sepiolidae. En: P. Jereb y CFE Roper , eds. Cefalópodos del mundo. Un catálogo comentado e ilustrado de especies conocidas hasta la fecha. Volumen 1. Nautilos y sepioides con cámara (Nautilidae, Sepiidae, Sepiolidae, Sepiadariidae, Idiosepiidae y Spirulidae) . Catálogo de especies de la FAO para fines pesqueros No. 4, vol. 1. FAO, Roma. págs. 153-203.
Reid, A., P. Jereb y CFE Roper (2005). Familia Sepiidae. En: P. Jereb y CFE Roper, eds. Cefalópodos del mundo. Un catálogo comentado e ilustrado de especies conocidas hasta la fecha. Volumen 1. Nautilos y sepioides con cámara (Nautilidae, Sepiidae, Sepiolidae, Sepiadariidae, Idiosepiidae y Spirulidae) . Catálogo de especies de la FAO para fines pesqueros No. 4, vol. 1. FAO, Roma. págs. 57-152.
Arroz, T. (1995). El informe más reciente sobre "Octopus giganteus". De Mar y Ribera 18 (3): 172.
Arroz, T. (1998). Pulpo giganteus otra vez. De Mar y Costa 21 (1): 52.
Roach, M. (2013). La lucha del pez diablo y el pulpo. Entre líneas: Blog de la biblioteca estatal de Washington, 26 de septiembre de 2013.
Robin, M.-P. (1952). Combate de monstruos. Le Chasseur français , noviembre de 1952. p. 701. (en francés)
Robson, GC (1933). Sobre Architeuthis clarkei , una nueva especie de calamar gigante, con observaciones sobre el género. Actas de la Sociedad Zoológica de Londres 1933 (3): 681–697. doi :10.1111/j.1096-3642.1933.tb01614.x
Roeleveld, MAC (2000). Picos de calamar gigante: implicaciones para la sistemática. Revista de la Asociación de Biología Marina del Reino Unido 80 (1): 185–187. doi :10.1017/S0025315499001769
Roeleveld, MAC (2002). Morfología del tentáculo del calamar gigante Architeuthis de los océanos Atlántico Norte y Pacífico. Boletín de Ciencias Marinas 71 (2): 725–737.
Romanov, EV, S. Jaquemet y L. Puetz (2017). Un calamar gigante ( Architeuthis dux ) frente a la isla Reunión, en el Océano Índico occidental: ¿el gigante más grande de todos los tiempos? Revista de la Asociación de Biología Marina del Reino Unido , publicada en línea el 12 de septiembre de 2017. doi :10.1017/S0025315417001588
Roper, CFE y KJ Boss (1982). El calamar gigante. Científico americano 246 (4): 15, 96–100, 104–105. JSTOR 24966572
Roper, CFE y P. Jereb (2010a). Familia Architeuthidae. En: P. Jereb y CFE Roper (eds.) Cefalópodos del mundo. Un catálogo comentado e ilustrado de especies conocidas hasta la fecha. Volumen 2. Calamares miopsidos y oegopsidos . Catálogo de especies de la FAO para fines pesqueros No. 4, vol. 2. FAO, Roma. págs. 121-123.
Roper, CFE y P. Jereb (2010b). Familia Chiroteuthidae. En: P. Jereb y CFE Roper (eds.) Cefalópodos del mundo. Un catálogo comentado e ilustrado de especies conocidas hasta la fecha. Volumen 2. Calamares miopsidos y oegopsidos . Catálogo de especies de la FAO para fines pesqueros No. 4, vol. 2. FAO, Roma. págs. 135-145.
Roper, CFE y P. Jereb (2010c). Familia Cranchiidae. En: P. Jereb y CFE Roper (eds.) Cefalópodos del mundo. Un catálogo comentado e ilustrado de especies conocidas hasta la fecha. Volumen 2. Calamares miopsidos y oegopsidos . Catálogo de especies de la FAO para fines pesqueros No. 4, vol. 2. FAO, Roma. págs. 148-178.
Roper, CFE y P. Jereb (2010). Familia Cycloteuthidae. En: P. Jereb y CFE Roper (eds.) Cefalópodos del mundo. Un catálogo comentado e ilustrado de especies conocidas hasta la fecha. Volumen 2. Calamares miopsidos y oegopsidos . Catálogo de especies de la FAO para fines pesqueros No. 4, vol. 2. FAO, Roma. págs. 179–182.
Roper, CFE y P. Jereb (2010d). Familia Lepidoteuthidae. En: P. Jereb y CFE Roper (eds.) Cefalópodos del mundo. Un catálogo comentado e ilustrado de especies conocidas hasta la fecha. Volumen 2. Calamares miopsidos y oegopsidos . Catálogo de especies de la FAO para fines pesqueros No. 4, vol. 2. FAO, Roma. págs. 239-240.
Roper, CFE y P. Jereb (2010e). Familia Magnapinnidae. En: P. Jereb y CFE Roper (eds.) Cefalópodos del mundo. Un catálogo comentado e ilustrado de especies conocidas hasta la fecha. Volumen 2. Calamares miopsidos y oegopsidos . Catálogo de especies de la FAO para fines pesqueros No. 4, vol. 2. FAO, Roma. págs. 247–249.
Roper, CFE y P. Jereb (2010 y siguientes). Familia Mastigoteuthidae. En: P. Jereb y CFE Roper (eds.) Cefalópodos del mundo. Un catálogo comentado e ilustrado de especies conocidas hasta la fecha. Volumen 2. Calamares miopsidos y oegopsidos . Catálogo de especies de la FAO para fines pesqueros No. 4, vol. 2. FAO, Roma. págs. 250–256.
Roper, CFE y P. Jereb (2010g). Familia Octopoteuthidae. En: P. Jereb y CFE Roper (eds.) Cefalópodos del mundo. Un catálogo comentado e ilustrado de especies conocidas hasta la fecha. Volumen 2. Calamares miopsidos y oegopsidos . Catálogo de especies de la FAO para fines pesqueros No. 4, vol. 2. FAO, Roma. págs. 262–268.
Roper, CFE y P. Jereb (2010h). Familia Onychoteuthidae. En: P. Jereb y CFE Roper (eds.) Cefalópodos del mundo. Un catálogo comentado e ilustrado de especies conocidas hasta la fecha. Volumen 2. Calamares miopsidos y oegopsidos . Catálogo de especies de la FAO para fines pesqueros No. 4, vol. 2. FAO, Roma. págs. 348–369.
Roper, CFE y P. Jereb (2010i). Familia Pholidoteuthidae. En: P. Jereb y CFE Roper (eds.) Cefalópodos del mundo. Un catálogo comentado e ilustrado de especies conocidas hasta la fecha. Volumen 2. Calamares miopsidos y oegopsidos . Catálogo de especies de la FAO para fines pesqueros No. 4, vol. 2. FAO, Roma. págs. 370–373.
Roper, CFE y P. Jereb (2010j). Familia Thysanoteuthidae. En: P. Jereb y CFE Roper (eds.) Cefalópodos del mundo. Un catálogo comentado e ilustrado de especies conocidas hasta la fecha. Volumen 2. Calamares miopsidos y oegopsidos . Catálogo de especies de la FAO para fines pesqueros No. 4, vol. 2. FAO, Roma. págs. 384–387.
Roper, CFE, EM Jorgensen, ON Katugin y P. Jereb (2010). Familia Gonatidae. En: P. Jereb y CFE Roper (eds.) Cefalópodos del mundo. Un catálogo comentado e ilustrado de especies conocidas hasta la fecha. Volumen 2. Calamares miopsidos y oegopsidos . Catálogo de especies de la FAO para fines pesqueros No. 4, vol. 2. FAO, Roma. págs. 200–222.
Roper, CFE, C. Nigmatullin y P. Jereb (2010). Familia Ommastrephidae. En: P. Jereb y CFE Roper (eds.) Cefalópodos del mundo. Un catálogo comentado e ilustrado de especies conocidas hasta la fecha. Volumen 2. Calamares miopsidos y oegopsidos . Catálogo de especies de la FAO para fines pesqueros No. 4, vol. 2. FAO, Roma. págs. 269–347.
Roper, CFE y EK Shea (2013). Las preguntas sin respuesta sobre el calamar gigante Architeuthis (Architeuthidae) ilustran nuestro conocimiento incompleto de los cefalópodos coleoideos. Boletín malacológico americano 31 (1): 109–122. doi :10.4003/006.031.0104
Roper CFE, MJ Sweeney y CE Nauen (1984). Cefalópodos del mundo . Organización para la Agricultura y la Alimentación, Roma, Italia.
Roper, CFE y M. Vecchione (1993). Una revisión geográfica y taxonómica de Taningia danae Joubin, 1931 (Cephalopoda: Octopoteuthidae), con nuevos registros y observaciones sobre bioluminiscencia. En: T. Okutani, RK O'Dor y T. Kubodera (eds.) Avances recientes en la biología de la pesca de cefalópodos . Prensa de la Universidad de Tokai, Tokio. págs. 441–456.
Roper, CFE y GL Voss (1983). Directrices para descripciones taxonómicas de especies de cefalópodos. Memorias del Museo Nacional de Victoria 44 : 48–63.
Roper, CFE y RE Young (1972). Primeros registros de calamar gigante juvenil, Architeuthis (Cephalopoda: Oegopsida). Actas de la Sociedad Biológica de Washington 85 (16): 205–222.
Rosa, R., L. González, HM Dierssen & BA Seibel (2012). Determinantes ambientales de las tendencias de tamaño latitudinal en cefalópodos. Serie de progreso de la ecología marina 464 : 153–165. JSTOR 24876065
Rosa, R., VM Lopes, M. Guerreiro, K. Bolstad & JC Xavier (2017). Biología y ecología del invertebrado más grande del mundo, el calamar colosal ( Mesonychoteuthis hamiltoni ): una breve reseña. Biología polar 40 (9): 1871–1883. doi :10.1007/s00300-017-2104-5
Rosa, R. y BA Seibel (2010). Ritmo de vida lento del calamar colosal antártico. Revista de la Asociación de Biología Marina del Reino Unido 90 (7): 1375–1378. doi :10.1017/S0025315409991494
Agua de rosas, JR (1965). La familia Tridacnidae en el Indo-Pacífico. Moluscos del Indo-Pacífico 1 (6): 347–408.
Sakamoto, S. (2013).ドキュメント 深海の超巨大イカを追え! Kobunsha, Tokio. 262 págs. ISBN 4334037534 . (en japonés)
Salcedo-Vargas, MA (1999). Un calamar asperoteutido (Mollusca: Cephalopoda: Chiroteuthidae) de Nueva Zelanda identificado erróneamente como Architeuthis . Mitteilungen aus dem Museum für Naturkunde en Berlín, Zoologische Reihe 75 (1): 47–49. doi :10.1002/mmnz.19990750106
Salvador, RB y BM Tomotani (2014). El Kraken: cuando el mito se encuentra con la ciencia. História, Ciências, Saúde – Manguinhos 21 (3): 971–994. doi :10.1590/S0104-59702014000300010
Sanderson, TI (1956). Sigue a la ballena. Little, Brown y compañía, Boston. 423 págs.
Santos, MB, GJ Pierce, Á.F. González, F. Santos, MA Vázquez, MA Santos & MA Collins (2001). Primeras citas de Taningia danae (Cephalopoda: Octopoteuthidae) en aguas gallegas (noroeste de España) y en aguas escocesas (Reino Unido). Revista de la Asociación de Biología Marina del Reino Unido 81 (2): 355–356. doi :10.1017/S0025315401003903
Sazima, I. y LB de Almeida (2008). El pájaro kraken: el pulpo se alimenta de un ave marina en una isla oceánica en el Atlántico occidental tropical. Registros de biodiversidad marina 1 : e47. doi :10.1017/S1755267206005458
Scheel, D. y R. Anderson (2012). Variabilidad en la especialización de la dieta de Enteroctopus dofleini (Cephalopoda: Octopodidae) en el Pacífico oriental examinada a partir del contenido de los basurales. Boletín malacológico americano 30 (2): 267–279. doi :10.4003/006.030.0206
Schlegelmilch, R. (1998). Die Belemniten des süddeutschen Jura: Ein Bestimmungsbuch für Geowissenschaftler und Fossiliensammler . Gustav Fisher, Stuttgart. ISBN 3437255266 . doi :10.1007/978-3-8274-3083-0 (en alemán)
Schmitz, L., R. Motani, CE Oufiero, CH Martin, MD McGee, AR Gamarra, JJ Lee y PC Wainwright (2013a). La alometría indica que los ojos gigantes de los calamares gigantes no son excepcionales. BMC Biología Evolutiva 13 : 45. doi : 10.1186/1471-2148-13-45
Schmitz, L., R. Motani, CE Oufiero, CH Martin, MD McGee y PC Wainwright (2013b). La posible mejora de la capacidad de los calamares gigantes para detectar cachalotes es una exaptación ligada a su gran tamaño corporal. BMC Biología Evolutiva 13 : 226. doi : 10.1186/1471-2148-13-226
Scott, G. y MH Moore (1928). Amonitas de enorme tamaño del Cretácico de Texas. Revista de Paleontología 2 (4): 273–279. JSTOR 1298015
Scully, T. (2008). Neurociencia: La gran caza del calamar. Naturaleza 454 (7207): 934–936. doi :10.1038/454934a
Sharpe, FA, MA Kyte, WA Kyte y M. Kyte (1990). Depredación del pulpo sobre aves marinas. Boletín del Grupo de Aves Marinas del Pacífico 17 (1): 34.
Zapatero, MT (1995). El misterio criptozoológico se profundiza. Revista INFORMACIÓN 73 : 49–50.
Sitges, FG (director) (2003). Proyecto Kraken: En Busca del Calamar Gigante. [película documental] Transglobe Films. 52 minutos. (en español) [Versión en inglés: Proyecto Kraken: En busca del calamar gigante.]
Smith, FA, JL Payne, NA Heim, MA Balk, S. Finnegan, M. Kowalewski, SK Lyons, CR McClain, DW McShea, PM Novack-Gottshall, PS Anich y SC Wang (2016). Evolución del tamaño corporal a lo largo del Geozoico. Revista anual de ciencias planetarias y de la Tierra 44 : 523–553. doi : 10.1146/annurev-earth-060115-012147
Smith, RD (1996). Más sobre "Octopus giganteus". De Mar y Costa 18 (4): 194, 220.
Snÿder, R. (1995). La forma gigante de Sthenoteuthis oualaniensis del Mar Arábigo. Maestría en Ciencias tesis, Universidad de Aberdeen, Aberdeen. 70 págs.
Snÿder, R. (1998). Aspectos de la biología de la forma gigante de Sthenoteuthis oualaniensis (Cephalopoda: Ommastrephidae) del Mar Arábigo. Revista de estudios de moluscos 64 (1): 21–34. doi :10.1093/mollus/64.1.21
Starkey, JD (1963). Vi un monstruo marino. Animales 2 (23): 629, 644.
Staub, F. (1993). Requin bleu, calmar géant et cachalot. Actas de la Real Sociedad de Artes y Ciencias de Mauricio 5 (3): 141–145. (en francés)
Staudinger, MD, F. Juanes y S. Carlson (2009). Reconstrucción del tamaño corporal original y estimación de relaciones alométricas para el calamar costero de aleta larga (Loligo pealeii) y el calamar de aleta corta del norte (Illex illecebrosus). Boletín de pesca 107 (1): 101–105.
Stevens, GR (1978a). Érase una vez un amonita gigante. Boletín de la Sociedad Geológica de Nueva Zelanda 45 : 3–5.
Stevens, GR (1978b). La historia de un amonita gigante. Cazador de rocas 11 (4): 2–9.
Stevens, GR (1979a). La historia continua de un amonites gigante. Cazador de rocas 13 (1): 33–35.
Stevens, GR (1979b). La historia continua de un amonites gigante. Boletín de la Sociedad Geológica de Nueva Zelanda 48 : 6–9.
Stevens, GR (1985). Una revisión de Lytoceratinae (subclase Ammonoidea), incluida Lytoceras taharoaense n. sp., Jurásico Superior, Nueva Zelanda. Revista de Geología y Geofísica de Nueva Zelanda 28 (1): 153–185. doi :10.1080/00288306.1985.10422282
Stevens, GR (1988). Amonitas gigantes: una revisión. En: J. Wiedmann y J. Kullmann (eds.) Cefalópodos: presente y pasado: OH Schindewolf Symposium Tübingen 1985 . E. Schweizerbart'sche Verlagsbuchhandlung, Stuttgart. págs. 141-166.
Stothers, RB (2004). Base científica antigua de la "gran serpiente" a partir de evidencia histórica. Isis 95 (2): 220–238. doi :10.1086/426195
Sucik, N. (2000). Justo cuando parecía seguro hacer snorkel: los pulpos gigantes de Hawái. Revisión norteamericana de BioFortean 3 (2): 11–17.
Summesberger, H. (1979). Eine obersantone Ammonitenfauna aus dem Becken von Gosau (Oberösterreich). Annalen des Naturhistorischen Museums Wien 82 : 109-176. (en alemán)
Sweeney, MJ y CFE Roper (2001). Registros de especímenes de Architeuthis de informes publicados. Museo Nacional de Historia Natural, Institución Smithsonian. 132 págs.
Switek, B. (2011a). El Calamar Gigante: Dragón de las Profundidades. Smithsonian , 21 de junio de 2011.
Switek, B. (2011b). El calamar gigante prehistórico que se comía el sentido común. Cableado , 10 de octubre de 2011.
Espadas, MD (1991). El Wasgo o Sisiutl: un animal marino criptozoológico de la costa noroeste del Pacífico de América. Revista de exploración científica 5 (1): 85–101. [ fuente poco confiable? ]
Tanabe, K., Y. Hikida e Y. Iba (2006). Dos mandíbulas coleoides del Cretácico Superior de Hokkaido, Japón. Revista de Paleontología 80 (1): 138-145. doi :10.1666/0022-3360(2006)080[0138:TCJFTU2.0.CO;2]
Tanabe, K., A. Misaki y T. Ubukata (2015). Registro del Cretácico tardío de grandes coleoides de cuerpo blando basado en restos de mandíbula inferior de Hokkaido, Japón. Acta Paleontológica Polonica 60 (1): 27–38. doi : 10.4202/app.00057.2013
Teichert, C. (1927). Der estländische Glint. Natur und Museum 57 (6): 264–272. (en alemán)
Teichert, C. y B. Kummel (1960). Tamaño de los cefalópodos endoceroides. Museo Breviora de Zoología Comparada 128 : 1–7.
Tratz, EP (1973). Der Riesenkalmar ( Architeuthis ) im Haus der Natur. Haus der Natur, Salzburgo. 15 págs. OCLC 721114691 (en alemán)
Trueman, AE (1940). La cámara del cuerpo de amonita, con especial referencia a la flotabilidad y modo de vida de la amonita viva. Revista trimestral de la Sociedad Geológica de Londres 96 (1–4): 339–383. doi :10.1144/GSL.JGS.1940.096.01-04.14
Tsuchiya, K. y T. Okutani (1993). Calamares raros e interesantes en Japón -X. Apariciones recientes de calamares grandes de Okinawa. Venus 52 : 299-311.
Vecchione, M., KM Mangold y RE Young (2008). Cirrata Grimpe, 1916. Proyecto Web Árbol de la Vida.
Vecchione, M., RE Young , Á. Guerra, DJ Lindsay, DA Clague, JM Bernhard, WW Sager, Á.F. González, FJ Rocha & M. Segonzac (2001a). Observaciones mundiales de notables calamares de aguas profundas. Ciencia 294 (5551): 2505–2506. doi :10.1126/ciencia.294.5551.2505
Vecchione, M., RE Young , Á. Guerra, DJ Lindsay, DA Clague, JM Bernhard, WW Sager, Á.F. González, FJ Rocha & M. Segonzac (2001b). Cefalópodos en acción. Museo Nacional Smithsonian de Historia Natural.
Vecchione, M., RE Young y K. Tsuchiya (2011). Onykia (Onykia) robsoni (Adam, 1962). Proyecto Web Árbol de la Vida.
Vergano, D. (2013). ¿El antiguo monstruo marino convirtió a sus presas en arte? National Geographic Daily News , 30 de octubre de 2013. [Archivado desde el original el 2 de noviembre de 2013.]
Vermeij, GJ (2016). El gigantismo y sus implicaciones para la historia de la vida. MÁS UNO 11 (1): e0146092. doi :10.1371/journal.pone.0146092
Verrill, AE (1874b). Las sepias gigantes de Terranova y los calamares comunes de la costa de Nueva Inglaterra. El naturalista americano 8 (3): 167–174. JSTOR 2448747
Verrill, AE (1875a). Los colosales cefalópodos del Atlántico norte. El naturalista americano 9 (1): 21–36. JSTOR 2448361
Verrill, AE (1875b). Los colosales cefalópodos del Atlántico norte, II. El naturalista americano 9 (2): 78–86. JSTOR 2448586
Verrill, AE (1875c). Breves contribuciones a la zoología del Museo de Yale College. No. XXXV.—Aviso de la aparición de otro cefalópodo gigantesco (Architeuthis) en la costa de Terranova, en diciembre de 1874. The American Journal of Science and Arts , serie 3, 10 (57): 213–214. doi :10.2475/ajs.s3-10.57.213 [Reimpreso: Verrill, AE (1875). Aviso de la aparición de otro cefalópodo gigantesco (Architeuthis) en la costa de Terranova, en diciembre de 1874. The Annals and Magazine of Natural History , serie 4, 16 (94): 266–268. doi :10.1080/00222937508681845]
Verrill, AE (1878). Aviso de incorporaciones recientes a la fauna marina de la costa oriental de América del Norte. The American Journal of Science and Arts , serie 3, 16 (93): 207–215. doi :10.2475/ajs.s3-16.93.207
Verrill, AE (1880a). Los cefalópodos de la costa nororiental de América. Parte I.—Los calamares gigantes (Architeuthis) y sus aliados; con observaciones de especies grandes similares de localidades extranjeras. Transacciones de la Academia de Artes y Ciencias de Connecticut 5 (5): 177–257. doi :10.5962/bhl.title.35965
Verrill, AE (1880b). Sinopsis de los Cefalópodos de la costa nororiental de América. The American Journal of Science , serie 3, 19 (112): 284–295.
Verrill, AE (1881a). Calamar gigante (Architeuthis) abundante en 1875, en los Grandes Bancos. The American Journal of Science , serie 3, 21 (123): 251–252. [Reimpreso: Verrill, AE (1881). Calamar gigante (Architeuthis) abundante en 1875 en los Grandes Bancos. Anales y revista de historia natural , series 5, 7 (40): 351–352. doi :10.1080/00222938109459527]
Verrill, AE (1881b). Los cefalópodos de la costa nororiental de América. Parte II. Los cefalópodos más pequeños, incluidos los "calamares" y los pulpos, con otras formas afines. Transacciones de la Academia de Artes y Ciencias de Connecticut 5 (5): 259–446.
Verrill, AE (1882a). Aparición de un espécimen adicional de Architeuthis en Terranova. The American Journal of Science , serie 3, 23 (133): 71–72.
Verrill, AE (1882b). [Revisión de] Descripciones de algunos cefalópodos nuevos y raros. Parte II. [...] Por Richard Owen. The American Journal of Science , serie 3, 23 (133): 72–75.
Verrill, AE (1882c). [Revisión de] Descripción de nuevos Cephalopoda; por TW Kirk. The American Journal of Science , serie 3, 24 (144): 477.
Verrill, AE (1897a). Un cefalópodo gigantesco de la costa de Florida. The American Journal of Science , serie 4, 3 (13): 79. [Reimpreso: Verrill, AE (1897). Un cefalópodo gigantesco de la costa de Florida. Los Anales y Revista de Historia Natural , serie 6, 19 (110): 240. doi :10.1080/00222939708680533]
Verrill, AE (1897b). Pulpo gigantesco. New York Herald , 3 de enero de 1897. p. 13.
Verrill, AE (1897c). Información adicional sobre el cefalópodo gigante de Florida. The American Journal of Science , serie 4, 3 (14): 162–163.
Verrill, AE (1897d). Un monstruo marino frente a la costa de Florida. New York Herald , 14 de febrero de 1897. p. 5.
Verrill, AE (1897e). El monstruo de Florida. Ciencia 5 (114): 392. doi :10.1126/ciencia.5.114.392
Verrill, AE (1897 y siguientes). ¿Qué es esta criatura? New York Herald , 7 de marzo de 1897. p. 13.
Verrill, AE (1897g). El monstruo marino de Florida. Ciencia 5 (116): 476. doi :10.1126/ciencia.5.116.476
Verrill, AE (1897h). El monstruo marino de Florida. El naturalista americano 31 (364): 304–307. doi :10.1086/276596
Verrill, AE (1897i). El supuesto gran pulpo de Florida; Ciertamente no es un cefalópodo. The American Journal of Science , serie 4, 3 (16): 355–356. [Reimpreso: Verrill, AE (1897). El supuesto gran pulpo de Florida: ciertamente no es un cefalópodo. Anales y revista de historia natural , serie 6, 19 (114): 682–683. doi :10.1080/00222939708680668]
Verrill, AH (1897a). Pulpo para New Haven. Registro vespertino de New Haven , 14 de febrero de 1897.
Verril, AH (1917). El océano y sus misterios. Duffield & Co., Nueva York. xv + 189 págs. doi :10.5962/bhl.title.29018
Verrill, AH (1952). La extraña historia de nuestra Tierra: un panorama del crecimiento de nuestro planeta revelado por las ciencias de la geología y la paleontología . LC Page & Co., Boston. xviii + 255 págs.
Verrill, GE (1958). La ascendencia, la vida y la obra de Addison E. Verrill de la Universidad de Yale. Compañía Editorial de la Costa del Pacífico, Santa Bárbara. vii + 99 págs. OCLC 427636826
Voss, GL (1959). Cazando monstruos marinos. Sea Frontiers: Boletín de la Fundación Oceanográfica Internacional 5 (3): 134–146.
Voss, GL (1988). Evolución y relaciones filogenéticas de los octópodos de aguas profundas (Cirrata e Incirrata). [páginas. 253–276] En: MR Clarke y ER Trueman (eds.). Los moluscos. Volumen 12. Paleontología y Neontología de Cefalópodos. Prensa académica, San Diego. xxiv+355 págs. ISBN 1-4832-4852-6 .
Caminante, M. (2010). Descubierto el órgano sexual del súper calamar. BBC Earth News , 7 de julio de 2010.
Walker, M. (2016). El monstruo marino gigante de 27 metros de largo: los calamares gigantes pueden alcanzar longitudes extraordinarias. BBC Tierra , 27 de mayo de 2016.
Watts, J. (2015). Monstruos marinos. Buzo , abril de 2015.
Webb, D. (1897). Un gran decápodo. El Nautilus 10 : 108.
Webb, WF (1945). Un catálogo de moluscos recientes de todas partes del mundo. Walter F. Webb, San Petersburgo, Florida. doi :10.5962/bhl.título.1714
Weis, R. y N. Mariotti (2007). Fauna de belemnitas de los lechos fronterizos entre Aaleniano y Bajociano del Gran Ducado de Luxemburgo (Cuenca NE de París). Bollettino della Società Paleontologica Italiana 46 (2–3): 149–174.
Weiss, R. (1995a). Los científicos hunden la historia de un monstruo marino. Las masas de carne no provienen de un pulpo colosal, concluye un estudio. The Washington Post , 2 de abril de 1995. p. A18.
Weiss, R. (1995b). Los científicos dicen que esas masas marinas no eran monstruos. Hartford Courant , 3 de abril de 1995. p. 2.
Westermann, GEG (1966). ¿El holotipo (plastotipo) de? Titanites occidentalis Frebold de la arenisca de Kootenay (Jurásico superior) del sur de Columbia Británica. Revista Canadiense de Ciencias de la Tierra 3 (5): 623–625. doi :10.1139/e66-043
Westwood, B. (2015). Calamar gigante. [transmisión de radio] BBC Radio 4 , 23 de junio de 2015.
Whitaker, I. (1986). Criaturas marinas del Atlántico norte en el Espejo del Rey ( Konungs Skuggsjá ). Registro polar 23 (142): 3–13. doi :10.1017/S0032247400006756
Winkelmann, I., PF Campos, J. Strugnell, Y. Cherel, PJ Smith, T. Kubodera , L. Allcock, M.-L. Kampmann, H. Schroeder, Á. Guerra, M. Norman , J. Finn, D. Ingrao, M. Clarke y MTP Gilbert (2013). Diversidad del genoma mitocondrial y estructura poblacional del calamar gigante Architeuthis : la genética arroja nueva luz sobre una de las especies marinas más enigmáticas. Actas de la Royal Society B: Ciencias Biológicas 280 (1759): 20130273. doi :10.1098/rspb.2013.0273
Wolff, GA (1981). Una clave de pico para ocho especies de cefalópodos del Pacífico tropical oriental con relaciones entre las dimensiones y el tamaño de su pico. Boletín de pesca 80 (2): 357–370.
Wolff, GA (1984). Identificación y estimación del tamaño de los picos de 18 especies de cefalópodos del Océano Pacífico. Informe técnico de la NOAA NMFS 17, NOAA/Servicio Nacional de Pesca Marina.
Wong, K. (2000). Calamares de gran tamaño. Scientific American , 8 de noviembre de 2000.
Madera, FG (1957). Pulpo San Agustín de seis toneladas. Mariner , septiembre de 1957. p. 4.
Madera, FG (1971a). Una trilogía de pulpos. Parte 1: coloso asombroso de las profundidades. Historia Natural 80 (3): 14–16, 18, 20–24.
Madera, FG (1971b). Una trilogía de pulpos. Parte 3: en la que los pescadores de las Bahamas cuentan sus aventuras con la bestia. Historia Natural 80 (3): 84, 86–87.
Madera, GL (1982). El Libro Guinness de hechos y hazañas animales . Tercera edicion. Superlativos Guinness, Londres. 252 págs. ISBN 0-85112-235-3 .
Wood, JB y RK O'Dor (2000). ¿Los cefalópodos más grandes viven más tiempo? Efectos de la temperatura y la filogenia en las comparaciones interespecíficas de edad y tamaño en la madurez. Biología Marina 136 (1): 91–99. doi :10.1007/s002270050012
Wormuth, JH (1976). La biogeografía y taxonomía numérica de la familia de calamares oegopsidos Ommastrephidae en el Océano Pacífico. Boletín del Instituto Scripps de Oceanografía 23 : 1–96.
Wright, CW y WJ Kennedy (1980). Origen, evolución y sistemática de la acantoceratida enana Protacanthoceras Spath, 1923 (Cretácico Ammonoidea). Boletín del Museo Británico (Historia Natural): Geología 34 (2): 65–107.
Xavier, JC, PG Rodhouse, PN Trathan y AG Wood (1999). Un Atlas del Sistema de Información Geográfica (SIG) de la distribución de cefalópodos en el Océano Austral. Ciencia Antártica 11 (1): 61–62. doi :10.1017/S0954102099000097 [datos complementarios en línea con mapas]
Yasuda, T. (2004).日本近海における巨大エチゼンクラゲ Nemopilema nomurai の大発生について. [Sobre la aparición inusual de la medusa gigante, Nemopilema nomurai en aguas japonesas.] Boletín de la Sociedad Japonesa de Pesca Científica 70 (3): 380–386. NAID 110003145750 (en japonés)
Joven, E. (2003). Un calamar gigante emerge de las profundidades del océano. Nuevo científico no. 2390, 12 de abril de 2003. p. 18.
Young, RE y KM Mangold (2010). Megalocranchia Pfeffer, 1884. Proyecto web Árbol de la vida.
Young, RE, CFE Roper y M. Vecchione (2015). Heteroteuthinae Appellof, 1898. Proyecto web Árbol de la vida.
Yujov, VL (1974). El descubrimiento de los calamares gigantes. Priroda [1974](6): 60–63. (en ruso)
Yujov, VL (2014). Гігантські кальмари роду Architeuthis у Південному океані / Calmantes gigantes Architeuthis en el océano Austral. [Hihantski kalmary rodu Architeuthis u Pivdennomu okeani.] Ucrania Antártica Journal 13 : 242–253. (en ucraniano)
Zeidberg, LD (2004). Las mediciones de alometría a partir de grabaciones de video in situ pueden determinar el tamaño y la velocidad de nado del calamar juvenil y adulto Loligo opalescens (Cephalopoda: Myopsida). La Revista de Biología Experimental 207 (24): 4195–4203. doi :10.1242/jeb.01276
Zeidler, W. (1981). Un calamar gigante de aguas profundas, Taningia sp., de aguas del sur de Australia. Transacciones de la Royal Society of South Australia 105 (4): 218.
Zuyev, G., C. Nigmatullin, M. Chesalin y KN Nesis (2002). Principales resultados de estudios mundiales a largo plazo sobre el género de calamares oceánicos nectónicos tropicales Sthenoteuthis: una visión general de las investigaciones soviéticas. Boletín de Ciencias Marinas 71 (2): 1019–1060.






.jpg/1280px-Tremoctopus_violaceus5_(rotated).jpg)





.jpg/1280px-Taningia_danae_(160_cm_ML).jpg)



.jpg/1280px-Parapuzosia_seppenradensis_(Plate_II).jpg)










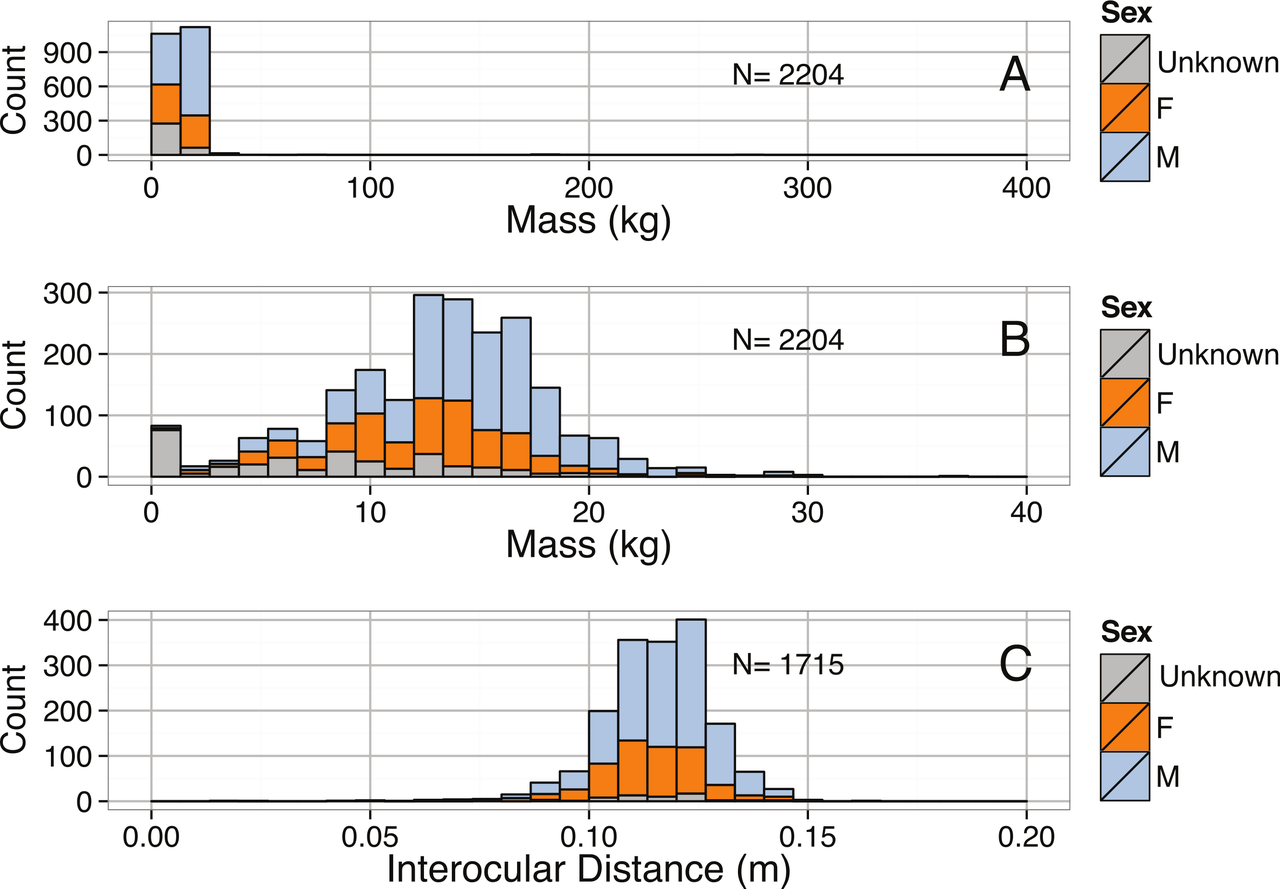










,_Northern_Hawaiian_waters.jpg/1280px-Ommastrephes_bartramii_(Neon_flying_squid),_Northern_Hawaiian_waters.jpg)






_dark_coloration.jpg/1280px-Sepia_latimanus_(Reef_cuttlefish)_dark_coloration.jpg)






.jpg/1280px-Parapuzosia_seppenradensis_(Plate_I).jpg)
























