Receptor 5-HT3
[2] Están más estrechamente relacionados por homología con el receptor nicotínico de acetilcolina.
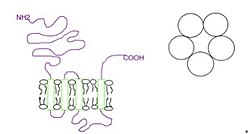
El receptor 5-HT3 difiere notablemente en estructura y mecanismo de los otros subtipos de receptores 5-HT acoplados a proteína G. Por lo general, el canal funcional está compuesto por cinco subunidades 5-HT3A idénticas (homopentaméricas).
Aún no se ha identificado el papel fisiopatológico de estas subunidades adicionales.
[14] A nivel celular, se ha demostrado que los receptores postsinápticos 5-HT3 median la transmisión sináptica excitatoria rápida en las interneuronas neocorticales de rata, la amígdala y el hipocampo, y en la corteza visual del hurón.
[14] Sin embargo, con el descubrimiento de que el receptor 5-HT3 desempeña un papel destacado en los vómitos inducidos por quimioterapia y radioterapia, y el desarrollo concomitante de antagonistas selectivos del receptor 5-HT 3 para suprimir estos efectos secundarios, despertó un gran interés en la industria farmacéutica.
[2][33] Desde entonces, siguió rápidamente la identificación de receptores 5-HT3 en líneas celulares y tejidos nativos.