Los aetosaurios ( / eɪˌɛt oʊˈsɔːr / ) son reptiles fuertemente acorazados pertenecientes al orden extinto Aetosauria ( / eɪˌɛt oʊˈsɔːr iə / ; del griego , ἀετός ( aetos , «águila») y σαυρος ( sauros , «lagarto»)). Eran pseudosuquios omnívoros o herbívoros de tamaño mediano a grande , parte de la rama de los arcosaurios más estrechamente relacionados con los cocodrilos que con las aves y otros dinosaurios . Todos los aetosaurios conocidos están restringidos al Triásico Superior , y en algunos estratos de esta época se encuentran entre los vertebrados fósiles más abundantes . Tienen cabezas pequeñas, hocicos respingados, extremidades erectas y un cuerpo adornado con cuatro filas de osteodermos en forma de placa ( escudos óseos ). Se conocen restos fósiles de aetosaurios en Europa , América del Norte y del Sur , partes de África y la India . Dado que sus placas acorazadas a menudo se conservan y son abundantes en ciertas localidades, los aetosaurios sirven como importantes fósiles índice de tetrápodos del Triásico Tardío . Muchos aetosaurios tenían amplias áreas de distribución geográfica, pero sus áreas de distribución estratigráficas eran relativamente cortas. Por lo tanto, la presencia de aetosaurios particulares puede datar con precisión un sitio en el que se encuentran. [1] [2] [3]
Casi todos los aetosaurios (excepto el género Aetosauroides ) pertenecen a la familia Stagonolepididae . Se han descrito más de 20 géneros de aetosaurios, y recientemente ha habido controversia con respecto a la descripción de algunos de estos géneros. Actualmente se reconocen dos subdivisiones distintas de aetosaurios, Desmatosuchia y Aetosaurinae , basadas principalmente en amplias diferencias en la morfología del cráneo . [4] La estructura de los osteodermos es generalmente uno de los rasgos más útiles para inferir las relaciones de los aetosaurios con mayor precisión. Entre otros arcosaurios, los aetosaurios están más estrechamente relacionados con Revueltosaurus , un pequeño reptil conocido originalmente a partir de dientes erróneamente referidos a dinosaurios herbívoros. [3]
Los restos de aetosaurios fueron descubiertos por primera vez a principios del siglo XIX, aunque los primeros restos que se describieron se confundieron con escamas de peces . Los aetosaurios fueron reconocidos más tarde como parientes de los cocodrilos , momento en el que se los interpretó como carroñeros semiacuáticos estrechamente relacionados con los fitosaurios . Trabajos posteriores han establecido que los aetosaurios eran animales completamente terrestres y probablemente herbívoros hasta cierto punto. Algunas formas tienen características que pueden haber sido adaptaciones para cavar en busca de alimento. Las supuestas estructuras de anidación también se han referido a los aetosaurios, pero esta conexión se considera ambigua. [3]

El cráneo de los aetosaurios es relativamente pequeño en comparación con el cuerpo, y tiene una forma bastante distintiva. Los dientes están ausentes tanto en la parte frontal del premaxilar (el hueso que forma la punta del hocico) como en la parte frontal del dentario (el hueso dentado de la mandíbula inferior). [5] [4] Los dientes que están presentes son generalmente pequeños y bulbosos, que van desde formas cónicas básicas hasta formas similares a hojas con grandes dentados. Estos son probablemente indicativos de una dieta omnívora o herbívora , y se observan adaptaciones similares en otros arcosaurios con menos dependencia de la carne en su dieta. [6] [3] Algunos aetosaurios tienen dientes con forma de zifodonte, lo que significa que los dientes son curvados, dentados y aplanados desde el lado. Esta forma, que es predominante en Aetosauroides y un pequeño espécimen tentativamente referido como Coahomasuchus , [4] es típica de los arcosaurios carnívoros . [7]
En algunos aetosaurios (en particular, los miembros del grupo Desmatosuchia), la punta del hocico se expande lateralmente en una forma aplanada de "pala", similar al hocico de un cerdo . Las fosas nasales externas (agujeros de las fosas nasales) son alargadas, mucho más grandes que las fenestras antorbitales (un agujero en el costado del cráneo). Muchos aetosaurios tienen una pequeña protuberancia en el premaxilar que se proyecta hacia las fosas nasales desde abajo. En todos los aetosaurios excepto Aetosauroides , el borde posterior de las fosas nasales recibe una contribución del borde frontal cóncavo del hueso maxilar . En la parte superior trasera del cráneo, un agujero conocido como fenestra supratemporal está posicionado y expuesto en el costado, a diferencia de la mayoría de los otros arcosaurios donde es principalmente visible cuando se ve el cráneo desde arriba. [8] [5] [4] La caja craneana es bastante estándar para los estándares de los pseudosuquios, aunque la abertura para el nervio abducens pasa a través del hueso parabasisfenoides (en la parte frontal inferior de la caja craneana), en lugar del hueso proótico (en la parte frontal superior). Este rasgo solo se ve en Revueltosaurus y crocodilomorfos entre los arcosaurios. [9] [5] [10] La mandíbula (mandíbula inferior) se describe como con forma de "zapatilla" en muchos aetosaurios. Esto se debe a una combinación de características: la parte frontal del dentario se estrecha fuertemente hasta un punto, mientras que la parte inferior del dentario a veces se flexiona en un "mentón" (proyección hacia abajo) que también puede exponer el hueso esplenial . La articulación de la mandíbula está situada en una posición baja, y el articular (el hueso de la mandíbula inferior que se conecta al cráneo) a menudo tiene una proyección alta justo detrás de la articulación de la mandíbula. [8] [4] [7]

En la mayoría de los aspectos, aparte de su cráneo y armadura, la anatomía esquelética de los aetosaurios era bastante estándar entre otros grandes pseudosuquios del Triásico. Las extremidades traseras desarrollaron una postura de extremidades "erguidas en forma de pilar" similar a la observada en los " rauisuquios ", un grado relacionado de pseudosuquios carnívoros del Triásico, ancestros de los crocodilomorfos. [1] Una postura de extremidades erguidas en forma de pilar es aquella en la que el fémur se articula verticalmente con el acetábulo de la cadera, que está en ángulo hacia abajo, de modo que la pierna se coloca debajo del cuerpo y actúa como un pilar de soporte de peso. Sin embargo, es probable que haya una variación significativa en la estructura de la cadera de los aetosaurios, y las extremidades delanteras pueden haber tenido una postura "híbrida" semidesparramada. Si bien la postura de las extremidades traseras es similar a la de los rauisuquios, otros rasgos son más plesiomórficos (típicos de los pseudosuquios ancestrales), como la pelvis robusta y los pies anchos de cinco dedos. [6] Las extremidades anteriores eran más pequeñas que las posteriores, y el radio en particular era mucho más corto que el húmero . Sin embargo, su forma corporal baja y pesada requiere que todos los aetosaurios fueran cuadrúpedos . Tenían múltiples adaptaciones para fortalecer el cuerpo en respuesta a su pesada armadura: el músculo iliofibular se adhirió a una posición inferior en el peroné , el cuarto trocánter del fémur se agrandó, los procesos transversales (inserciones de las costillas) se desarrollaron en pedestales largos y masivos, y las especies más grandes incluso adquirieron refuerzos hiposfeno-hipantro entre sus vértebras. [3] [4]
Aunque los aetosaurios eran generalmente reptiles de cuerpo ancho, hay alguna variación en el grado de esta tendencia. Los tipotóracos , ejemplificados por Typothorax y Paratypothorax , tenían un caparazón muy ancho, en forma de disco, bordeado por pequeñas espinas o quillas y en transición a una cola estrecha. La especie más grande de tipotóracos puede haber medido alrededor de 3 metros (9,8 pies) de largo y 110 kg (243 libras) de peso. Los desmatosuquinos ( Desmatosuchinae sensu stricto ), como Desmatosuchus y Longosuchus , tenían cuerpos moderadamente más estrechos y no tenían armadura abdominal. Sin embargo, también adquirieron una armadura posterior más espinosa, especialmente en la región cervical (cuello). [11] Desmatosuchus fue probablemente uno de los aetosaurios más grandes conocidos, con 4-6 m (13-20 pies) de largo y 280 kg (620 libras) de peso. [12] [3] [13] Los aetosaurios que no encajan en estas dos categorías, como Stagonolepis y Neoaetosauroides , generalmente tenían formas estrechas, extremidades delgadas y una restricción en el caparazón por encima de la cadera. [3] Este tipo de cuerpo es plesiomórfico (ancestral) a las otras dos formas, con algunos aetosaurios de cuerpo estrecho más estrechamente relacionados con los tipotoracidos y otros más cercanos a los desmatosuquinos. [4] Algunos géneros plesiomórficos, como el género noriano Aetosaurus y el carniano Coahomasuchus , tendían a ser pequeños, de alrededor de un metro (3,2 pies) de longitud. [14] Otros eran más grandes, como el aetosaurio más basal Aetosauroides y el desmatosuquino temprano Calyptosuchus . [3]

Los aetosaurios estaban fuertemente blindados, con filas de grandes placas óseas entrelazadas, conocidas como osteodermos , que protegían la espalda, los costados, el vientre y la cola. [6] Estos osteodermos generalmente tienen una forma cuadrangular (de cuatro lados) y, con toda seguridad, se usaban como defensa contra los depredadores. La mayoría de los osteodermos están muy picados en sus superficies superiores y lisos en sus caras inferiores. Tienen una estructura interna heterogénea: la porción interna de cada osteodermo está hecha de hueso esponjoso o canceloso (también llamado diploe ) y sus porciones externas están formadas por hueso compacto. [3] En vida, estas placas probablemente estaban cubiertas por una cubierta queratinosa (cuerno), como los escudos de los cocodrilos modernos , que son otro ejemplo de osteodermos pseudosuquios. [15] Los osteodermos son útiles para diagnosticar taxones de aetosaurios, y las especies de aetosaurios a menudo se pueden identificar a partir de escudos individuales en función de su forma, estructura o patrón de ornamentación. [16] [17] [3]
Los aetosaurios tienen cuatro filas de osteodermos que recorren su lado dorsal (espalda), formando una placa continua a menudo llamada caparazón . Las dos filas internas, que flanquean la línea media de la columna vertebral, se conocen como osteodermos paramedianos. Estos tienden a ser más anchos que largos y están fuertemente ornamentados con hoyos o surcos radiales. Casi todos los aetosaurios poseen una pequeña protuberancia o superficie elevada, conocida como eminencia dorsal, en la superficie superior de cada placa. La eminencia dorsal a menudo se encuentra posteriormente (hacia atrás) o medialmente (hacia adentro) en su respectivo osteodermo paramediano, aunque hay muchas excepciones dentro del grupo. Los osteodermos paramedianos casi siempre tienen bordes anteriores elevados o deprimidos, donde las placas se superponen con las que están frente a ellos. Si el borde anterior está elevado, el área se llama barra anterior, mientras que si está deprimido, el área se llama lámina anterior. Aunque dos filas de osteodermos paramedianos son comunes en los primeros arcosauriformes, pocos reptiles se acercan a los aetosaurios en la complejidad de sus osteodermos. Los osteodermos de los doswélidos , erpetosúquidos y ciertos crocodilomorfos se confunden o se comparan ocasionalmente con los de los aetosaurios. Tanto una barra anterior como una eminencia dorsal se encuentran en Acaenasuchus , un pariente cercano de los aetosaurios originalmente identificado erróneamente como un desmatosuquino juvenil. [8] [3] [4] [18]

Las dos filas externas de osteodermos, que se encuentran al lado de los paramedianos, se conocen como osteodermos laterales. Son paralelos a los paramedianos en casi toda la parte posterior, aunque los dos primeros paramedianos detrás de la cabeza (conocidos como osteodermos nucales) son solitarios. Las placas laterales generalmente están separadas en dos superficies, o bridas, flexionadas entre su eminencia dorsal. La brida superior o dorsal se encuentra en el mismo plano que los osteodermos paramedianos. La brida inferior/externa o lateral se envuelve hacia abajo sobre el costado del cuerpo. La eminencia dorsal entre estas bridas a menudo tiene la forma de una hoja baja, un bulto o una espiga. En los osteodermos laterales cervicales, que se colocan en el cuello, la eminencia dorsal tiende a manifestarse como una espiga prominente. [19] Esto se lleva al extremo en desmatosuchinos como Longosuchus y Desmatosuchus , donde la espiga se agranda en un cuerno muy curvado. [8]
En la mayoría de los aetosaurios (una excepción importante son los desmatosuquinos ), la parte inferior del animal también está protegida por osteodermos. Estos osteodermos ventrales (del vientre) son generalmente más pequeños y planos que la serie dorsal, y están dispuestos en un mayor número de filas (normalmente de 5 a 14 filas), al menos en el torso. Las filas de osteodermos ventrales suelen curvarse hacia fuera y separarse bajo la cadera, dejando un amplio espacio para la abertura cloacal . Grandes espinas en forma de gancho aparecen alrededor de esta abertura en Typothorax , una de las pocas excepciones a una regla general de osteodermos ventrales lisos. Las filas ventrales se dividen en una piel de pequeñas placas en el cuello y una pequeña cantidad de filas anchas debajo de la cola. Una densa variedad de placas pequeñas que no se superponen, conocidas como osteodermos apendiculares, cubrían las extremidades delanteras y traseras. [3] [20]
El material de Aetosaurio fue descrito por primera vez por el paleontólogo suizo Louis Agassiz en 1844. Nombró al género Stagonolepis de la arenisca de Lossiemouth en Elgin , Escocia , pero consideró que el fósil era un pez devónico en lugar de un reptil triásico. Esto puede deberse a que consideró que los estratos eran parte de la arenisca roja antigua y, por lo tanto, de edad paleozoica . Agassiz confundió los osteodermos con grandes escamas romboidales, que pensó que estaban dispuestas en un patrón similar a las de los pejelagartos . También pensó que estas supuestas escamas eran muy similares a las del pez de aletas lobuladas Megalichthys debido a su gran tamaño. [21]
El biólogo inglés Thomas Henry Huxley reconsideró las escamas de pez descritas por Agassiz y consideró que pertenecían a un cocodrilo. Propuso esto por primera vez a la Sociedad Geológica de Londres en 1858, y entró en más detalles en un artículo de 1875 en la revista trimestral de la sociedad. En ese momento, se había descubierto nuevo material de Elgin que indicaba que Stagonolepis no era un pez, sino un reptil. Sin embargo, Stagonolepis todavía era conocido principalmente por escudos e impresiones de escudos, muchos de los cuales no estaban bien conservados. [22]

En la década de 1870 se encontraron restos más completos de aetosaurios en el Bajo Stubensandtein de Alemania . Entre ellos se encontraban esqueletos articulados completos de 22 aetosaurios. Estos especímenes se encontraron en un gran afloramiento de arenisca cerca de Stuttgart y se conservaron juntos en un área de menos de 2 metros cuadrados. Los animales probablemente fueron enterrados bajo sedimentos del lago poco después de su muerte, y el flujo de agua reposicionó sus cuerpos en el lecho del lago y los puso muy cerca unos de otros. En 1877, el paleontólogo alemán Oscar Fraas asignó estos especímenes al género recién erigido Aetosaurus . Fraas nombró al género por la semejanza del cráneo con la cabeza de un águila, con un cráneo estrecho y alargado y un hocico puntiagudo. [23]
Los primeros aetosaurios de América del Norte también fueron descritos en la década de 1870. El paleontólogo estadounidense Edward Drinker Cope nombró a Typothorax en 1875 y a Episcoposaurus en 1877, ambos de Nuevo México . Sin embargo, Cope consideró que estos géneros eran fitosaurios , arcosaurios acuáticos similares a los cocodrilos que también eran comunes en el Triásico Tardío. Si bien se ha encontrado nuevo material atribuible a Typothorax en los últimos años en todo el suroeste de Estados Unidos, Episcoposaurus ya no se considera válido. Cope, junto con paleontólogos posteriores como Friedrich von Huene , reconoció que los restos de la especie tipo E. horridus en realidad pertenecían a Typothorax . En ese momento, Cope consideró que Aetosauria pertenecía a Rhynchocephalia , un orden de reptiles que incluye al tuátara actual . También pensó que los osteodermos estrechamente ajustados, que se pensaba que estaban fusionados a las costillas, indicaban que los aetosaurios eran una transición entre los rincocéfalos y las tortugas . [24] Otra especie, E. haplocerus , fue reasignada a Desmatosuchus en 1953. [25]

Un tercer género norteamericano llamado Stegomus fue nombrado por Othniel Charles Marsh en 1896. [26] Marsh tuvo una rivalidad de larga data con Cope que se hizo famosa en las Guerras de los Huesos de finales del siglo XIX, en las que los dos intentaron superarse entre sí en el campo y en la literatura científica. A diferencia de los aetosaurios de Cope, Stegomus fue encontrado en el este de los Estados Unidos en Connecticut . Marsh también reconoció a Stegomus como un aetosaurio en lugar de un fitosaurio en su descripción inicial del género. Al igual que Cope, muchos paleontólogos tendieron a considerar que los escudos de los aetosaurios pertenecían a los fitosaurios durante este período de tiempo. [17] Marsh consideró que los aetosaurios estaban estrechamente relacionados con los dinosaurios basándose en sus metatarsianos alargados (huesos del pie). [24]
Como grupo diferenciado, Aetosauria fue nombrado en 1889 por el naturalista inglés Richard Lydekker y el zoólogo Henry Alleyne Nicholson . [24] Consideraron que Aetosauria era uno de los tres subórdenes del orden Crocodilia, siendo los otros dos Parasuchia (un grupo que incluye a los fitosaurios y otras formas del Triásico) y Eusuchia (un grupo que incluye a todos los crocodilomorfos post-Triásico). Nicholson y Lydekker colocaron una sola familia dentro del suborden, Aëtosauridæ. Consideraron que los aetosaurios eran similares a los crocodilianos actuales a pesar de sus metatarsianos más largos. [24]
El material de Aetosaurio continuó siendo descrito hasta principios del siglo XX, con paleontólogos notables como Barnum Brown y Charles Camp recolectando especímenes. [27] Los mejores restos de aetosaurios americanos ayudaron a establecer información anatómica para algunas especies comunes, como Desmatosuchus spurensis (descrito en detalle por EC Case en 1922) y "Typothorax" meadei (ahora Longosuchus , descrito por HJ Sawin en 1947). [28] Sin embargo, los restos de aetosaurios todavía se confundían con los de fitosaurios, y todavía se consideraban "parasuquios" predominantemente carnívoros y semiacuáticos. La redescripción de Stagonolepis robertsoni de AD Walker en 1961 ayudó a desviar estas nociones y estableció una visión más moderna de la anatomía y ecología de los aetosaurios. [29]
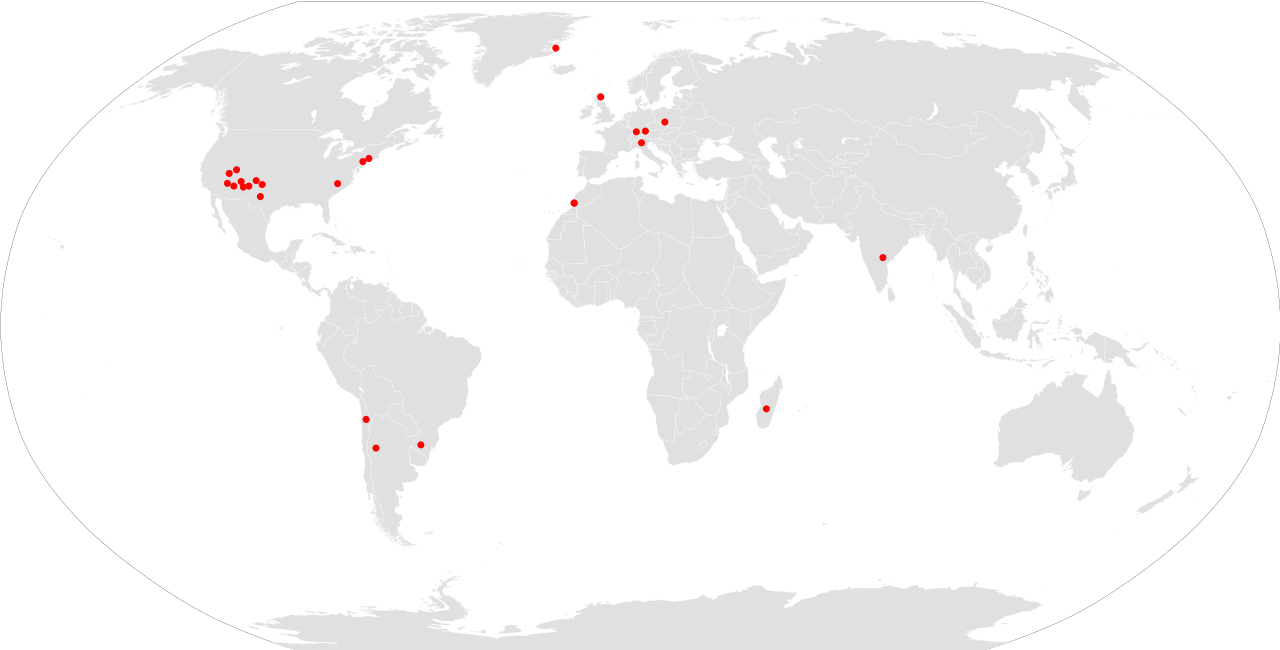
Se han encontrado fósiles de Aetosaurio en todos los continentes excepto Australia y la Antártida , lo que les da una distribución casi mundial durante el Triásico Tardío. Los restos de Aetosaurio son más abundantes en los continentes del norte ( Laurasiático ), y los fósiles son particularmente comunes en la Formación Chinle y el Grupo Dockum del suroeste de los Estados Unidos . La mayoría de los fósiles se han encontrado en Arizona , Nuevo México y Texas . [30] También se han encontrado algunos restos en Wyoming , Colorado y Utah , particularmente en el Parque Nacional Canyonlands y el Parque Nacional Zion . [31] También se conocen Aetosaurios del Supergrupo Newark a lo largo de la costa este de los Estados Unidos, en estados como Connecticut y Carolina del Norte . [32] Aetosaurus y Paratypothorax se han recuperado de la Formación Fleming Fjord de Jameson Land , en la costa este de Groenlandia . [33] Estos dos géneros fueron nombrados originalmente a partir del Stubensandstein Inferior de Alemania. Escocia , Polonia e Italia también han producido fósiles de aetosaurios. Se conocen algunos aetosaurios del norte de África, con escudos que se han encontrado en la Formación Timesgadiouine del Carniense tardío en Marruecos. [34] [35] Durante el Triásico Tardío, Marruecos habría estado en estrecha proximidad con el Supergrupo Newark de América del Norte en el supercontinente de Pangea . También es posible que Desmatosuchus estuviera presente en África, ya que los fósiles de la Serie Zarzaitina en Argelia se han referido al género. [14]

Los aetosaurios también se encuentran en áreas correspondientes a Gondwana (sur de Pangea), aunque son considerablemente menos comunes o diversos que en los continentes del norte. Se conocen aetosaurios sudamericanos de Argentina , Brasil y Chile . En Argentina, Aetosauroides y Neoaetosauroides provienen de la Formación Ischigualasto del Carniano y la Formación Los Colorados del Noriano , respectivamente. [36] [1] En Brasil, se han encontrado fósiles en las Formaciones Santa Maria y Caturrita en Rio Grande do Sul (Paleorrota). [14] [36] Los aetosaurios chilenos están representados por un género fragmentario, Chilenosuchus , de la Región de Antofagasta . [37] También se han encontrado aetosaurios en la India , que, junto con Sudamérica, fue parte de Gondwana durante el Triásico Tardío. Los primeros relatos sobre los aetosaurios indios se basaban en material de la Formación Maleri en el centro-sur de la India, aunque la mayoría de estos restos eran demasiado inadecuados para asignar especímenes a un género en particular. Según las descripciones publicadas, los fósiles de aetosaurios indios se parecen más a Longosuchus y Paratypothorax, que son considerablemente más especializados que la mayoría de los aetosaurios de Gondwana descritos. [38] En 2023, un nuevo aetosaurio de la Formación Dharmaram Inferior de la India fue nombrado Venkatasuchus armatus , un aetosaurio tipotorácico . [39] Los informes de aetosaurios de Madagascar [40] [41] se basan en probables escudos de crocodilomorfos . [42] Las supuestas huellas de aetosaurios y material esquelético de Sudáfrica tampoco están corroborados. [43]
Las huellas pertenecientes al icnogénero Brachychirotherium se asocian a menudo con aetosaurios. [44] [45] Se conocen huellas de Brachychirotherium en Rio Grande do Sul en Paleorrota, Brasil [36] así como en Italia , [46] Alemania , [47] el este de los Estados Unidos, [48] y muchos otros lugares con aetosaurios. También son comunes en el suroeste de los Estados Unidos, habiéndose encontrado en el Parque Nacional Canyonlands y el Monumento Nacional de los Dinosaurios . [31] Muchas de estas huellas tienen un ancho de vía estrecho (lo que significa que las huellas izquierda y derecha están colocadas muy juntas) y casi se sobreponen una a la otra. Un análisis funcional de 2011 del esqueleto de Typothorax coccinarum indicó que tenía el rango de movimiento necesario para producir las huellas. [49]
Los aetosaurios pertenecen a Pseudosuchia , un clado de arcosaurios que incluye a los crocodilianos actuales y se caracteriza por la estructura distintiva de los huesos del tobillo . [ dudoso – discutir ] Los aetosaurios eran tradicionalmente referidos a un grupo (ahora obsoleto) llamado tecodontos , que incluía a todos los parientes " primitivos " de los crocodilianos que vivieron en el Triásico. Con el surgimiento de la filogenética , los aetosaurios fueron posteriormente colocados en un grupo llamado Suchia , que incluía a muchos crurotarsos del Triásico, así como a crurotarsos posteriores, incluidos los crocodilianos. [50]
Originalmente, todos los aetosaurios eran considerados miembros de la familia Stagonolepididae. Los primeros análisis filogenéticos dividieron a los aetosaurios en dos subfamilias, Aetosaurinae y Desmatosuchinae. Los aetosaurios se caracterizan por proyecciones llamadas eminencias en los osteodermos paramedianos dorsales que están cerca de la línea media de la espalda. Los desmatosuquinos tienen algunas características distintivas más, incluyendo surcos en los paramedianos dorsales que los ayudan a trabarse con las placas laterales en una articulación apretada. Muchos desmatosuquinos tienen largas púas que proyectan desde las placas laterales. Estas púas son especialmente prominentes en Desmatosuchus . Los aetosaurios, por otro lado, tienden a tener menos púas. Muchos aetosaurios, como Aetosaurus y Neoaetosauroides , tienen caparazones lisos y carecen de púas por completo. [8] Estudios más recientes (ver abajo) han favorecido un tercer grupo, Typothoracinae, que al igual que Desmatosuchinae tiene espinas largas, pero se diferencia en que tiene articulaciones más anguladas entre los osteodermos. Además, el género Aetosauroides ahora se clasifica a menudo fuera de Stagonolepididae como un aetosaurio no estagonolepídido, lo que hace que los nombres Aetosauria y Stagonolepididae ya no sean sinónimos. [51]
La filogenia de los aetosaurios fue investigada por primera vez en 1994 por el paleontólogo J. Michael Parrish. Aetosauroides , Aetosaurus , Desmatosuchus , Longosuchus , Neoaetosauroides , Stagonolepis y Typothorax se incluyeron en el análisis filogenético. Se descubrió que los aetosaurios formaban un clado con los rauisuquios, al que Parrish denominó Rauisuchiformes. Rauisuchiformes también incluía el superorden Crocodylomorpha , al que pertenecen los crocodilianos actuales . Parrish descubrió que Aetosauria era un grupo monofilético y, por lo tanto, un clado verdadero que consistía en un ancestro común de los aetosaurios y todos sus descendientes. Para definir filogenéticamente a Aetosauria, Parrish identificó cinco sinapomorfías o características compartidas. La primera sinapomorfia se refería a la mandíbula, con el premaxilar en su punta edéntulo (sin dientes), vuelto hacia arriba y ancho para formar una "pala". Además, el hueso dentario en la mandíbula inferior también es desdentado, vuelto hacia arriba y ancho. El tamaño reducido y la forma cónica simple de los dientes se consideraron otra sinapomorfia. Dos sinapomorfias más de los aetosaurios son compartidas con los crocodilomorfos, pero no se consideraron una indicación de una relación filogenética cercana; el cuerpo está cubierto de una armadura dorsal y ventral para formar un caparazón completo, y los osteodermos paramedianos son mucho más anchos que largos, con hoyuelos distintivos. Una sinapomorfia final se encontró en la estructura de los huesos de las extremidades. En todos los aetosaurios, las extremidades son muy robustas, con grandes inserciones musculares como la cresta deltopectoral del húmero , el cuarto trocánter del fémur , la cresta intracondílea de la tibia y el trocánter iliofibular del peroné . [52]
En el análisis filogenético de Parrish, se encontró que Aetosaurus era el miembro más basal del clado, el primero en divergir después del ancestro común más reciente . Después de Aetosaurus , hay una politomía de tres clados más pequeños en los que se desconoce qué clado divergió primero del grupo. Dentro de esta politomía estaba Neoaetosauroides , un clado que contenía a Aetosauroides y Stagonolepis , y otra politomía que incluía a Longosuchus , Desmatosuchus y un clado que contenía a Paratypothorax y Typothorax .
Un estudio posterior de los paleontólogos Andrew B. Heckert y Spencer G. Lucas en 1999 amplió el número de sinapomorfias que diagnostican Aetosauria a 18. Las nuevas sinapomorfias incluían fenestras temporales , u orificios, que se abrían en el costado del cráneo en lugar de en la parte superior, osteodermos laterales que se articulaban con los paramedianos y osteodermos que cubrían las extremidades. Se seguía descubriendo que Aetosaurus era el miembro más basal, pero la filogenia de los aetosaurios más derivados difería en que Typothorax y Paratypothorax se dividían en dos clados diferentes, siendo sus taxones hermanos Desmatosuchus y Longosuchus , respectivamente. Más importante aún, se incluyó en el análisis un nuevo aetosaurio llamado Coahomasuchus . Se descubrió que Coahomasuchus era un aetosaurio basal estrechamente relacionado con Stagonolepis , y también apareció temprano en el registro fósil de los aetosaurios. Anteriormente, los miembros basales solo se conocían de épocas posteriores, y aparecieron después de los aetosaurios más avanzados. [14]
En 2003, los paleontólogos Simon R. Harris, David J. Gower y Mark Wilkinson examinaron estudios filogenéticos previos de aetosaurios y criticaron la forma en que usaron ciertos caracteres para producir cladogramas. Llegaron a la conclusión de que solo tres hipótesis de relaciones de aetosaurios de estudios previos seguían siendo ciertas: que Aetosaurus es el aetosaurio más basal, que Aetosauroides es el taxón hermano de Stagonolepis robertsoni y que Longosuchus y Desmatosuchus están más estrechamente relacionados entre sí que con Neoaetosauroides . También corrigieron los árboles de todos los análisis previos. [53]
Más recientemente, un análisis de 2007 realizado por el paleontólogo William G. Parker resultó en un árbol más grande de la filogenética de los aetosaurios con la inclusión de Heliocanthus . Basándose en el árbol, Parker definió los clados Typothoracisinae y Paratypothoracisini , ambos dentro de Aetosaurinae. Parker también dio una definición filogenética revisada de Aetosauria, mencionando que la definición anterior, hecha por Heckert y Lucas en 2000, era algo ambigua. [8] Heckert y Lucas (2000) definieron a Aetosauria como un taxón basado en tallos , afirmando que Aetosauria incluía a todos los crurotarsos que estaban más estrechamente relacionados con Desmatosuchus que con el grupo hermano inmediato de Aetosauria. [54] Debido a que el grupo hermano inmediato de Aetosauria era incierto, Parker ofreció una nueva definición con varios géneros de crurotarsos no aetosaurios en lugar de un grupo hermano. Según Parker, Aetosauria incluía todos los taxones más estrechamente relacionados con Aetosaurus y Desmatosuchus que con Leptosuchus , Postosuchus , Prestosuchus , Poposaurus , Sphenosuchus , Alligator , Gracilisuchus y Revueltosaurus . [8]
En 2012 se nombró un nuevo género de aetosaurio, Aetobarbakinoides . El análisis filogenético de ese estudio encontró que Aetosaurinae es una agrupación parafilética . Como grupo parafilético, los aetosaurinos compartirían un ancestro común más reciente que también es el ancestro de otros aetosaurios no aetosaurinos, y por lo tanto no podrían formar su propio clado . [51] El análisis de Parker de 2007 aceptó esta definición. En 2002, Heckert y Lucas definieron a Aetosaurinae como "un taxón basado en tallos que contiene todos los taxones más estrechamente relacionados con Aetosaurus que con el último ancestro común de Aetosaurus y Desmatosuchus ". [8] El estudio de 2012 colocó a Aetosaurus en la base del clado de los estagonolepídidos, con los taxones tradicionales de aetosaurinos ubicados en posiciones sucesivamente más derivadas . En el análisis, estos taxones están en realidad más estrechamente relacionados con Desmatosuchus que con Aetosaurus . Por lo tanto, según la definición de Heckert y Lucas, Aetosaurinae podría restringirse únicamente al propio Aetosaurus .
Otro hallazgo de este estudio fue que Aetosauroides se encuentra fuera de Stagonolepididae. Si esta filogenia es correcta, Stagonolepididae y Aetosauria no serían agrupaciones equivalentes, y Aetosauroides sería el primer aetosaurio no estagonolepídido. El siguiente cladograma se simplificó después de un análisis presentado por Devin K. Hoffman, Andrew B. Heckert y Lindsay E. Zanno. [55]
En 2016, William Parker realizó un nuevo análisis filogenético de Aetosauria y propuso una hipótesis alternativa sobre las relaciones entre los aetosaurios. A continuación se muestra el cladograma: [4]

Aunque los aetosaurios son conocidos exclusivamente del Triásico Tardío, su posición actualmente aceptada en la filogenia de los arcosaurios indica que se originaron a partir de arcosaurios pseudosuquios más basales en el Triásico Temprano o Medio. Dado que los aetosaurios están altamente especializados con muchas características anatómicas que no se ven en otros pseudosuquios, los orígenes evolutivos del grupo son poco comprendidos. El reciente descubrimiento de especímenes completos del pseudosuquio del Triásico Tardío Revueltosaurus callenderi indica que puede haber sido cercano a la ascendencia de los aetosaurios. Varios análisis filogenéticos lo ubican como el taxón hermano o pariente más cercano de Aetosauria. Al igual que los aetosaurios, Revueltosaurus tiene dos filas de osteodermos paramedianos a lo largo de su espalda y, en la región de las mejillas del cráneo, un maxilar que encaja en un surco del hueso yugal . [3] Un análisis filogenético coloca a Turfanosuchus dabanensis , un pseudosuquio del Triásico Medio, como el taxón hermano de Revueltosaurus y Aetosauria, convirtiéndolo potencialmente en el " tallo " de los aetosaurios más antiguo conocido ("tallo" significa que se encuentra en la rama que incluye a los aetosaurios, pero no es en sí mismo un aetosaurio). [56] Sin embargo, otros estudios han considerado a Turfanosuchus parte de una familia diferente de pseudosuquios, los gracilisúquidos . [57]
En 2012 se describió otro "aetosaurio madre" de los lechos Manda del Triásico Medio de Tanzania. Se diferencia de otros pseudosuquios del Triásico Medio en que tiene un cráneo largo, una pequeña fenestra antorbital que encaja en una gran fosa antorbital delante de la cuenca del ojo, dientes afilados y curvados y osteodermos que cubren gran parte de su cuerpo. Al igual que los aetosaurios y Revueltosaurus , tiene un maxilar que encaja en el yugal. Revueltosaurus , Turfanosuchus y el pseudosuquio tanzano sin nombre son buenos candidatos para el ancestro hipotético de los aetosaurios porque ambos tienen filas dobles de osteodermos en forma de hoja a lo largo de sus espaldas que potencialmente podrían haber evolucionado hasta convertirse en los osteodermos paramedianos de los aetosaurios, que encajan perfectamente. [56]
En 1904, el paleontólogo estadounidense Henry Fairfield Osborn describió a los aetosaurios como animales acuáticos carnívoros del orden Parasuchia, mencionando que "[Parasuchia] constituye un orden independiente, probablemente formas de agua dulce, litorales, carnívoras, de hocico corto (Aëtosaurus) o de hocico largo (Phytosaurus, Mystriosuchus), análogas en sus hábitos a los modernos Crocodilia". [58] Los primeros restos de aetosaurios se encontraron a menudo en arcillas junto a esqueletos de animales acuáticos como fitosaurios y animales terrestres como dinosaurios y trilofosaurios . Esto puede haber llevado a algunos paleontólogos a creer que los animales habían muerto en entornos pantanosos. Debido a que en estas arcillas había una gran cantidad de esqueletos de animales que normalmente no habrían habitado pantanos, algunos paleontólogos incluso sugirieron que los aetosaurios se alimentaban de los cadáveres de animales que quedaban atrapados en los pantanos y morían. Más tarde surgieron dudas sobre este estilo de vida, ya que los dientes de los aetosaurios muestran pocos indicios de carnivoría y el peso de la armadura sugiere que los aetosaurios tenían "un modo de vida pasivo". Sin embargo, los aetosaurios todavía se consideraban parcialmente acuáticos hasta mediados del siglo XX. [28]

Los aetosaurios eran herbívoros, probablemente alimentándose de helechos y helechos con semillas que eran comunes en el Triásico. La forma hacia arriba de su hocico sugiere que los aetosaurios pueden haber desenterrado raíces y tubérculos. [59] Los aetosaurios tienen varias características anatómicas que pueden haber sido adaptaciones para excavar, incluido un radio corto en relación con el húmero (observado en muchos otros tetrápodos excavadores) y una gran cresta deltopectoral en el húmero que sirvió como unión para los músculos. Los aetosaurios también tienen grandes patas traseras, o pes , con grandes garras que probablemente usaban para cavar. Un aetosaurio, Typothorax , tiene un entepicóndilo en el húmero, que es el origen de los músculos pronadores del antebrazo y flexores manuales que a menudo se usan para cavar. Además, tiene un gran proceso olécranon en el cúbito que se proyecta hacia atrás más allá del codo, lo que proporciona una gran área para la inserción del músculo tríceps . [45] Si bien muchos estudios han sugerido que los aetosaurios tenían un estilo de vida fosorial o de excavación, [52] los aetosaurios tienen pocas de las características que tienen los animales fosoriales como adaptaciones a la excavación. Por lo tanto, es probable que los aetosaurios pudieran excavar hasta cierto punto, posiblemente buscando comida, pero no podían excavar. [45]
Aunque las características de las extremidades indican que los aetosaurios probablemente cavaban para alimentarse, las características del cráneo y los dientes pueden indicar qué tipo de comida comían. Los aetosaurios tienen muchas características derivadas que no se ven en otros crurotarsos, lo que indica que están adaptados a una dieta diferente. A diferencia de los dientes afilados y curvados de otros arcosaurios triásicos, los aetosaurios tenían dientes simples y cónicos. Las puntas de las mandíbulas eran edéntulas , o sin dientes, y probablemente sostenían un pico. Los dientes tienen muy poco desgaste, lo que sugiere que los aetosaurios no consumían material vegetal rígido y duro. Es más probable que consumieran vegetación no abrasiva como hojas blandas. [52]
Se han propuesto teorías alternativas sobre la dieta de los aetosaurios. En 1947, H. J. Sawin propuso que el aetosaurio Longosuchus era un carroñero basándose en la proximidad de algunos especímenes a una gran cantidad de esqueletos que probablemente eran cadáveres. [28] Un estudio de 2009 sobre la biomecánica de la mandíbula del género sudamericano Neoaetosauroides sugirió que el animal podría haberse alimentado de larvas e insectos sin exoesqueletos duros. Esto se debe a que Neoaetosauroides carece de dentados o facetas de desgaste en los dientes y tiene una palanca de mandíbula que no está diseñada para fuerzas fuertes como aplastar y cortar. El estudio reconoció que los aetosaurios del norte como Desmatosuchus y Stagonolepis sí tenían mandíbulas que habrían soportado una musculatura fuerte y probablemente estaban mejor adaptadas para comer material vegetal. [60] [61]

Al menos algunos aetosaurios construían nidos y protegían sus huevos. [62]
En 1996, el geólogo Stephen Hasiotis descubrió fosas fosilizadas de 220 millones de años de antigüedad en el Bosque Petrificado de Arizona, en parte de la Formación Chinle , que se supone son nidos de aetosaurios y fitosaurios . Los "nidos" están compactados y parecen muy similares a los nidos de los cocodrilos actuales que cuidan sus nidos. [59] Sin embargo, parece que estos "nidos" son en realidad el resultado de la erosión de la arenisca. [63]
Se conoce un segundo posible sitio de anidación de aetosaurios en el noreste de Italia. Los nidos se conservan como depresiones en rocas carbonatadas que tienen forma circular o de herradura, con altas crestas alrededor de los lados. Parecen ser inusualmente complejos para nidos creados por reptiles del Triásico. Se encontraron huellas de arcosaurios cerca que se parecían a las de los aetosaurios, aunque no estaban presentes en la misma capa. Debido a que las huellas se encontraron tan cerca de los nidos, es probable que los aetosaurios los construyeran. [62]
La edad de los aetosaurios individuales se puede determinar examinando sus osteodermos. Se ha afirmado que algunos osteodermos aislados pertenecen a aetosaurios juveniles basándose en su tamaño y forma, pero estas hipótesis a menudo han sido cuestionadas. Por ejemplo, los osteodermos juveniles de Calyptosuchus fueron identificados más tarde como los del pseudosuquio de cuerpo pequeño Revueltosaurus (que no es un aetosaurio), y los osteodermos juveniles de Desmatosuchus han sido reinterpretados como los del aetosaurio Acaenasuchus , que tenía un tamaño corporal relativamente pequeño en la madurez. Los estudios de la estructura ósea de los osteodermos paramedianos indican que se depositó hueso nuevo a lo largo de los bordes de cada placa a lo largo de la vida de un aetosaurio. Esto significa que las líneas de crecimiento detenido en las caras inferiores de los osteodermos paramedianos se pueden utilizar para determinar la edad de un individuo. [3] Comparar las edades de los especímenes individuales con sus longitudes corporales totales indica que los aetosaurios aumentaron en longitud a tasas relativamente constantes, pero aumentaron en masa corporal a diferentes tasas dependiendo de si tenían cuerpos anchos como Typothorax o cuerpos estrechos como Aetobarbakinoides . Los aetosaurios también parecen haber crecido más lentamente que los cocodrilos modernos. [64] El análisis de los huesos de las extremidades de los aetosaurios indica que crecieron rápidamente cuando eran jóvenes y más lentamente cuando eran adultos. Este patrón de crecimiento se observa en la mayoría de los otros pseudosuquios. [3]
Debido a que las especies de aetosaurios suelen tener rangos fósiles restringidos y son abundantes en los estratos en los que se encuentran, son útiles en biocronología . Los osteodermos son los restos más comunes asociados con los aetosaurios, por lo que un único escudo identificable puede datar con precisión la capa en la que se encuentra.
Un aetosaurio, Typothorax coccinarum , se ha utilizado para definir el faunacrono de vertebrados terrestres Revueltiense . Un faunacrono de vertebrados terrestres (LVF) es un intervalo de tiempo que se define por el dato de primera aparición (FAD), o primera aparición, de un fósil índice de tetrápodo y se utiliza comúnmente para fechar estratos terrestres del Triásico Tardío y Jurásico Temprano . [67] [68] Dado que el FAD de T. coccinarum está al comienzo de la etapa del Noriano , el LVF Revueltiense comienza a principios del Noriano hace unos 216 millones de años . El Revueltiense termina con el siguiente FAD, que resulta ser el del fitosaurio Redondasaurus y el comienzo del LVF Apacheano . [2] [69]
Se han desarrollado biocronos para géneros de aetosaurios para datar estratos en el Grupo Chinle del suroeste de los Estados Unidos. Se conocen hasta 13 géneros de aetosaurios del Grupo Chinle, y la mayoría se encuentran en múltiples localidades y en períodos de tiempo cortos. En 1996, los paleontólogos Spencer G. Lucas y Andrew B. Heckert reconocieron cinco biocronos basados en la presencia de aetosaurios en todo el Grupo Chinle. [70] El número de biocronos aumentó a 11 en un estudio de 2007 realizado por Heckert y Lucas junto con Adrian P. Hunt y Justin A. Spielmann. Estos biocronos se dieron desde el LVF Otischalkiense hasta el LVF Apacheense e incluyeron géneros como Longosuchus , Tecovasuchus y Typothorax . [66]
En 2007, los paleontólogos del Museo de Historia Natural y Ciencia de Nuevo México en Albuquerque, Nuevo México, fueron acusados de plagio en algunos de sus artículos publicados que trataban sobre aetosaurios. En diciembre de 2006, el género Rioarribasuchus fue erigido como un nombre de reemplazo para "Desmatosuchus" chamaensis en el boletín del museo. [71] Sin embargo, cuatro años antes, el paleontólogo William Parker reasignó "D." chamaensis al género recién nombrado Heliocanthus en una tesis inédita, que fue ampliamente difundida entre los investigadores de aetosaurios. [72] Debido a que el nombre no fue publicado formalmente, se consideró un nomen nudum hasta enero de 2007, cuando la descripción de Parker de Heliocanthus fue publicada en el Journal of Systematic Palaeontology . [8] Los autores del artículo de 2006, Spencer G. Lucas, Adrian P. Hunt y Justin A. Spielmann, fueron acusados de "robo intelectual" por los paleontólogos Jeff Martz, Mike Taylor , Matt Wedel y Darren Naish , quienes afirmaron que Lucas et al. sabían que Parker eventualmente redescribiría la especie y erigirá formalmente un nuevo género. Según Martz, Taylor, Wedel y Naish, los autores se apresuraron a publicar su propio nombre antes de que Parker pudiera publicar el suyo. [73] En ese momento, Lucas, Hunt y Spielmann estaban en el consejo editorial del boletín del NMMNHS, mientras que Hunt también era el director del museo.
Una controversia coincidente ocurrió después de que Spielmann, Hunt y Lucas publicaran un artículo en 2006 mencionando que el holotipo de Redondasuchus no era un paramediano izquierdo, sino uno derecho. [74] En 2002, Jeff Martz llegó a la misma conclusión en una tesis inédita. [75] Él, junto con Taylor, Wedel y Naish, afirmó que esta era otra forma de plagio, ya que la tesis de Martz de 2002 fue citada por Spielmann et al. (2006), incluso si su conclusión sobre Redondasuchus no fue mencionada. [73]
Las acusaciones fueron rechazadas en una reunión interna del NMMNHS, informada por Lucas en 2008. [76] [77] [78] También fueron llevadas a la atención de una parte independiente, el Comité de Educación Ética de la Sociedad de Paleontología de Vertebrados (SVP) en 2007, y se dio una respuesta en 2008. Con respecto a Redondasuchus , la SVP no encontró evidencia explícita de plagio. En el caso de Heliocanthus y Rioarribasuchus , la SVP no intentó resolver el problema, ya que Lucas et al. y Parker ofrecieron relatos contradictorios sobre la comunicación y la intención. La respuesta de la SVP concluyó con una actualización de su política ética y recomendaciones sobre cómo se podrían manejar mejor controversias similares en el futuro. [79] [80] [81] Toda la controversia llegó a conocerse como "Aetogate", en referencia al famoso escándalo Watergate de la década de 1970. [73] Recibió amplia atención de los periódicos locales de Albuquerque y de los blogs científicos. [82] También fue el foco de un artículo de noticias en un número de 2008 de la revista Nature . [83]
{{cite journal}}: Requiere citar revista |journal=( ayuda )