El desarrollo embrionario humano o embriogénesis humana es el desarrollo y la formación del embrión humano . Se caracteriza por los procesos de división celular y diferenciación celular del embrión que ocurren durante las primeras etapas del desarrollo. En términos biológicos, el desarrollo del cuerpo humano implica el crecimiento desde un cigoto unicelular hasta un ser humano adulto . La fertilización ocurre cuando el espermatozoide ingresa y se fusiona con éxito con un óvulo . El material genético del espermatozoide y el óvulo se combinan para formar el cigoto unicelular y comienza la etapa germinal del desarrollo. El desarrollo embrionario humano cubre las primeras ocho semanas de desarrollo, que tienen 23 etapas, llamadas etapas de Carnegie . Al comienzo de la novena semana, el embrión se denomina feto (escrito "foetus" en inglés británico ). En comparación con el embrión, el feto tiene características externas más reconocibles y un conjunto más completo de órganos en desarrollo.
La embriología humana es el estudio de este desarrollo durante las primeras ocho semanas después de la fecundación. El período normal de gestación (embarazo) es de aproximadamente nueve meses o 40 semanas.
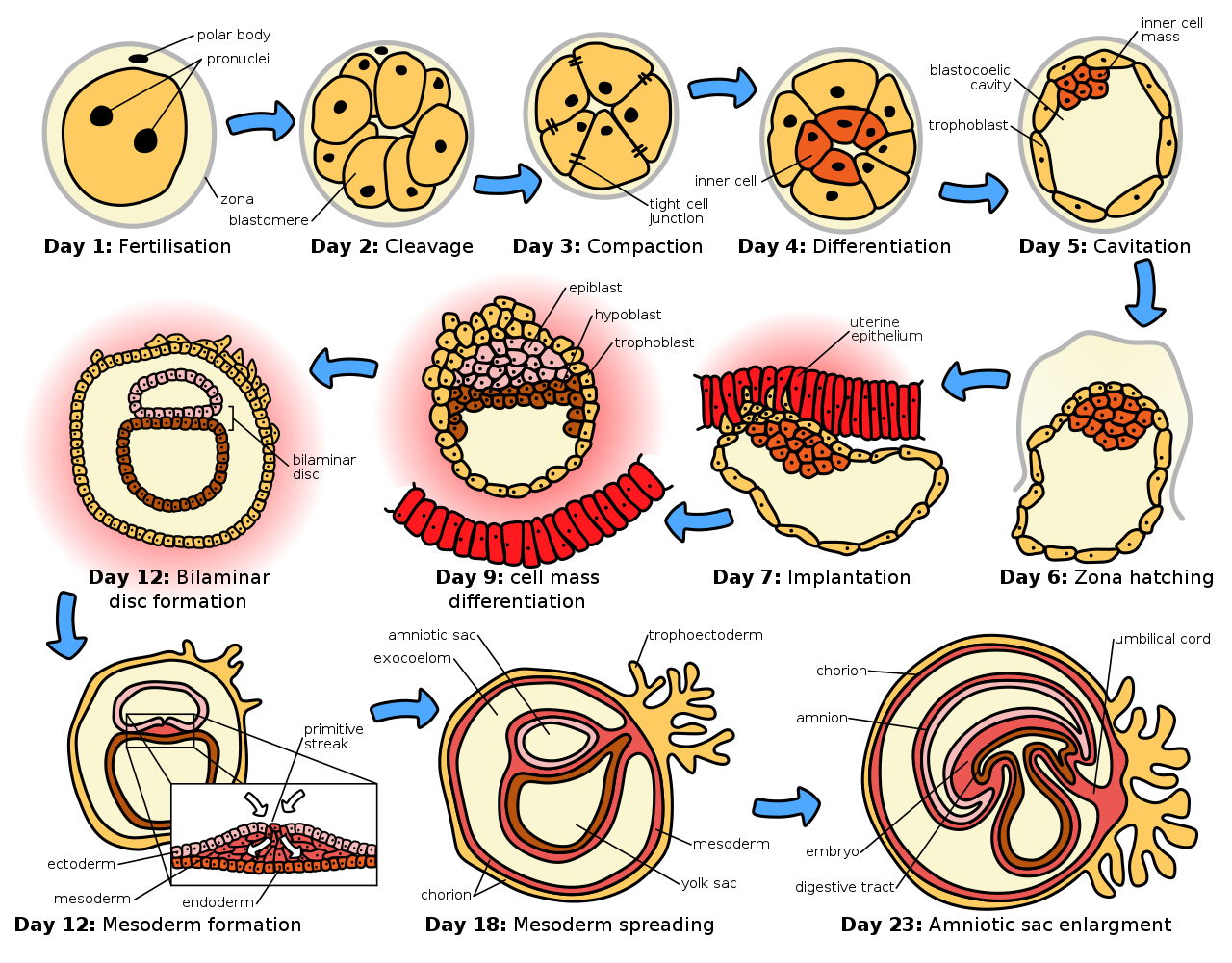
La etapa germinal se refiere al tiempo que transcurre desde la fertilización, pasando por el desarrollo del embrión temprano, hasta que se completa la implantación en el útero . La etapa germinal dura alrededor de 10 días. [1] Durante esta etapa, el cigoto se divide en un proceso llamado segmentación . Luego se forma un blastocisto que se implanta en el útero . La embriogénesis continúa con la siguiente etapa de gastrulación , cuando se forman las tres capas germinales del embrión en un proceso llamado histogénesis , y siguen los procesos de neurulación y organogénesis .
Todo el proceso de embriogénesis implica cambios espaciales y temporales coordinados en la expresión génica , el crecimiento celular y la diferenciación celular . Un proceso casi idéntico ocurre en otras especies, especialmente entre los cordados .
La fecundación se produce cuando el espermatozoide ha penetrado con éxito en el óvulo y los dos conjuntos de material genético que llevan los gametos se fusionan, dando lugar al cigoto (una única célula diploide ). Esto suele tener lugar en la ampolla de una de las trompas de Falopio . El cigoto contiene el material genético combinado que llevan tanto los gametos masculinos como femeninos, que consiste en los 23 cromosomas del núcleo del óvulo y los 23 cromosomas del núcleo del espermatozoide. Los 46 cromosomas sufren cambios antes de la división mitótica que conduce a la formación del embrión con dos células.
La fertilización exitosa es posible gracias a tres procesos que también actúan como controles para garantizar la especificidad de la especie. El primero es el de la quimiotaxis , que dirige el movimiento del espermatozoide hacia el óvulo. [2] En segundo lugar, se produce una compatibilidad adhesiva entre el espermatozoide y el óvulo. Con el espermatozoide adherido al óvulo, tiene lugar el tercer proceso de reacción acrosómica ; la parte frontal de la cabeza del espermatozoide está cubierta por un acrosoma que contiene enzimas digestivas para descomponer la zona pelúcida y permitir su entrada. [3] La entrada del espermatozoide hace que se libere calcio que bloquea la entrada a otros espermatozoides. [3] En el óvulo tiene lugar una reacción paralela llamada reacción de la zona . En ella se liberan gránulos corticales que liberan enzimas que digieren las proteínas receptoras de los espermatozoides, evitando así la poliespermia . [4] Los gránulos también se fusionan con la membrana plasmática y modifican la zona pelúcida de tal manera que impiden la entrada de más espermatozoides.

El proceso de segmentación comienza cuando el cigoto se divide por mitosis en dos células. Esta mitosis continúa y las dos primeras células se dividen en cuatro células, luego en ocho células y así sucesivamente. Cada división dura entre 12 y 24 horas. El cigoto es grande en comparación con cualquier otra célula y sufre la segmentación sin ningún aumento general de tamaño. Esto significa que con cada subdivisión sucesiva, la proporción de material nuclear y citoplasmático aumenta. [5]
Inicialmente, las células en división, llamadas blastómeros ( blastos, en griego significa brote), no están diferenciadas y se agrupan en una esfera encerrada dentro de la zona pelúcida del óvulo. Cuando se han formado ocho blastómeros , comienzan a compactarse . [6] Comienzan a desarrollar uniones en hendidura , lo que les permite desarrollarse de manera integrada y coordinar su respuesta a las señales fisiológicas y las señales ambientales. [7]
Cuando el número de células es aproximadamente dieciséis, la esfera sólida de células dentro de la zona pelúcida se denomina mórula . [ 8]

La segmentación en sí es la primera etapa de la blastulación , el proceso de formación del blastocisto . Las células se diferencian en una capa externa de células llamada trofoblasto y una masa celular interna . Con una mayor compactación, los blastómeros externos individuales, los trofoblastos, se vuelven indistinguibles. Todavía están encerrados dentro de la zona pelúcida . Esta compactación sirve para hacer que la estructura sea hermética, conteniendo el líquido que las células secretarán más tarde. La masa interna de células se diferencia para convertirse en embrioblastos y se polarizan en un extremo. Se cierran entre sí y forman uniones en hendidura , que facilitan la comunicación celular. Esta polarización deja una cavidad, el blastocele , creando una estructura que ahora se denomina blastocisto. (En animales distintos de los mamíferos, esto se llama blástula ) .
Los trofoblastos secretan líquido en el blastocele. El aumento de tamaño resultante del blastocisto hace que éste eclosione a través de la zona pelúcida, que luego se desintegra. [5] Este proceso se llama eclosión de la zona y tiene lugar el sexto día del desarrollo embrionario, inmediatamente antes del proceso de implantación. La eclosión del embrión humano está apoyada por proteasas secretadas por las células del blastocisto, que digieren proteínas de la zona pelúcida, dando lugar a un agujero. Luego, debido a la expansión y contracciones rítmicas del blastocisto, se produce un aumento de la presión dentro del propio blastocisto, el agujero se expande y finalmente el blastocisto puede emerger de esta envoltura rígida.
La masa celular interna dará lugar al preembrión , [9] el amnios , el saco vitelino y el alantoides , mientras que la parte fetal de la placenta se formará a partir de la capa externa del trofoblasto. El embrión más sus membranas se llama concepto , y en esta etapa el concepto ha llegado al útero . La zona pelúcida finalmente desaparece por completo, y las células ahora expuestas del trofoblasto permiten que el blastocisto se adhiera al endometrio , donde se implantará . La formación del hipoblasto y el epiblasto , que son las dos capas principales del disco germinativo bilaminar, ocurre al comienzo de la segunda semana. [10] Tanto el embrioblasto como el trofoblasto se convertirán en dos subcapas. [11] Las células internas se convertirán en la capa de hipoblasto, que rodeará a la otra capa, llamada epiblasto, y estas capas formarán el disco embrionario que se desarrollará en el embrión. [10] [11]
El trofoblasto también desarrollará dos subcapas: el citotrofoblasto , que está delante del sinciciotrofoblasto , que a su vez se encuentra dentro del endometrio . [10] A continuación, aparecerá otra capa llamada membrana exocelómica o membrana de Heuser que rodeará al citotrofoblasto, así como al saco vitelino primitivo. [11] El sinciciotrofoblasto crecerá y entrará en una fase llamada estadio lacunar, en la que aparecerán algunas vacuolas que se llenarán de sangre en los días siguientes. [10] [11] El desarrollo del saco vitelino se inicia con las células planas hipoblásticas que forman la membrana exocelómica, que recubrirá la parte interna del citotrofoblasto para formar el saco vitelino primitivo. Una erosión del revestimiento endotelial de los capilares maternos por las células sinciciotrofoblásticas da como resultado la formación de los sinusoides maternos desde donde la sangre comenzará a penetrar y fluir hacia y a través de las lagunas trofoblásticas para dar lugar a la circulación úteroplacentaria. [12] [13] Posteriormente, nuevas células derivadas del saco vitelino se establecerán entre el trofoblasto y la membrana exocelómica y darán lugar al mesodermo extraembrionario , que formará la cavidad coriónica . [11]
Al final de la segunda semana de desarrollo, algunas células del trofoblasto penetran y forman columnas redondeadas en el sinciciotrofoblasto. Estas columnas se conocen como vellosidades primarias . Al mismo tiempo, otras células migratorias forman en la cavidad exocelómica una nueva cavidad denominada saco vitelino secundario o definitivo, más pequeño que el saco vitelino primitivo. [11] [12]

Después de la ovulación , el revestimiento endometrial se transforma en un revestimiento secretor en preparación para aceptar el embrión. Se engrosa, sus glándulas secretoras se alargan y es cada vez más vascular . Este revestimiento de la cavidad uterina (o matriz) ahora se conoce como decidua , y produce una gran cantidad de células deciduales grandes en su tejido interglandular aumentado. Los blastómeros en el blastocisto se organizan en una capa externa llamada trofoblasto . Luego, el trofoblasto se diferencia en una capa interna, el citotrofoblasto , y una capa externa, el sinciciotrofoblasto . El citotrofoblasto contiene células epiteliales cuboidales y es la fuente de las células en división , y el sinciciotrofoblasto es una capa sincitial sin límites celulares.
El sinciciotrofoblasto implanta el blastocisto en el epitelio decidual mediante proyecciones de vellosidades coriónicas , formando la parte embrionaria de la placenta. La placenta se desarrolla una vez que se implanta el blastocisto, conectando el embrión a la pared uterina. La decidua aquí se denomina decidua basal; se encuentra entre el blastocisto y el miometrio y forma la parte materna de la placenta . La implantación es asistida por enzimas hidrolíticas que erosionan el epitelio . El sinciciotrofoblasto también produce gonadotropina coriónica humana , una hormona que estimula la liberación de progesterona del cuerpo lúteo . La progesterona enriquece el útero con un revestimiento grueso de vasos sanguíneos y capilares para que pueda oxigenar y sostener al embrión en desarrollo. El útero libera azúcar del glucógeno almacenado de sus células para nutrir al embrión . [14] Las vellosidades comienzan a ramificarse y contienen vasos sanguíneos del embrión. Otras vellosidades, llamadas vellosidades terminales o libres, intercambian nutrientes. El embrión está unido a la cubierta trofoblástica por un tallo de conexión estrecho que se desarrolla en el cordón umbilical para unir la placenta al embrión. [11] [15] Las arterias en la decidua se remodelan para aumentar el flujo sanguíneo materno hacia los espacios intervellosos de la placenta, lo que permite el intercambio de gases y la transferencia de nutrientes al embrión. Los productos de desecho del embrión se difundirán a través de la placenta.
A medida que el sinciciotrofoblasto comienza a penetrar la pared uterina, también se desarrolla la masa celular interna (embrioblasto). La masa celular interna es la fuente de las células madre embrionarias , que son pluripotentes y pueden convertirse en cualquiera de las tres células de la capa germinal, y que tienen la capacidad de dar origen a todos los tejidos y órganos.
El embrioblasto forma un disco embrionario de dos capas, la capa superior se llama epiblasto y la capa inferior, hipoblasto . El disco se estira entre lo que se convertirá en la cavidad amniótica y el saco vitelino. El epiblasto está adyacente al trofoblasto y está formado por células columnares; el hipoblasto es el más cercano a la cavidad del blastocisto y está formado por células cuboidales. El epiblasto migra desde el trofoblasto hacia abajo, formando la cavidad amniótica, cuyo revestimiento se forma a partir de amnioblastos desarrollados a partir del epiblasto. El hipoblasto es empujado hacia abajo y forma el revestimiento del saco vitelino (cavidad exocelómica). Algunas células hipoblásticas migran a lo largo del revestimiento interno del citotrofoblasto del blastocele, secretando una matriz extracelular en el camino. Estas células hipoblásticas y la matriz extracelular se denominan membrana de Heuser (o membrana exocelómica), y cubren el blastocele para formar el saco vitelino (o cavidad exocelómica). Las células del hipoblasto migran a lo largo de los bordes externos de este retículo y forman el mesodermo extraembrionario; esto altera el retículo extraembrionario. Pronto se forman bolsas en el retículo, que finalmente se fusionan para formar la cavidad coriónica (celoma extraembrionario).



Aparece la línea primitiva , una colección lineal de células formadas por el epiblasto migratorio, y esto marca el comienzo de la gastrulación , que tiene lugar alrededor del decimoséptimo día (semana 3) después de la fertilización. El proceso de gastrulación reorganiza el embrión de dos capas en un embrión de tres capas, y también le da al embrión su orientación específica de cabeza a cola y de adelante hacia atrás, a través de la línea primitiva que establece la simetría bilateral . Un nódulo primitivo (o nudo primitivo) se forma frente a la línea primitiva que es el organizador de la neurulación . Un hoyo primitivo se forma como una depresión en el centro del nódulo primitivo que se conecta con la notocorda que se encuentra directamente debajo. El nódulo ha surgido de los epiblastos del piso de la cavidad amniótica, y es este nódulo el que induce la formación de la placa neural que sirve como base para el sistema nervioso.
La placa neural se formará opuesta a la línea primitiva a partir del tejido ectodérmico, que se engrosa y se aplana formando la placa neural. El epiblasto de esa región se desplaza hacia abajo hasta la línea en la ubicación del surco primitivo, donde se produce el proceso llamado ingresión , que conduce a la formación del mesodermo. Esta ingresión hace que las células del epiblasto se desplacen hacia la línea primitiva en una transición epitelial-mesenquimal ; las células epiteliales se convierten en células madre mesenquimales, células estromales multipotentes que pueden diferenciarse en varios tipos de células. El hipoblasto es empujado fuera del camino y continúa formando el amnios . El epiblasto sigue moviéndose y forma una segunda capa, el mesodermo. El epiblasto ahora se ha diferenciado en las tres capas germinales del embrión, de modo que el disco bilaminar ahora es un disco trilaminar, la gástrula .
Las tres capas germinales son el ectodermo , mesodermo y endodermo , y están formadas como tres discos planos superpuestos. Es de estas tres capas de donde se derivarán todas las estructuras y órganos del cuerpo a través de los procesos de somitogénesis , histogénesis y organogénesis . [16] El endodermo embrionario se forma por invaginación de células epiblásticas que migran al hipoblasto, mientras que el mesodermo está formado por las células que se desarrollan entre el epiblasto y el endodermo. En general, todas las capas germinales derivarán del epiblasto. [11] [15] La capa superior del ectodermo dará lugar a la capa más externa de la piel, los sistemas nerviosos central y periférico , los ojos , el oído interno y muchos tejidos conectivos . [17] La capa media del mesodermo dará lugar al corazón y al inicio del sistema circulatorio así como a los huesos , músculos y riñones . La capa interna del endodermo servirá como punto de partida para el desarrollo de los pulmones , el intestino , la tiroides , el páncreas y la vejiga .
Después de la ingresión, se desarrolla un blastoporo en el lugar donde las células han ingresado, en un lado del embrión, y se profundiza para convertirse en el arquenterón , la primera etapa formativa del intestino . Como en todos los deuteróstomos , el blastoporo se convierte en el ano , mientras que el intestino atraviesa el embrión hasta el otro lado, donde la abertura se convierte en la boca. Con un tubo digestivo en funcionamiento, ahora se completa la gastrulación y puede comenzar la siguiente etapa de neurulación .


Después de la gastrulación, el ectodermo da lugar al tejido epitelial y neural , y la gástrula ahora se conoce como neurula . La placa neural que se ha formado como una placa engrosada a partir del ectodermo, continúa ensanchándose y sus extremos comienzan a doblarse hacia arriba como pliegues neurales . La neurulación se refiere a este proceso de plegado por el cual la placa neural se transforma en el tubo neural , y esto tiene lugar durante la cuarta semana. Se pliegan, a lo largo de un surco neural poco profundo que se ha formado como una línea media divisoria en la placa neural. Esto se profundiza a medida que los pliegues continúan ganando altura, cuando se unirán y se cerrarán en la cresta neural . Las células que migran por la parte más craneal de la línea primitiva forman el mesodermo paraxial , que dará lugar a los somitómeros que en el proceso de somitogénesis se diferenciarán en somitas que formarán los esclerotomos , los sindetomos , [18] los miotomas y los dermatomas para formar cartílago y hueso , tendones , dermis (piel), y músculo . El mesodermo intermedio da lugar al tracto urogenital y está formado por células que migran desde la región media de la línea primitiva. Otras células migran por la parte caudal de la línea primitiva y forman el mesodermo lateral, y aquellas células que migran por la parte más caudal contribuyen al mesodermo extraembrionario. [11] [15]
El disco embrionario comienza plano y redondo, pero eventualmente se alarga hasta tener una parte cefálica más ancha y un extremo caudal de forma estrecha. [10] Al principio, la línea primitiva se extiende en dirección cefálica y 18 días después de la fecundación retorna caudalmente hasta desaparecer. En la porción cefálica, la capa germinativa muestra diferenciación específica al inicio de la cuarta semana, mientras que en la porción caudal ocurre al final de la cuarta semana. [11] Los neuroporos craneales y caudales se hacen progresivamente más pequeños hasta que se cierran por completo (hacia el día 26) formando el tubo neural . [19]

La organogénesis es el desarrollo de los órganos que comienza entre la tercera y la octava semana y continúa hasta el nacimiento. A veces, el desarrollo completo, como en el caso de los pulmones, continúa después del nacimiento. Diferentes órganos participan en el desarrollo de los numerosos sistemas orgánicos del cuerpo.
Las células madre hematopoyéticas que dan origen a todas las células sanguíneas se desarrollan a partir del mesodermo. El desarrollo de la formación de la sangre tiene lugar en grupos de células sanguíneas, conocidos como islotes sanguíneos , en el saco vitelino . Los islotes sanguíneos se desarrollan fuera del embrión, en la vesícula umbilical, la alantoides, el tallo conector y el corion, a partir de los hemangioblastos mesodérmicos .
En el centro de una isla sanguínea, los hemangioblastos forman las células madre hematopoyéticas, que son las precursoras de todos los tipos de células sanguíneas. En la periferia de una isla sanguínea, los hemangioblastos se diferencian en angioblastos , los precursores de los vasos sanguíneos. [20]

El corazón es el primer órgano funcional en desarrollarse y comienza a latir y bombear sangre alrededor de los 22 días. [21] Los mioblastos cardíacos y las islas de sangre en el mesénquima esplácnopleúrico a cada lado de la placa neural dan lugar a la región cardiogénica . [11] : 165 Esta es un área con forma de herradura cerca de la cabeza del embrión. Para el día 19, después de la señalización celular , dos hebras comienzan a formarse como tubos en esta región, a medida que se desarrolla un lumen dentro de ellas. Estos dos tubos endocárdicos crecen y para el día 21 han migrado uno hacia el otro y se han fusionado para formar un solo tubo cardíaco primitivo, el corazón tubular . Esto es posible gracias al plegamiento del embrión que empuja los tubos hacia la cavidad torácica . [22]
También al mismo tiempo que se forman los tubos endocárdicos, ha comenzado la vasculogénesis (el desarrollo del sistema circulatorio). Esto comienza el día 18 con las células del mesodermo esplacnopleúrico que se diferencian en angioblastos que se desarrollan en células endoteliales aplanadas. Estos se unen para formar pequeñas vesículas llamadas angioquistes que se unen para formar vasos largos llamados cordones angioblásticos. Estos cordones se desarrollan en una red generalizada de plexos en la formación de la red vascular. Esta red crece por la gemación y el brote adicional de nuevos vasos en el proceso de angiogénesis . [22] Después de la vasculogénesis y el desarrollo de una vasculatura temprana, tiene lugar una etapa de remodelación vascular .
El corazón tubular forma rápidamente cinco regiones distintas. De la cabeza a la cola, estas son el infundíbulo , el bulbo cardíaco , el ventrículo primitivo , la aurícula primitiva y el seno venoso . Inicialmente, toda la sangre venosa fluye hacia el seno venoso y es impulsada desde la cola hasta la cabeza hasta el tronco arterioso . Este se dividirá para formar la aorta y la arteria pulmonar ; el bulbo cardíaco se desarrollará en el ventrículo derecho (primitivo); el ventrículo primitivo formará el ventrículo izquierdo; la aurícula primitiva se convertirá en las partes delanteras de las aurículas izquierda y derecha y sus apéndices, y el seno venoso se desarrollará en la parte posterior de la aurícula derecha , el nódulo sinoatrial y el seno coronario . [21]
El bucle cardíaco comienza a dar forma al corazón como uno de los procesos de morfogénesis , y esto se completa al final de la cuarta semana. La muerte celular programada ( apoptosis ) en las superficies de unión permite que se produzca la fusión. [22] A mediados de la cuarta semana, el seno venoso recibe sangre de las tres venas principales: la vitelina , la umbilical y la cardinal común .
Durante los dos primeros meses de desarrollo, comienza a formarse el tabique interauricular , que divide la aurícula primitiva en una aurícula derecha y otra izquierda . En un principio, comienza como una pieza de tejido en forma de medialuna que crece hacia abajo hasta convertirse en el septum primum . La forma de medialuna impide el cierre completo de las aurículas, lo que permite que la sangre se desvíe de la aurícula derecha a la izquierda a través de la abertura conocida como ostium primum . Esta abertura se cierra con el desarrollo posterior del sistema, pero antes de que esto ocurra, comienza a formarse una segunda abertura (el ostium secundum ) en la aurícula superior, lo que permite la derivación continua de la sangre. [22]
A la derecha del septum primum comienza a formarse un segundo tabique (el septum secundum ). Esto también deja una pequeña abertura, el foramen oval, que se continúa con la abertura anterior, el ostium secundum. El septum primum se reduce a una pequeña solapa que actúa como válvula del foramen oval y que permanece hasta su cierre en el nacimiento. Entre los ventrículos también se forma el septum inferius , que se desarrolla en el tabique interventricular muscular . [22]
El sistema digestivo comienza a desarrollarse a partir de la tercera semana y hacia la duodécima semana los órganos ya están posicionados correctamente.
El sistema respiratorio se desarrolla a partir del brote pulmonar , que aparece en la pared ventral del intestino anterior aproximadamente a las cuatro semanas de desarrollo. El brote pulmonar forma la tráquea y dos crecimientos laterales conocidos como brotes bronquiales, que se agrandan al comienzo de la quinta semana para formar los bronquios principales izquierdo y derecho . Estos bronquios a su vez forman bronquios secundarios (lobulares); tres a la derecha y dos a la izquierda (que reflejan el número de lóbulos pulmonares). Los bronquios terciarios se forman a partir de los bronquios secundarios.
Mientras que el revestimiento interno de la laringe se origina en el brote pulmonar , sus cartílagos y músculos se originan en el cuarto y sexto arcos faríngeos . [23]
En el embrión en desarrollo se forman tres sistemas renales diferentes : el pronefros , el mesonefros y el metanefros . Solo el metanefros se desarrolla hasta convertirse en el riñón permanente. Los tres derivan del mesodermo intermedio .
El pronefros deriva del mesodermo intermedio en la región cervical. No es funcional y se degenera antes de finalizar la cuarta semana.
El mesonefros deriva del mesodermo intermedio en los segmentos torácico superior y lumbar superior. Se forman túbulos excretores que ingresan al conducto mesonéfrico , que termina en la cloaca . El conducto mesonéfrico se atrofia en las hembras, pero participa en el desarrollo del sistema reproductivo en los machos.
El metanefros aparece en la quinta semana de desarrollo. Una excrecencia del conducto mesonéfrico, la yema ureteral , penetra en el tejido metanéfrico para formar la pelvis renal primitiva , los cálices renales y las pirámides renales . También se forma el uréter .
Entre la cuarta y la séptima semana de desarrollo, el tabique urorrectal divide la cloaca en el seno urogenital y el canal anal . La parte superior del seno urogenital forma la vejiga , mientras que la parte inferior forma la uretra . [23]
La capa superficial de la piel , la epidermis , deriva del ectodermo . La capa más profunda, la dermis , deriva del mesénquima .
La formación de la epidermis comienza en el segundo mes de desarrollo y adquiere su disposición definitiva al final del cuarto mes. El ectodermo se divide para formar una capa plana de células en la superficie conocida como peridermo. La división posterior forma las capas individuales de la epidermis .
El mesénquima que formará la dermis proviene de tres fuentes:

A finales de la cuarta semana, la parte superior del tubo neural se dobla ventralmente como la flexura cefálica a nivel del futuro mesencéfalo : el mesencéfalo . [24] Por encima del mesencéfalo se encuentra el prosencéfalo (futuro prosencéfalo) y debajo de él se encuentra el rombencéfalo (futuro rombencéfalo).
Las células de la cresta neural craneal migran a los arcos faríngeos como células madre neurales , donde se desarrollan en el proceso de neurogénesis en neuronas .
La vesícula óptica (que finalmente se convierte en el nervio óptico , la retina y el iris ) se forma en la placa basal del prosencéfalo. La placa alar del prosencéfalo se expande para formar los hemisferios cerebrales (el telencéfalo), mientras que su placa basal se convierte en el diencéfalo. Finalmente, la vesícula óptica crece para formar una excrecencia óptica.
Desde la tercera a la octava semana se desarrollan la cara y el cuello .
El oído interno , el oído medio y el oído externo tienen orígenes embriológicos distintos.
Aproximadamente a los 22 días de desarrollo, el ectodermo de cada lado del rombencéfalo se engrosa para formar las placodas óticas . Estas placodas se invaginan para formar fosas óticas y luego vesículas óticas . Las vesículas óticas luego forman componentes ventrales y dorsales.
El componente ventral forma el sáculo y el conducto coclear . En la sexta semana de desarrollo emerge el conducto coclear y penetra en el mesénquima circundante , desplazándose en forma de espiral hasta formar 2,5 vueltas al final de la octava semana. El sáculo es la parte restante del componente ventral. Permanece conectado al conducto coclear a través del estrecho ductus reuniens .
El componente dorsal forma el utrículo y los canales semicirculares .
La primera bolsa faríngea se alarga y se expande para formar el receso tubotimpánico . Este receso se diferencia para formar la mayor parte de la cavidad timpánica del oído medio y la totalidad de la trompa de Eustaquio o auditiva . La estrecha trompa auditiva conecta la cavidad timpánica con la faringe . [25]
Los huesos del oído medio, los huesecillos , derivan de los cartílagos de los arcos faríngeos . El martillo y el yunque derivan del cartílago del primer arco faríngeo , mientras que el estribo deriva del cartílago del segundo arco faríngeo .
El conducto auditivo externo se desarrolla a partir de la porción dorsal de la primera hendidura faríngea . Seis montículos auriculares, que son proliferaciones mesenquimales en los aspectos dorsales del primer y segundo arcos faríngeos, forman la aurícula del oído. [23]
Los ojos comienzan a desarrollarse desde la tercera semana hasta la décima semana.

Al final de la cuarta semana comienza el desarrollo de las extremidades . Las yemas de las extremidades aparecen en el aspecto ventrolateral del cuerpo. Consisten en una capa externa de ectodermo y una parte interna que consiste en mesénquima que se deriva de la capa parietal del mesodermo de la placa lateral . Las células ectodérmicas en el extremo distal de las yemas forman la cresta ectodérmica apical , que crea un área de células mesenquimales de rápida proliferación conocida como zona de progreso . El cartílago (parte del cual finalmente se convierte en hueso ) y el músculo se desarrollan a partir del mesénquima. [23]
Las exposiciones tóxicas en el período embrionario pueden ser causa de malformaciones congénitas importantes , ya que los precursores de los principales sistemas de órganos ahora están en desarrollo.
Cada célula del embrión preimplantacional tiene el potencial de formar todos los diferentes tipos celulares del embrión en desarrollo. Esta potencia celular significa que algunas células pueden ser extraídas del embrión preimplantacional y las células restantes compensarán su ausencia. Esto ha permitido el desarrollo de una técnica conocida como diagnóstico genético preimplantacional , mediante la cual un pequeño número de células del embrión preimplantacional creado por FIV , pueden ser extraídas mediante biopsia y sometidas a diagnóstico genético. Esto permite seleccionar embriones que no están afectados por enfermedades genéticas definidas y luego transferirlos al útero de la madre .
Se cree que los teratomas sacrococcígeos , tumores formados a partir de diferentes tipos de tejido, están relacionados con restos de estrías primitivas, que normalmente desaparecen. [10] [11] [13]
Los síndromes del primer arco son trastornos congénitos de deformidades faciales, causados por la falla de las células de la cresta neural para migrar al primer arco faríngeo.
La espina bífida es un trastorno congénito que es el resultado del cierre incompleto del tubo neural.
Las infecciones de transmisión vertical pueden pasar de la madre al feto en cualquier etapa de su desarrollo .
La hipoxia, un estado de suministro inadecuado de oxígeno, puede ser una consecuencia grave de un parto prematuro .






