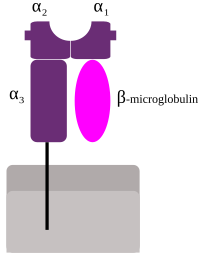
[4] El CMH es la zona del genoma humano más variable y contiene numerosos genes funcionales, caracterizados por un gran polimorfismo.Los productos de estos genes son mencionados por muchos autores también como CMH o bien HLA.Su investigación fue seguida por George Snell quién planteó que el rechazo de tejidos por el ratón era debido a incompatibilidad en algunos antígenos.De las regiones MHC secuenciadas completamente, la menos compleja es la del pollo, que contiene solo 19 genes en 92 kb.[6] La región MHC en los marsupiales Monodelphis doméstica (Didelphimorphia gris de cola corta) está flanqueada por los marcadores genéticos MOG y COL11A2, comprende 3.95 Mb y contiene 114 genes, 87 compartidos con los humanos.Son altamente polimórficos: se han descrito alrededor de 3000 alelos para cada uno.Junto a estos genes «clásicos» (o 1a) se ubican otros denominados «no clásicos» (o 1b): HLA-E, HLA-F, HLA-G, HFE y los MIC (acrónimo de cadena relacionadas con CMH de clase I, en inglés, MHC class I Chain-related): MICA y MICB.Excepto los MIC, todos estos genes codifican glucoproteínas, expresándose de manera codominante como heterodímeros en la membrana de cada célula nucleada, unido en forma no covalente con la cadena β2 microglobulina codificada por un gen del cromosoma 15.[1][2][4] Sus productos tienen estructura de inmunoglobulina: presentan una cadena pesada tipo α que se subdivide en tres regiones: α1, α2 y α3.En humanos, existen muchos isotipos (genes diferentes) de las moléculas de Clase-I, que pueden agruparse en: Los genes del CMH de clase II se encuentran en la región más cercana al centrómero, y codifican las dos cadenas del heterodímero funcional: HLA-DR, HLA-DQ, HLA-DP, HLA-DM, y HLA-DO.Inicialmente fueron denominados genes Ir por estar involucrados en la respuesta inmune (del acrónimo inglés para immune response).Entre las regiones ocupadas por los genes de clase I y de clase II, se encuentran numerosos genes que tienen importancia en la respuesta inmune pero no están relacionadas al CMH.El polimorfismo es tan grande que en una población mixta (no endogámica) no existen dos individuos que tengan exactamente el mismo juego de genes y moléculas MHC, excepto los gemelos idénticos.Esta propiedad es muy importante en el trasplante de órganos, e implica que, durante su desarrollo, los linfocitos T deben "aprender" a reconocer las moléculas MHC propias del individuo, mediante el proceso complejo de maduración y selección que tiene lugar en el timo.Las moléculas del CMH solo pueden presentar péptidos, lo que implica que los linfocitos T, dado que solo pueden reconocer un antígeno si viene asociado a una molécula del CMH, solo pueden reaccionar ante antígenos de origen proteico (procedentes de microbios) y no a otro tipo de compuestos químicos (ni lípidos, ni ácidos nucleicos, ni azúcares).Sin embargo, una molécula del CMH dada tiene una especificidad amplia, porque puede presentar muchos péptidos diferentes (aunque no todos).Por tanto, los péptidos que presentan las moléculas del CMH provienen de microbios que están en el interior celular, y ésta es la razón por la cual los linfocitos T, que solo identifican péptidos cuando están asociados a moléculas del CMH, solo detectan microbios asociados a células y desencadenan una respuesta inmune contra microbios intracelulares.Las moléculas del CMH cargadas con un péptido pueden permanecer en la membrana durante días, el tiempo suficiente para asegurar que un linfocito T adecuado reconozca el complejo e inicie la respuesta inmunitaria.Los péptidos propios, por otro lado, no pueden iniciar una respuesta inmune (excepto en los casos de las enfermedades autoinmunes), porque los linfocitos T específicos para los antígenos propios son destruidos o inactivados en el timo.Sin embargo, la presencia de péptidos propios asociados a las moléculas MHC es esencial para la función supervisora de los linfocitos T: estas células están constantemente patrullando el organismo, verificando la presencia de péptidos propios asociados a las moléculas del CMH y desencadenando una respuesta inmune en los casos raros en los que detectan un péptido extraño.Por esta razón, los linfocitos del individuo receptor interpretan el complejo presente en las células del órgano trasplantado como "péptido extraño:MHC propio" y desencadenan una respuesta inmune contra el órgano "invasor", porque lo perciben de la misma manera que un tejido propio infectado o tumoral, pero con un número mucho más elevado de complejos capaces de iniciar una respuesta.Pueden producirse dos tipos de rechazo de trasplantes mediado por las moléculas del CHM o antígenos leucocitarios humanos (HLA): En ambos casos, se produce una reacción inmune contra el órgano trasplantado, que puede generar lesiones en el mismo, lo que conlleva la pérdida de función, inmediata en el primer caso y progresiva en el segundo.La compatibilidad total solo existe entre gemelos idénticos, pero en la actualidad existen bases de datos de donantes a nivel mundial que permiten optimizar la compatibilidad HLA entre un donante potencial y un receptor.
Molécula de MHC-I
Estructura de una molécula MHC de Clase-I.
Estructura de una molécula MHC de Clase-II.
Expresión codominante de los genes HLA / MHC.
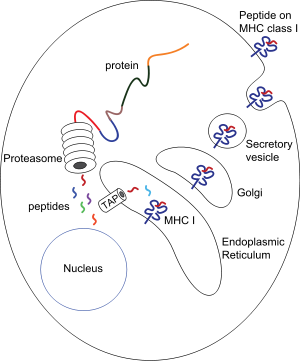
Procesamiento de péptidos asociados a moléculas del CMH de clase I: proteínas presentes en el
citosol
son degradadas por el
proteasoma
, y los péptidos resultantes son internalizados por el canal TAP en el
retículo endoplásmico
, donde se asocian con las moléculas recién sintetizadas del CMH de clase I. Los complejos
péptido-CMH de clase I
pasan al
aparato de Golgi
, donde son glucosilados, y de ahí a vesículas secretoras, que se fusionan con la
membrana celular
, de forma que los complejos quedan expuestos hacia el exterior, permitiendo el contacto con los linfocitos T circulantes.