Actina
El origen evolutivo de esta proteína se puede rastrear en las células procariotas, donde existen equivalentes.
Szent-Györgyi había descrito previamente una forma más viscosa de miosina, producida por extracciones lentas en músculo, como «miosina activada» y puesto que la proteína de Straub producía el efecto activador, la denominó actina.
La viscosidad disminuía si se añadía ATP a la mezcla de ambas proteínas, conocida como actomiosina.
[8] Straub continuó trabajando en la actina hasta 1950, publicando que podía unirse al ATP y que, durante la polimerización de la proteína para formar microfilamentos, se hidrolizaba a ADP + Pi, el cual permanecía unido al microfilamento.
Otros enfoques, como el uso de criomicroscopía electrónica o radiación sincrotrón han permitido recientemente[¿cuándo?]
[21] La actina es una de las proteínas más abundantes entre los eucariotas y se encuentra presente en todo el citoplasma.
Comienza con un aspartato acetilado en su grupo amino, mientras que su C-terminal es básico, formado por una fenilalanina precedida por una cisteína de cierta importancia funcional.
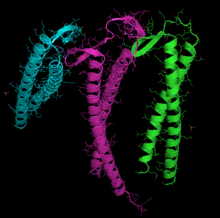
[28] Está formada por dos dominios conocidos como grande y pequeño, separados por una hendidura en cuyo centro se sitúa el lugar de unión al ATP-ADP+Pi.
Para formar parte de un filamento, según este modelo, los monómeros estarían en una configuración llamada «plana», en la que los subdominios giran entre sí, y que también parece encontrarse en el homólogo bacteriano de la actina MreB.
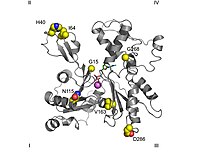
[41] Se emplean subunidades de reconocimiento diferentes para la actina y la tubulina, aunque solapadas.
[42] Tanto la actina como la tubulina se unen a la CCT en conformaciones abiertas en ausencia de ATP.
Aunque existe mucha polémica al respecto, parece claro que para la hidrólisis de ATP se precisa una conformación «cerrada», y se cree que acerca los residuos implicados a la distancia adecuada.
Según este modelo, la polimerización y la función ATPasa estarían desacopladas en un primer momento.
En cualquier caso, no se libera el Pi resultante, sino que permanece un tiempo unido no covalentemente a la actina ADP, con lo cual existirían tres especies de actina en un filamento: ATP-Actina, ADP+Pi-Actina y ADP-Actina.
[51] Por ejemplo, existen elementos que secuestran la actina G, impidiendo su incorporación a los microfilamentos.
Es decir, si un microfilamento, que posee dos extremos a los que pueden unirse o disociarse monómeros, es cortado dos veces, resultan tres nuevos microfilamentos con seis extremos; la nueva situación favorece la dinámica de ensamblaje y desensamblaje.
[68] Los microfilamentos intervienen en el movimiento de todas las células móviles, incluso las no musculares, pues se ha descrito que los fármacos que desorganizan la actina F (como las citocalasinas) afectan a la actividad de dichas células.
[72] Si bien las células vegetales poseen generalmente una pared que define su morfología e impide su movimiento, sus microfilamentos generan las fuerzas necesarias para varias actividades celulares, por ejemplo, las corrientes citoplasmáticas generadas por los microfilamentos y las miosinas.
[77] El proceso global se dispara mediante una señal externa, típicamente mediante un potencial de acción excitador del músculo que alberga las células especializadas ricas en filamentos de actina y miosina en su interior.
[94] A fecha de 2006, el ENMC (European Neuromuscular Centre) había publicado 116 mutaciones relacionadas con patologías, conocidas como actinopatías.
También se ha detectado mutaciones que producen miopatía con cores (zonas desprovistas de actividad oxidativa).
[103] El número de afecciones en las que podría estar implicado este gen está en aumento.
Se le ha relacionado con la enfermedad de Moyamoya, y parece ser que algunas mutaciones en heterocigosis podrían conferir predisposición a muchas patologías vasculares, como el aneurisma de aorta torácica y la cardiopatía isquémica.
Se secuenció por primera vez por Hamada y colaboradores en 1982, observándose que estaba interrumpido por cinco intrones.
[106] Fue el primer gen de los seis donde se encontraron alelos implicados en procesos patológicos.
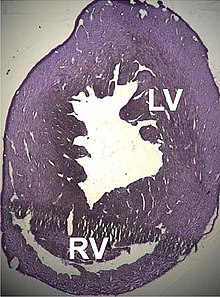
[113] Recientemente se han encontrado mutaciones de ACTC1 implicadas en otros dos procesos patológicos: la miocardiopatía restrictiva idiopática infantil,[114] y el miocardio ventricular izquierdo no compacto.
Existen multitud de pseudogenes repartidos en todo el genoma, y su secuencia contiene seis exones que pueden dar lugar hasta 21 transcritos diferentes por splicing alternativo, conocidos como actinas β.
La β actina es la proteína más abundante en los tejidos humanos, pero no así en las células ciliadas, lo que explicaría la localización de la patología.
No obstante, MreB y ParM poseen una alta similitud estructural con la actina eucariota.
ParM, un homólogo de la actina codificado en un plásmido, interviene en la gestión del ADN plasmídico.