Tyrannosauridae (o tiranosáuridos , que significa " lagartos tiranos ") es una familia de dinosaurios terópodos celurosaurios que comprende dos subfamilias que contienen hasta quince géneros , incluido el epónimo Tyrannosaurus . El número exacto de géneros es controvertido, ya que algunos expertos reconocen tan solo tres. Todos estos animales vivieron cerca del final del Período Cretácico y sus fósiles se han encontrado solo en América del Norte y Asia .
Aunque descienden de ancestros más pequeños , los tiranosáuridos casi siempre fueron los depredadores más grandes en sus respectivos ecosistemas , lo que los coloca en la cúspide de la cadena alimentaria . La especie más grande fue Tyrannosaurus rex , el depredador terrestre más masivo conocido, que medía más de 13 metros (43 pies) de longitud [2] y según la mayoría de las estimaciones modernas hasta 8,87 toneladas métricas (9,78 toneladas cortas) de peso. [3] [4] Los tiranosáuridos eran carnívoros bípedos con cráneos masivos llenos de dientes grandes. A pesar de su gran tamaño, sus piernas eran largas y proporcionadas para un movimiento rápido. En contraste, sus brazos eran muy pequeños, con solo dos dígitos funcionales .
A diferencia de la mayoría de los otros grupos de dinosaurios, se han descubierto restos muy completos de la mayoría de los tiranosáuridos conocidos. Esto ha permitido realizar diversas investigaciones sobre su biología . Los estudios científicos se han centrado en su ontogenia , biomecánica y ecología , entre otros temas.

Los primeros restos de tiranosáuridos fueron descubiertos durante expediciones lideradas por el Servicio Geológico de Canadá , que localizó numerosos dientes dispersos. Estos distintivos dientes de dinosaurio recibieron el nombre de Deinodon ("diente terrible") por Joseph Leidy en 1856. Los primeros buenos ejemplares de un tiranosáurido fueron encontrados en la Formación Horseshoe Canyon de Alberta , y consistían en cráneos casi completos con esqueletos parciales. Estos restos fueron estudiados por primera vez por Edward Drinker Cope en 1876, quien los consideró una especie del tiranosáurido oriental Dryptosaurus . En 1905, Henry Fairfield Osborn reconoció que los restos de Alberta diferían considerablemente de Dryptosaurus , y acuñó un nuevo nombre para ellos: Albertosaurus sarcophagus ("lagarto carnívoro de Alberta"). [5] Cope describió más material de tiranosáurido en 1892, en forma de vértebras aisladas, y le dio a este animal el nombre de Manospondylus gigas . Este descubrimiento fue pasado por alto durante más de un siglo y causó controversia a principios de la década de 2000 cuando se descubrió que este material en realidad pertenecía a Tyrannosaurus rex y tenía prioridad de nombre sobre él . [6]

En su artículo de 1905 en el que nombró a Albertosaurus , Osborn describió dos especímenes de tiranosaurio adicionales que habían sido recolectados en Montana y Wyoming durante una expedición de 1902 del Museo Americano de Historia Natural , dirigida por Barnum Brown . Inicialmente, Osborn consideró que se trataba de especies distintas. Al primero, lo llamó Dynamosaurus imperiosus ("lagarto del poder del emperador"), y al segundo, Tyrannosaurus rex ("lagarto tirano rey"). Un año después, Osborn reconoció que estos dos especímenes en realidad provenían de la misma especie. A pesar del hecho de que Dynamosaurus había sido encontrado primero, el nombre Tyrannosaurus había aparecido una página antes en su artículo original que describía a ambos especímenes. Por lo tanto, de acuerdo con el Código Internacional de Nomenclatura Zoológica (ICZN), se utilizó el nombre Tyrannosaurus . [7]
Barnum Brown recolectó varios especímenes más de tiranosáuridos de Alberta, incluido el primero en preservar las extremidades anteriores acortadas y con dos dedos características del grupo (al que Lawrence Lambe llamó Gorgosaurus libratus , "lagarto feroz equilibrado", en 1914). Un segundo hallazgo significativo atribuido a Gorgosaurus se realizó en 1942, en forma de un cráneo completo bien conservado, aunque inusualmente pequeño. El espécimen esperó hasta después del final de la Segunda Guerra Mundial para ser estudiado por Charles W. Gilmore , quien lo nombró Gorgosaurus lancesnis . [5] Este cráneo fue reestudiado por Robert T. Bakker , Phil Currie y Michael Williams en 1988, y asignado al nuevo género Nanotyrannus . [8] También fue en 1946 que los paleontólogos de la Unión Soviética comenzaron expediciones a Mongolia y descubrieron los primeros restos de tiranosáurido de Asia. En 1955, Evgeny Maleev describió nuevas especies mongolas de Tyrannosaurus y Gorgosaurus , y un nuevo género: Tarbosaurus ("lagarto aterrador"). Sin embargo, estudios posteriores demostraron que todas las especies de tiranosaurios de Maleev eran en realidad una especie de Tarbosaurus en diferentes etapas de crecimiento. Una segunda especie de tiranosáurido mongol fue descubierta más tarde, descrita por Sergei Kurzanov en 1976, y recibió el nombre de Alioramus remotus ("rama diferente y remota"), aunque su estatus como un verdadero tiranosáurido y no un tiranosáurido más primitivo aún es controvertido. [9] [5]

Los tiranosáuridos eran todos animales grandes, y todas las especies podían pesar al menos una tonelada métrica. [10] Se ha descubierto un único ejemplar de Alioramus de un individuo estimado entre 5 y 6 metros (16 y 20 pies) de largo, [9] aunque algunos expertos lo consideran un juvenil. [10] [11] Albertosaurus , Gorgosaurus y Daspletosaurus medían entre 8 y 9 metros (26 y 30 pies) de largo, [12] mientras que Tarbosaurus alcanzaba longitudes de 11 metros (36 pies) desde el hocico hasta la cola. [13] El enorme Tyrannosaurus alcanzó los 13 metros (43 pies) en el espécimen más grande, RSM P2523.8 . [14]
La anatomía del cráneo de los tiranosáuridos es bien conocida, ya que se conocen cráneos completos de todos los géneros excepto Alioramus , que se conoce solo a partir de restos de cráneos parciales. [15] Tyrannosaurus , Tarbosaurus y Daspletosaurus tenían cráneos que superaban 1 m (3,3 pies) de longitud. [12] Los tiranosáuridos adultos tenían cráneos altos y masivos, con muchos huesos fusionados y reforzados para mayor resistencia. Al mismo tiempo, las cámaras huecas dentro de muchos huesos del cráneo y las grandes aberturas ( fenestras ) entre esos huesos ayudaron a reducir el peso del cráneo. Muchas características de los cráneos de los tiranosáuridos también se encontraron en sus antepasados inmediatos, incluidos premaxilares altos y huesos nasales fusionados . [10]
Los cráneos de los tiranosáuridos tenían muchas características únicas, incluyendo huesos parietales fusionados con una cresta sagital prominente , que corría longitudinalmente a lo largo de la sutura sagital y separaba las dos fenestras supratemporales en el techo del cráneo. Detrás de estas fenestras, los tiranosáuridos tenían una cresta nucal característicamente alta, que también surgía de los parietales pero corría a lo largo de un plano transversal en lugar de longitudinalmente. La cresta nucal estaba especialmente bien desarrollada en Tyrannosaurus , Tarbosaurus y Alioramus . Albertosaurus , Daspletosaurus y Gorgosaurus tenían crestas altas frente a los ojos en los huesos lagrimales , mientras que Tarbosaurus y Tyrannosaurus tenían huesos postorbitales extremadamente engrosados que formaban crestas en forma de medialuna detrás de los ojos. Alioramus tenía una fila de seis crestas óseas en la parte superior de su hocico, que surgían de los huesos nasales; Se han reportado crestas inferiores en algunos especímenes de Daspletosaurus y Tarbosaurus , así como en el tiranosáurido más basal Appalachiosaurus . [11] [16] El hocico y otras partes del cráneo también lucían numerosos agujeros . Según el estudio de 2017 que describió a D. horneri , el tegumento escamoso, así como la sensibilidad táctil, se correlacionaron con las múltiples filas de agujeros neurovasculares observados en cocodrilos y tiranosáuridos. [17]
Los tiranosáuridos, al igual que sus ancestros tiranosáuridos, eran heterodontes , con dientes premaxilares en forma de D en sección transversal y más pequeños que el resto. A diferencia de los tiranosáuridos anteriores y la mayoría de los otros terópodos, los dientes maxilares y mandibulares de los tiranosáuridos maduros no tienen forma de cuchilla, sino que son extremadamente engrosados y a menudo circulares en sección transversal, y algunas especies tienen dentados reducidos. [10] Los recuentos de dientes tienden a ser consistentes dentro de las especies, y las especies más grandes tienden a tener recuentos de dientes más bajos que las más pequeñas. Por ejemplo, Alioramus tenía entre 76 y 78 dientes en sus mandíbulas, mientras que Tyrannosaurus tenía entre 54 y 60. [18]
William Abler observó en 2001 que las dentaduras de los dientes de Albertosaurus se asemejan a una grieta en el diente que termina en un hueco redondo llamado ampolla. [19] Los dientes de los tiranosáuridos se usaban como puntos de sujeción para arrancar la carne de un cuerpo, por lo que cuando un tiranosáurido hubiera tirado de un trozo de carne, la tensión podría causar que una dentada puramente parecida a una grieta se extendiera a través del diente. [19] Sin embargo, la presencia de la ampolla habría distribuido estas fuerzas sobre una superficie más grande y disminuido el riesgo de daño al diente bajo tensión. [19] La presencia de incisiones que terminan en huecos tiene paralelos en la ingeniería humana. Los fabricantes de guitarras usan incisiones que terminan en huecos para, como describe Abler, "impartir regiones alternas de flexibilidad y rigidez" a la madera con la que trabajan. [19] El uso de un taladro para crear una especie de "ampolla" y evitar la propagación de grietas a través del material también se usa para proteger las superficies de los aviones . [19] Abler demostró que una barra de plexiglás con incisiones llamadas "ranuras" y agujeros perforados era más de un 25% más fuerte que una con solo incisiones colocadas de manera regular. [19] A diferencia de los tiranosaurios y otros terópodos, los depredadores antiguos como los fitosaurios y el Dimetrodon no tenían adaptaciones para evitar que las dentadas de sus dientes, similares a grietas, se extendieran cuando se los sometía a las fuerzas de la alimentación. [19]

El cráneo estaba posado al final de un cuello grueso en forma de S , y una cola larga y pesada actuaba como contrapeso para equilibrar la cabeza y el torso, con el centro de masa sobre las caderas. Los tiranosáuridos son conocidos por sus extremidades anteriores proporcionalmente muy pequeñas de dos dedos, aunque a veces se encuentran restos de un tercer dedo vestigial . [10] [20] Tarbosaurus tenía las extremidades anteriores más cortas en comparación con el tamaño de su cuerpo, mientras que Daspletosaurus tenía las más largas.

Los tiranosáuridos caminaban exclusivamente sobre sus extremidades traseras, por lo que los huesos de sus piernas eran enormes. A diferencia de las extremidades delanteras, las extremidades traseras eran más largas en comparación con el tamaño corporal que casi cualquier otro terópodo. Los juveniles e incluso algunos adultos más pequeños, como los tiranosáuridos más basales, tenían tibias más largas que fémures , una característica de los dinosaurios cursoriales (de carrera rápida) como los ornitomímidos . Los adultos más grandes tenían proporciones de piernas características de animales de movimiento más lento, pero no en la medida observada en otros terópodos grandes como los abelisáuridos o los carnosáuridos . Los terceros metatarsianos de los tiranosáuridos estaban pinzados entre el segundo y el cuarto metatarsianos, formando una estructura conocida como arctometatarso . [10] Los tiranosáuridos también tenían grandes inserciones musculares de las piernas preservadas y baja inercia rotacional en relación con su masa corporal, lo que indica que podían girar más rápidamente que otros terópodos grandes. [21] [22]
No está claro cuándo evolucionó por primera vez el arctometatarso; no estaba presente en los primeros tiranosáuridos como Dilong , [23] pero se encontró en el posterior Appalachiosaurus . [16] Esta estructura también caracterizó a los troodóntidos , ornitomímidos y caenagnátidos , [24] pero su ausencia en los primeros tiranosáuridos indica que fue adquirida por evolución convergente . [23]
Un análisis comparativo de los dientes sugiere que los tiranosáuridos, así como los otros grandes terópodos, tenían labios que protegían sus dientes de daños externos. Esta característica anatómica les daba a estos animales un mayor parecido visual con los lepidosáuridos que con sus parientes cercanos, los cocodrilos. [25]
El nombre Deinodontidae fue acuñado por Edward Drinker Cope en 1866 para esta familia, [26] y continuó siendo usado en lugar del nombre más nuevo Tyrannosauridae hasta la década de 1960. [27] El género tipo de Deinodontidae es Deinodon , que recibió su nombre a partir de dientes aislados de Montana . [28] Sin embargo, en una revisión de 1970 de los tiranosaurios norteamericanos, Dale Russell concluyó que Deinodon no era un taxón válido y usó el nombre Tyrannosauridae en lugar de Deinodontidae, afirmando que esto estaba de acuerdo con las reglas de ICZN . [12] Por lo tanto, los expertos modernos prefieren Tyrannosauridae. [5]
El nombre Tyrannosaurus fue dado a conocer por Henry Fairfield Osborn en 1905, junto con la familia Tyrannosauridae. [29] El nombre se deriva de las palabras griegas antiguas τυραννος ( tyrannos ) ('tirano') y σαυρος ( sauros ) ('lagarto'). El sufijo muy común -idae normalmente se añade a los nombres de las familias zoológicas y se deriva del sufijo griego -ιδαι - idai , que indica un sustantivo plural. [30]

Tyrannosauridae es una familia en la taxonomía linneana basada en rangos , dentro de la superfamilia Tyrannosauroidea y el suborden Theropoda .
Tyrannosauridae se divide indiscutiblemente en dos subfamilias. Albertosaurinae comprende los géneros norteamericanos Albertosaurus y Gorgosaurus , mientras que Tyrannosaurinae incluye a Daspletosaurus , Teratophoneus , Bistahieversor , Tarbosaurus , Nanuqsaurus , Zhuchengtyrannus y al propio Tyrannosaurus . [31] Algunos autores incluyen las especies Gorgosaurus libratus en el género Albertosaurus y Tarbosaurus bataar en el género Tyrannosaurus , [16] [5] [32] mientras que otros prefieren mantener a Gorgosaurus y Tarbosaurus como géneros separados. [10] [11] Los albertosaurinos se caracterizan por estructuras más delgadas, cráneos más bajos y tibias proporcionalmente más largas que los tiranosaurios. [10] En los tiranosaurios, la cresta sagital en los parietales continúa hacia adelante sobre los frontales. [11] En 2014, Lü Junchang et al. describieron a los Alioramini como una tribu dentro de los Tyrannosauridae que contiene los géneros Alioramus y Qianzhousaurus . Su análisis filogenético indicó que la tribu estaba ubicada en la base de los Tyrannosaurinae. [33] [34] Algunos autores, como George Olshevsky y Tracy Ford, han creado otras subdivisiones o tribus para varias combinaciones de tiranosáuridos dentro de las subfamilias. [35] [36] Sin embargo, estas no han sido definidas filogenéticamente y generalmente consistían en géneros que ahora se consideran sinónimos de otros géneros o especies. [18]
Se han nombrado subfamilias adicionales para géneros más fragmentarios, incluidos Aublysodontinae y Deinodontinae . Sin embargo, los géneros Aublysodon y Deinodon generalmente se consideran nomina dubia , por lo que ellos y sus subfamilias epónimas generalmente se excluyen de las taxonomías de tiranosáuridos. Un tiranosáurido adicional, Raptorex , fue descrito inicialmente como un tiranosáuroide más primitivo, pero probablemente representa un tiranosáurido juvenil similar a Tarbosaurus . Sin embargo, como solo se conoce de un espécimen juvenil, actualmente también se considera un nomen dubium . [37] Sin embargo, Thomas Carr mantiene su validez y lo considera similar a los tiranosáuridos. [38]
Con el advenimiento de la taxonomía filogenética en la paleontología de vertebrados, Tyrannosauridae ha recibido varias definiciones explícitas. La original fue producida por Paul Sereno en 1998, e incluía todos los tiranosáuridos más cercanos a Tyrannosaurus que a Alectrosaurus , Aublysodon o Nanotyrannus . [39] Sin embargo, Nanotyrannus suele considerarse un Tyrannosaurus rex juvenil , mientras que Aublysodon suele considerarse un nomen dubium inadecuado para su uso en la definición de un clado . [10] Las definiciones desde entonces se han basado en géneros mejor establecidos.
En 2001, Thomas R. Holtz Jr. publicó un análisis cladístico de los Tyrannosauridae. [40] Concluyó que había dos subfamilias : los Aublysodontinae más primitivos, caracterizados por dientes premaxilares no aserrados ; y los Tyrannosaurinae . [40] Los Aublysodontinae incluían a Aublysodon , el " Aublysodon de Kirtland " y Alectrosaurus . [40] Holtz también encontró que Siamotyrannus exhibía algunas de las sinapomorfías de los Tyrannosauridae, pero se encontraba "fuera de la [ familia ] propiamente dicha". [40]
Más adelante en el mismo artículo, propuso que Tyrannosauridae se definiera como "todos los descendientes del ancestro común más reciente de Tyrannosaurus y Aublysodon ". [40] También criticó las definiciones propuestas previamente por otros investigadores, como una propuesta por Paul Sereno , de que Tyrannosauridae era "todos los taxones más cercanos a "Tyrannosaurus" que a Alectrosaurus , Aublysodon y Nanotyrannus ". [40] Holtz observó que dado que Nanotyrannus probablemente era un juvenil de T. rex mal identificado , la definición propuesta por Sereno tendría a la familia Tyrannosauridae como un subtaxón del género Tyrannosaurus . [40] Además, su definición propuesta de la subfamilia Tyrannosaurinae también se limitaría a Tyrannosaurus . [40]
Un intento de 2003 por Christopher Brochu incluyó a Albertosaurus , Alectrosaurus , Alioramus , Daspletosaurus , Gorgosaurus , Tarbosaurus y Tyrannosaurus en la definición. [41] Holtz redefinió el clado en 2004 para usar todos los anteriores como especificadores excepto Alioramus y Alectrosaurus , que su análisis no pudo ubicar con certeza. Sin embargo, en el mismo artículo, Holtz también proporcionó una definición completamente diferente, incluyendo a todos los terópodos más estrechamente relacionados con Tyrannosaurus que con Eotyrannus . [10] La definición más reciente es la de Sereno en 2005, que definió a Tyrannosauridae como el clado menos inclusivo que contiene a Albertosaurus , Gorgosaurus y Tyrannosaurus . [42]
Los análisis cladísticos de la filogenia de los tiranosáuridos a menudo encuentran que Tarbosaurus y Tyrannosaurus son taxones hermanos , con Daspletosaurus más basal que cualquiera de ellos. Una relación cercana entre Tarbosaurus y Tyrannosaurus está respaldada por numerosas características del cráneo, incluido el patrón de suturas entre ciertos huesos, la presencia de una cresta en forma de medialuna en el hueso postorbital detrás de cada ojo y un maxilar muy profundo con una notable curva hacia abajo en el borde inferior, entre otros. [10] [16] Una hipótesis alternativa fue presentada en un estudio de 2003 por Phil Currie y colegas, que encontró un apoyo débil para Daspletosaurus como miembro basal de un clado que también incluye a Tarbosaurus y Alioramus , ambos de Asia, basado en la ausencia de una punta ósea que conecta los huesos nasal y lagrimal. [18] Alioramus resultó ser el pariente más cercano de Tarbosaurus en este estudio, basado en un patrón similar de distribución de estrés en el cráneo.
Un estudio relacionado también señaló un mecanismo de bloqueo en la mandíbula inferior compartido entre los dos géneros. [43] En un artículo separado, Currie señaló la posibilidad de que Alioramus pudiera representar un Tarbosaurus juvenil , pero afirmó que el recuento de dientes mucho mayor y las crestas nasales más prominentes en Alioramus sugieren que es un género distinto. De manera similar, Currie usa el alto recuento de dientes de Nanotyrannus para sugerir que puede ser un género distinto, [11] en lugar de un Tyrannosaurus juvenil como creen la mayoría de los otros expertos. [10] [44] Sin embargo, el descubrimiento y la descripción de Qianzhousaurus revela que Alioramus no es un pariente cercano de Tarbosaurus , sino que pertenece a una tribu de tiranosáuridos recientemente descrita; los Alioramini. Qianzhousaurus revela además que tiranosáuridos de hocico largo similares estaban ampliamente distribuidos por toda Asia y habrían compartido el mismo entorno mientras evitaban la competencia con tiranosáuridos más grandes y robustos al cazar presas diferentes. [45]
El paleontólogo Gregory Erickson y sus colegas han estudiado el crecimiento y la historia de vida de los tiranosáuridos. El análisis de la histología ósea puede determinar la edad de un espécimen cuando murió. Las tasas de crecimiento se pueden examinar cuando se traza la edad de varios individuos contra su tamaño en un gráfico. Erickson ha demostrado que después de un largo tiempo como juveniles, los tiranosáuridos experimentaron tremendos brotes de crecimiento durante unos cuatro años a mitad de sus vidas. Después de que la fase de crecimiento rápido terminó con la madurez sexual , el crecimiento se desaceleró considerablemente en los animales adultos. La curva de crecimiento de un tiranosáurido tiene forma de S, y la tasa de crecimiento máxima de los individuos alrededor de los 14 años de edad. [49]

Se estima que el individuo más pequeño conocido de Tyrannosaurus rex ( LACM 28471, el "terópodo de Jordan") pesaba solo 29,9 kilogramos (66 libras) con solo 2 años de edad, mientras que el más grande, como FMNH PR2081 (" Sue "), probablemente pesaba alrededor de 5654 kilogramos (12 465 libras), y se estima que tenía 28 años, una edad que puede haber estado cerca del máximo para la especie. [49] Los juveniles de T. rex se mantuvieron por debajo de los 1800 kilogramos (4000 libras) hasta aproximadamente los 14 años de edad, cuando el tamaño corporal comenzó a aumentar drásticamente. Durante esta fase de rápido crecimiento, un T. rex joven ganaría un promedio de 600 kilogramos (1300 libras) al año durante los siguientes cuatro años. Esto se desaceleró después de los 16 años y, a los 18 años de edad, la curva se estabiliza nuevamente, lo que indica que el crecimiento se desaceleró drásticamente. [50] Por ejemplo, sólo 600 kg (1.300 lb) separaban a "Sue", de 28 años, de un ejemplar canadiense de 22 años ( RTMP 81.12.1). [49] Este cambio repentino en la tasa de crecimiento puede indicar madurez física, una hipótesis que está respaldada por el descubrimiento de tejido medular en el fémur de un T. rex de 18 años de Montana ( MOR 1125, también conocido como "B-rex"). [51] El tejido medular se encuentra sólo en aves hembras durante la ovulación, lo que indica que el "B-rex" estaba en edad reproductiva. [52]
Otros tiranosáuridos exhiben curvas de crecimiento extremadamente similares, aunque con tasas de crecimiento más bajas correspondientes a sus tamaños adultos más bajos. [53] Comparado con los albertosaurinos, Daspletosaurus mostró una tasa de crecimiento más rápida durante el período de crecimiento rápido debido a su mayor peso adulto. La tasa máxima de crecimiento en Daspletosaurus fue de 180 kilogramos (400 lb) por año, basada en una estimación de masa de 1.800 kg (4.000 lb) en adultos. Otros autores han sugerido pesos adultos más altos para Daspletosaurus ; esto cambiaría la magnitud de la tasa de crecimiento, pero no el patrón general. [49] El Albertosaurus más joven conocido es un Albertosaurus de dos años descubierto en el lecho de huesos de Dry Island, que habría pesado alrededor de 50 kg (110 lb) y medido un poco más de 2 metros (6,6 pies) de longitud. El espécimen de 10 metros (33 pies) de la misma cantera es el más antiguo y más grande conocido, con 28 años de edad. Se estima que la tasa de crecimiento más rápida ocurre alrededor de los 12 a 16 años de edad, alcanzando 122 kg (269 lb) por año, basado en un adulto de 1300 kg (2900 lb), que es aproximadamente una quinta parte de la tasa del T.-rex . Para Gorgosaurus , la tasa de crecimiento máxima calculada es de aproximadamente 110 kilogramos (240 lb) durante la fase de crecimiento rápido, que es comparable a la de Albertosaurus . [49]
El descubrimiento de un tiranosáurido embrionario de un género aún desconocido sugiere que los tiranosáuridos desarrollaron sus características esqueléticas distintivas mientras se desarrollaban en el huevo. Además, el tamaño del espécimen, un dentario de 1,1 pulgadas (2,8 cm) de la mandíbula inferior encontrado en la Formación Two Medicine de Montana en 1983 y una garra de pata encontrada en la Formación Horseshoe Canyon en 2018 y descrita en 2020, sugiere que los tiranosáuridos neonatos nacieron con cráneos del tamaño de un ratón o roedores de tamaño similar y pueden haber sido aproximadamente del tamaño de un perro pequeño al nacer. Se cree que el espécimen de mandíbula proviene de un animal de aproximadamente 2,5 pies (0,76 m), mientras que se cree que la garra pertenece a un espécimen que mide alrededor de 3 pies (0,91 m). Si bien no se han encontrado cáscaras de huevo asociadas con ninguno de los especímenes, la ubicación donde se descubrieron estos tiranosáuridos neonatos sugiere que estos animales usaban los mismos sitios de anidación que otras especies con las que vivían y de las que se alimentaban. [54] La falta de cáscaras de huevo asociadas con estos especímenes también ha abierto la especulación sobre la posibilidad de que los tiranosáuridos pusieran huevos de cáscara blanda como se cree que lo hicieron los géneros Mussaurus y Protoceratops . [55]
Las huellas fósiles de la Formación Wapiti sugieren que, a medida que los tiranosáuridos crecían, sus pies se ensanchaban y sus dedos se hacían más gruesos para soportar su peso. Los pies más anchos sugieren que los tiranosáuridos adultos se movían más lentamente que sus crías. [56] [57]

El final de la fase de crecimiento rápido sugiere el inicio de la madurez sexual en Albertosaurus , aunque el crecimiento continuó a un ritmo más lento durante toda la vida de los animales. [49] [53] La maduración sexual mientras todavía está creciendo activamente parece ser un rasgo compartido entre los dinosaurios pequeños [58] y grandes [59] , así como en los mamíferos grandes, como los humanos y los elefantes . [59] Este patrón de maduración sexual relativamente temprana difiere notablemente del patrón en las aves, que retrasan su madurez sexual hasta después de que han terminado de crecer. [59] [60]
Al tabular el número de especímenes de cada grupo de edad, Erickson y sus colegas pudieron sacar conclusiones sobre la historia de vida en las poblaciones de tiranosáuridos. Su análisis mostró que, si bien los juveniles eran raros en el registro fósil, los subadultos en la fase de crecimiento rápido y los adultos eran mucho más comunes. Más de la mitad de los especímenes conocidos de T. rex parecen haber muerto dentro de los seis años de alcanzar la madurez sexual, un patrón que también se observa en otros tiranosaurios y en algunas aves y mamíferos grandes y longevos de la actualidad. Estas especies se caracterizan por altas tasas de mortalidad infantil, seguidas de una mortalidad relativamente baja entre los juveniles. La mortalidad aumenta nuevamente después de la madurez sexual, en parte debido al estrés de la reproducción. Si bien esto podría deberse a sesgos de conservación o recolección , Erickson planteó la hipótesis de que la diferencia se debía a la baja mortalidad entre los juveniles de un cierto tamaño, que también se observa en algunos grandes mamíferos modernos, como los elefantes . Esta baja mortalidad puede haber sido resultado de una falta de depredación, ya que los tiranosaurios superaron en tamaño a todos los depredadores contemporáneos a la edad de dos años. Los paleontólogos no han encontrado suficientes restos de Daspletosaurus para un análisis similar, pero Erickson señala que parece aplicarse la misma tendencia general. [53]
Los tiranosáuridos pasaron hasta la mitad de su vida en la fase juvenil antes de crecer hasta alcanzar un tamaño casi máximo en solo unos pocos años. [49] Esto, junto con la completa falta de depredadores de tamaño intermedio entre los enormes tiranosáuridos adultos y otros terópodos pequeños, sugiere que estos nichos pueden haber sido ocupados por tiranosáuridos juveniles. Esto se ve en los dragones de Komodo modernos , donde las crías comienzan como insectívoros que viven en los árboles y maduran lentamente hasta convertirse en depredadores de ápice masivos capaces de derribar grandes vertebrados. [10] Por ejemplo, se han encontrado Albertosaurus en agregaciones que algunos han sugerido que representan manadas de edades mixtas . [61] [62]
Las habilidades de locomoción se han estudiado mejor en el caso del Tyrannosaurus , y hay dos cuestiones principales relacionadas con esto: qué tan bien podía girar y cuál era probable que fuera su velocidad máxima en línea recta. El Tyrannosaurus puede haber sido lento para girar, posiblemente tardando uno o dos segundos en girar solo 45°, una cantidad que los humanos, al estar orientados verticalmente y no tener cola, pueden girar en una fracción de segundo. [63] La causa de la dificultad es la inercia rotacional , ya que gran parte de la masa del Tyrannosaurus estaba a cierta distancia de su centro de gravedad, como un humano que lleva una madera pesada. [64]
Los científicos han elaborado una amplia gama de estimaciones de velocidad máxima, en su mayoría de alrededor de 11 metros por segundo (25 mph), pero unas pocas de tan solo 5-11 metros por segundo (11-25 mph), y unas pocas de hasta 20 metros por segundo (45 mph). Los investigadores tienen que confiar en varias técnicas de estimación porque, si bien hay muchas huellas de terópodos muy grandes caminando, hasta ahora no se ha encontrado ninguna de terópodos muy grandes corriendo, y esta ausencia puede indicar que no corrían. [65]
Jack Horner y Don Lessem argumentaron en 1993 que el Tyrannosaurus era lento y probablemente no podía correr (no hay fase aérea a mitad de la zancada). [66] Sin embargo, Holtz (1998) concluyó que los tiranosáuridos y sus parientes cercanos eran los terópodos grandes más rápidos. [67] Christiansen (1998) estimó que los huesos de las patas del Tyrannosaurus no eran significativamente más fuertes que los de los elefantes, que están relativamente limitados en su velocidad máxima y nunca corren en realidad (no hay fase aérea), y por lo tanto propuso que la velocidad máxima del dinosaurio habría sido de unos 11 metros por segundo (25 mph), que es aproximadamente la velocidad de un velocista humano. [68] Farlow y sus colegas (1995) han argumentado que un Tyrannosaurus de 6 a 8 toneladas habría resultado gravemente herido o incluso fatalmente herido si se hubiera caído mientras se movía rápidamente, ya que su torso se habría estrellado contra el suelo con una desaceleración de 6 g (seis veces la aceleración debida a la gravedad, o unos 60 metros/s 2 ) y sus diminutos brazos no podrían haber reducido el impacto. [69] [70] Sin embargo, se sabe que las jirafas galopan a 50 km/h (31 mph), a pesar del riesgo de que se rompan una pata o algo peor, lo que puede ser fatal incluso en un entorno "seguro" como un zoológico. [71] [72] Por lo tanto, es muy posible que el Tyrannosaurus también se moviera rápido cuando fuera necesario y tuviera que aceptar tales riesgos; este escenario también se ha estudiado para el Allosaurus . [73] [74] La investigación más reciente sobre la locomoción del Tyrannosaurus no limita las velocidades más allá de un rango de 17 a 40 km/h (11 a 25 mph), es decir, desde caminar o correr lentamente hasta correr a velocidad moderada. [65] [75] [76] Un estudio de modelos de computadora en 2007 estimó las velocidades de carrera, basándose en datos tomados directamente de fósiles, y afirmó que el T. rex tenía una velocidad máxima de carrera de 8 metros por segundo (18 mph). [77] [78] (Probablemente un individuo juvenil. [79] )
Estudios de Eric Snively et al., publicados en 2019, indican que los tiranosáuridos como Tarbosaurus y el propio Tyrannosaurus eran más maniobrables que los alosauroides de tamaño comparable debido a la baja inercia rotacional en comparación con su masa corporal combinada con grandes músculos en las patas. Como resultado, se plantea la hipótesis de que los tiranosáuridos eran capaces de hacer giros relativamente rápidos y probablemente podían hacer pivotar sus cuerpos más rápidamente cuando estaban cerca de su presa, o que mientras giraban, podían "hacer piruetas" sobre una sola pata plantada mientras la pata alternada se mantenía extendida en un balanceo suspendido durante la persecución. Los resultados de este estudio potencialmente podrían arrojar luz sobre cómo la agilidad podría haber contribuido al éxito de la evolución de los tiranosáuridos. [80]
Además, un estudio de 2020 indica que los tiranosáuridos eran caminantes excepcionalmente eficientes. Los estudios de Dececchi et al. compararon las proporciones de las patas, la masa corporal y la forma de andar de más de 70 especies de dinosaurios terópodos, incluidos los tiranosáuridos. Luego, el equipo de investigación aplicó una variedad de métodos para estimar la velocidad máxima de cada dinosaurio al correr, así como la cantidad de energía que gastaba cada dinosaurio mientras se movía a velocidades más relajadas, como al caminar. Entre las especies de tamaño pequeño a mediano, como los dromeosáuridos, las patas más largas parecen ser una adaptación para correr más rápido, en línea con los resultados anteriores de otros investigadores. Pero para los terópodos que pesan más de 1000 kg (2200 lb), la velocidad máxima al correr está limitada por el tamaño del cuerpo, por lo que se descubrió que las patas más largas se correlacionaban con caminar con baja energía. Los resultados del estudio indicaron además que los terópodos más pequeños desarrollaron patas largas para aumentar la velocidad como medio para ayudar en la caza y escapar de depredadores más grandes, mientras que los terópodos depredadores más grandes que desarrollaron patas largas lo hicieron para reducir los costos de energía y aumentar la eficiencia de búsqueda de alimento, ya que se liberaron de las demandas de la presión de la depredación debido a su papel como depredadores de ápice. En comparación con los grupos más basales de terópodos en el estudio, los tiranosáuridos mostraron un marcado aumento en la eficiencia de búsqueda de alimento debido a la reducción del gasto de energía durante la caza y el carroñeo. Esto probablemente resultó en que los tiranosaurios tuvieran una menor necesidad de incursiones de caza y, como resultado, requirieran menos alimento para mantenerse. Además, la investigación, junto con estudios que muestran que los tiranosaurios eran más ágiles que otros terópodos de gran tamaño, indica que estaban bastante bien adaptados a una estrategia de acecho de larga distancia seguida de una rápida ráfaga de velocidad para ir a matar. Como resultado, se pueden observar analogías entre los tiranosáuridos y los lobos modernos, respaldadas por evidencia de que al menos algunos tiranosáuridos como Albertosaurus cazaban en entornos grupales. [81] [82]

Un debate en curso en la comunidad paleontológica rodea la extensión y la naturaleza de la cubierta tegumentaria de los tiranosáuridos. Se han preservado largas estructuras filamentosas junto con restos esqueléticos de numerosos celurosaurios de la Formación Yixian del Cretácico Inferior y otras formaciones geológicas cercanas de Liaoning , China. [83] Estos filamentos generalmente se han interpretado como "protoplumas", homólogas a las plumas ramificadas que se encuentran en las aves y algunos terópodos no aviares , [84] [85] aunque se han propuesto otras hipótesis. [86] En 2004 se describió un esqueleto de Dilong que incluía el primer ejemplo de "protoplumas" en un tiranosáurido. De manera similar a las plumas de plumón de las aves modernas, las "protoplumas" encontradas en Dilong eran ramificadas pero no pennáceas , y pueden haber sido utilizadas como aislamiento . [23] El descubrimiento y descripción del tiranosáurido emplumado de 9 metros (30 pies) Yutyrannus en 2012 indica la posibilidad de que los grandes tiranosáuridos también tuvieran plumas cuando eran adultos. [87]

Basándose en el principio de horquillado filogenético , se predijo que los tiranosáuridos también podrían poseer dicho plumaje. Sin embargo, un estudio en 2017 publicado por un equipo de investigadores en Biology Letters describió impresiones de piel de tiranosáuridos recolectadas en Alberta, Montana y Mongolia, que provenían de cinco géneros ( Tyrannosaurus , Albertosaurus , Gorgosaurus , Daspletosaurus y Tarbosaurus ). [88] Aunque las impresiones de piel son pequeñas, están ampliamente dispersas en el postcráneo, y se ubican colectivamente en el abdomen, la región torácica, el íleon, la pelvis, la cola y el cuello. Muestran un patrón apretado de escamas finas y no superpuestas (que el coautor Scott Persons comparó con las observadas en los flancos de un cocodrilo [89] ) y no conservan indicios de plumas. La textura básica está compuesta por diminutas "escamas basales" de aproximadamente 1 a 2 mm de diámetro, con algunas impresiones que muestran "escamas características" de 7 mm intercaladas entre ellas. Se pueden ver escamas adicionales en las huellas de tiranosáuridos. [90] Los estudios han descubierto que el tegumento facial de los tiranosáuridos tenía escamas en el dentario y el maxilar, epidermis cornificada y piel similar a una armadura en las regiones subordinadas. [91] [92]
Bell et al. realizaron una reconstrucción de caracteres ancestrales basándose en lo que se sabe sobre la distribución del tegumento en los tiranosáuridos. A pesar de una probabilidad del 89% de que los tiranosáuridos comenzaran con plumas, determinaron que los tiranosáuridos escamosos tienen una probabilidad del 97% de ser cierto. Los datos "proporcionan evidencia convincente de una cubierta completamente escamosa en Tyrannosaurus", escribió el equipo, aunque admitieron que el plumaje todavía puede haber estado presente en la región dorsal donde aún no se han encontrado impresiones de piel. Bell et al. plantea la hipótesis de que las impresiones de escamas de los tiranosáuridos son posiblemente retículos que se derivan secundariamente de las plumas, aunque se necesitan pruebas para respaldar esto. [88] Sin embargo, otros argumentan que esto se debe al sesgo tafonómico en los tiranosáuridos. [93]
Aún no se ha determinado por qué pudo haber ocurrido tal cambio tegumentario. Un precedente de pérdida de plumas se puede ver en otros grupos de dinosaurios como los ornitisquios , en los que se perdieron las estructuras filamentosas y reaparecieron las escamas. [94] Aunque se ha sugerido el gigantismo como un mecanismo, Phil R. Bell, coautor del estudio, señaló que el Yutyrannus emplumado se superponía en tamaño con Gorgosaurus y Albertosaurus . "El problema aquí es que tenemos grandes tiranosaurios, algunos con plumas, otros sin ellas, que viven en climas bastante similares. Entonces, ¿cuál es la razón de esta diferencia? Realmente no lo sabemos". [95]

Las cuencas oculares de Tyrannosaurus están posicionadas de manera que los ojos apuntarían hacia adelante, lo que les da una visión binocular ligeramente mejor que la de los halcones modernos . Mientras que los terópodos depredadores en general tenían visión binocular directamente frente a su cráneo, los tiranosaurios tenían un área de superposición significativamente mayor. Jack Horner también señaló que el linaje de los tiranosaurios tenía una historia de mejora constante de la visión binocular. Es difícil ver cómo la selección natural habría favorecido esta tendencia a largo plazo si los tiranosaurios hubieran sido carroñeros puros, que no habrían necesitado la percepción de profundidad avanzada que proporciona la visión estereoscópica . [96] [97] En los animales modernos, la visión binocular se encuentra principalmente en los depredadores (las principales excepciones son los primates , que la necesitan para saltar de rama en rama). A diferencia de Tyrannosaurus , Tarbosaurus tenía un cráneo más estrecho más típico de otros tiranosáuridos en los que los ojos miraban principalmente hacia los lados. Todo esto sugiere que Tarbosaurus dependía más de sus sentidos del olfato y el oído que de su vista. [98] En los especímenes de Gorgosaurus , la cuenca del ojo era circular en lugar de ovalada o con forma de ojo de cerradura como en otros géneros de tiranosáuridos. [11] En Daspletosaurus , esta era un óvalo alto, en algún lugar entre la forma circular vista en Gorgosaurus y la forma de "ojo de cerradura" de Tyrannosaurus . [10] [11] [44]
Basándose en comparaciones de la textura ósea de Daspletosaurus con los cocodrilos actuales , un estudio detallado en 2017 por Thomas D. Carr et al. encontró que los tiranosaurios tenían escamas grandes y planas en sus hocicos . [99] [100] En el centro de estas escamas había pequeños parches queratinizados . En los cocodrilos , estos parches cubren haces de neuronas sensoriales que pueden detectar estímulos mecánicos, térmicos y químicos . [101] [102] Propusieron que los tiranosaurios probablemente también tenían haces de neuronas sensoriales debajo de sus escamas faciales y pueden haberlas usado para identificar objetos, medir la temperatura de sus nidos y recoger suavemente huevos y crías . [99]

Las crestas óseas se encuentran en los cráneos de muchos terópodos, incluidos muchos tiranosáuridos. Alioramus , un posible tiranosáurido de Mongolia, tiene una sola fila de cinco protuberancias óseas prominentes en los huesos nasales; una fila similar de protuberancias mucho más bajas está presente en el cráneo de Appalachiosaurus , así como en algunos especímenes de Daspletosaurus , Albertosaurus y Tarbosaurus . [16] En Albertosaurus , Gorgosaurus y Daspletosaurus , hay un cuerno prominente frente a cada ojo en el hueso lagrimal. El cuerno lagrimal está ausente en Tarbosaurus y Tyrannosaurus , que en cambio tienen una cresta en forma de medialuna detrás de cada ojo en el hueso postorbital . Estas crestas de la cabeza pueden haber sido utilizadas para exhibición , tal vez para el reconocimiento de especies o el comportamiento de cortejo . [10]
Durante mucho tiempo se creyó que el Tyrannosaurus , como la mayoría de los dinosaurios, tenía un metabolismo reptil ectotérmico ("de sangre fría") , pero fue cuestionado por científicos como Robert T. Bakker y John Ostrom en los primeros años del " Renacimiento de los Dinosaurios ", que comenzó a fines de la década de 1960. [103] [104] Se afirmó que el propio Tyrannosaurus rex era endotérmico ("de sangre caliente"), lo que implica un estilo de vida muy activo. [105] Desde entonces, varios paleontólogos han buscado determinar la capacidad del Tyrannosaurus para regular su temperatura corporal . La evidencia histológica de altas tasas de crecimiento en jóvenes T. rex , comparables a las de los mamíferos y las aves, puede apoyar la hipótesis de un metabolismo alto. Las curvas de crecimiento indican que, como en los mamíferos y las aves, el crecimiento del T. rex se limitó principalmente a los animales inmaduros, en lugar del crecimiento indeterminado observado en la mayoría de los otros vertebrados . [50] Se ha indicado que la diferencia de temperatura puede no haber sido más de 4 a 5 °C (7 a 9 °F) entre las vértebras del torso y la tibia de la parte inferior de la pierna. Este pequeño rango de temperatura entre el núcleo del cuerpo y las extremidades fue afirmado por el paleontólogo Reese Barrick y el geoquímico William Showers para indicar que T. rex mantenía una temperatura corporal interna constante ( homeotermia ) y que disfrutaba de un metabolismo en algún lugar entre los reptiles ectotérmicos y los mamíferos endotérmicos. [106] Más tarde encontraron resultados similares en especímenes de Giganotosaurus , que vivieron en un continente diferente y decenas de millones de años antes en el tiempo. [107] Incluso si Tyrannosaurus rex muestra evidencia de homeotermia, no significa necesariamente que fuera endotérmico. Tal termorregulación también puede explicarse por la gigantotermia , como en algunas tortugas marinas actuales . [108] [109] [110]

En la Formación Dinosaur Park, Gorgosaurus vivió junto a una especie más rara del tiranosaurio Daspletosaurus . Este es uno de los pocos ejemplos de dos géneros de tiranosaurios coexistiendo. Los depredadores de tamaño similar en los gremios de depredadores modernos están separados en diferentes nichos ecológicos por diferencias anatómicas, de comportamiento o geográficas que limitan la competencia. La diferenciación de nichos entre los tiranosáuridos de Dinosaur Park no se entiende bien. [111] En 1970, Dale Russell planteó la hipótesis de que el más común Gorgosaurus cazaba activamente hadrosaurios de pies ligeros , mientras que los ceratopsianos y anquilosaurios más raros y problemáticos (dinosaurios con cuernos y fuertemente acorazados ) se dejaban al Daspletosaurus de complexión más pesada . [12] Sin embargo, un espécimen de Daspletosaurus (OTM 200) de la contemporánea Formación Two Medicine de Montana conserva los restos digeridos de un hadrosaurio juvenil en su región intestinal. [112] A diferencia de otros grupos de dinosaurios, ninguno de los géneros era más común en elevaciones más altas o más bajas que el otro. [111] Sin embargo, Gorgosaurus parece ser más común en formaciones del norte como el Dinosaur Park, con especies de Daspletosaurus más abundantes al sur. El mismo patrón se observa en otros grupos de dinosaurios. Los ceratopsianos chasmosaurinos y los hadrosaurios hadrosaurinos también son más comunes en la Formación Two Medicine de Montana y en el suroeste de América del Norte durante el Campaniano, mientras que los centrosaurinos y los lambeosaurinos dominan en latitudes del norte. Holtz ha sugerido que este patrón indica preferencias ecológicas compartidas entre tiranosaurinos, chasmosaurinos y hadrosaurinos. Al final de la etapa tardía del Maastrichtiano, los tiranosaurios como el Tyrannosaurus rex , los hadrosaurinos como el Edmontosaurus y los chasmosaurinos como el Triceratops estaban muy extendidos por todo el oeste de América del Norte, mientras que los albertosaurinos y los centrosaurinos se extinguieron y los lambeosaurinos eran raros. [10]
Hay evidencia limitada de comportamiento social entre los tiranosáuridos. Los investigadores informaron que se encontraron un esqueleto de subadulto y uno juvenil en la misma cantera que el espécimen "Sue", lo que se ha utilizado para apoyar la hipótesis de que los tiranosáuridos pueden haber vivido en grupos sociales de algún tipo. [113] Si bien no hay evidencia de comportamiento gregario en Gorgosaurus , [61] [62] hay evidencia de algún comportamiento de manada en Albertosaurus y Daspletosaurus .
Un ejemplar joven de la especie Dinosaur Park Daspletosaurus ( TMP 94.143.1) muestra marcas de mordeduras en la cara que fueron infligidas por otro tiranosaurio. Las marcas de mordeduras están curadas, lo que indica que el animal sobrevivió a la mordedura. Un Dinosaur Park Daspletosaurus (TMP 85.62.1) adulto también exhibe marcas de mordeduras de tiranosaurio, lo que demuestra que los ataques en la cara no se limitaban a animales más jóvenes. Si bien es posible que las mordeduras fueran atribuibles a otras especies, la agresión intraespecífica, incluidas las mordeduras faciales, es muy común entre los depredadores. Las mordeduras faciales se observan en otros tiranosaurios como Gorgosaurus y Tyrannosaurus , así como en otros géneros de terópodos como Sinraptor y Saurornitholestes . Darren Tanke y Phil Currie plantean la hipótesis de que las mordeduras se deben a la competencia intraespecífica por territorio o recursos, o por el dominio dentro de un grupo social. [61]

La evidencia de que Daspletosaurus vivía en grupos sociales proviene de un lecho de huesos encontrado en la Formación Two Medicine de Montana. El lecho de huesos incluye los restos de tres Daspletosaurus , incluido un adulto grande, un juvenil pequeño y otro individuo de tamaño intermedio. Al menos cinco hadrosaurios se conservan en el mismo lugar. La evidencia geológica indica que los restos no fueron reunidos por las corrientes del río , sino que todos los animales fueron enterrados simultáneamente en el mismo lugar. Los restos de hadrosaurios están dispersos y tienen muchas marcas de dientes de tiranosaurio, lo que indica que Daspletosaurus se estaba alimentando de los hadrosaurios en el momento de la muerte. La causa de la muerte es desconocida. Currie especula que los daspletosaurios formaban una manada , aunque esto no se puede afirmar con certeza. [62] Otros científicos son escépticos sobre la evidencia de grupos sociales en Daspletosaurus y otros grandes terópodos; [114] Brian Roach y Daniel Brinkman han sugerido que la interacción social del Daspletosaurus se habría parecido más al moderno dragón de Komodo , donde los individuos no cooperativos se agolpan en los cadáveres, atacándose con frecuencia e incluso canibalizándose entre sí en el proceso. [115]
El yacimiento de huesos de Dry Island descubierto por Barnum Brown y su equipo contiene los restos de 22 Albertosaurus , la mayor cantidad de individuos encontrados en una localidad de cualquier terópodo del Cretácico, y la segunda mayor cantidad de cualquier dinosaurio terópodo grande detrás del conjunto Allosaurus en la cantera de dinosaurios Cleveland-Lloyd en Utah . El grupo parece estar compuesto por un adulto muy viejo; ocho adultos entre 17 y 23 años; siete subadultos que atraviesan sus fases de crecimiento rápido entre los 12 y 16 años de edad; y seis juveniles entre las edades de 2 y 11 años, que aún no habían alcanzado la fase de crecimiento. [53] La casi ausencia de restos de herbívoros y el estado similar de conservación entre los muchos individuos en la cantera del yacimiento de huesos de Albertosaurus llevaron a Phil Currie a concluir que la localidad no era una trampa para depredadores como los pozos de alquitrán de La Brea en California , y que todos los animales preservados murieron al mismo tiempo. Currie afirma esto como evidencia del comportamiento de manada. [116] Otros científicos son escépticos y observan que los animales pueden haber sido obligados a unirse por la sequía, las inundaciones o por otras razones. [53] [114] [117]
Aunque en general sigue siendo controvertido, existe evidencia que apoya la teoría de que al menos algunos tiranosáuridos eran sociales. En la Formación Wapiti de Columbia Británica , un proveedor local llamado Aaron Fredlund descubrió un rastro compuesto por las huellas de tres tiranosáuridos individuales (llamados icnogénero Bellatoripes fredlundi ) y lo describió en la revista PLOS One Richard McCrea et al. Un examen del rastro no encontró evidencia de que se hubiera dejado un rastro mucho después de que se hubiera hecho otro, lo que respalda aún más la hipótesis de que tres tiranosáuridos individuales viajaban juntos como grupo. Investigaciones posteriores revelaron que los animales viajaban a una velocidad de entre 3,9 y 5,2 mph (6,3 y 8,4 km/h) y probablemente tenían una altura de cadera de alrededor de 7 a 9 pies. Como se conocen tres géneros diferentes de tiranosáuridos ( Gorgosaurus , Daspletosaurus y Albertosaurus , respectivamente) de la formación, se desconoce qué género fue el creador de la huella. [118] [119] [120] Evidencia adicional en forma de un lecho de huesos de la cantera Rainbows and Unicorns en la Formación Kaiparowits del sur de Utah descrita en 2021 atribuida a Teratophoneus sugiere que otros tiranosáuridos también eran animales sociales. Los fósiles, que consisten en cuatro o posiblemente cinco animales diferentes de entre 4 y 22 años de edad, sugieren un evento de mortalidad masiva, posiblemente causado por inundaciones, o menos probablemente por toxicosis cianobacteriana, incendio o sequía. El hecho de que todos los animales preservados parecieran haber perecido en un corto lapso de tiempo refuerza aún más el argumento del comportamiento gregario en los tiranosáuridos, con los lechos óseos de géneros como Teratophoneus , Albertosaurus , Tyrannosaurus y Daspletosaurus mostrando que el comportamiento social sugerido puede haber sido generalizado entre los tiranosáuridos en general. [121] [122] [123]
Las marcas de dientes de tiranosaurio son los rastros de alimentación más comúnmente preservados de los dinosaurios carnívoros. [124] Se han reportado en ceratopsianos , hadrosaurios y otros tiranosaurios. [124] Los huesos de tiranosáuridos con marcas de dientes representan aproximadamente el 2% de los fósiles conocidos con marcas de dientes preservadas. [124] Los dientes de tiranosáurido se usaban como puntos de sujeción para arrancar carne de un cuerpo, en lugar de funciones de corte similares a las de un cuchillo . [125] Los patrones de desgaste de los dientes sugieren que los complejos comportamientos de sacudidas de cabeza pueden haber estado involucrados en la alimentación de los tiranosáuridos. [125]

Algunos investigadores especularon sobre los hábitos de caza en grupo de Albertosaurus y sugirieron que los miembros más jóvenes de la manada pueden haber sido los responsables de conducir a sus presas hacia los adultos, que eran más grandes y poderosos, pero también más lentos. [116] Los juveniles también pueden haber tenido estilos de vida diferentes a los adultos, llenando nichos de depredadores entre los de los enormes adultos y los terópodos contemporáneos más pequeños, los más grandes de los cuales eran dos órdenes de magnitud más pequeños que un Albertosaurus adulto en masa. [10] Sin embargo, como la preservación del comportamiento en el registro fósil es extremadamente rara, estas ideas no se pueden probar fácilmente. Phil Currie especula que Daspletosaurus formaba manadas para cazar , aunque esto no se puede afirmar con certeza. [62] No hay evidencia de tal comportamiento gregario en Gorgosaurus . [61] [62]
El debate sobre si el Tyrannosaurus era un depredador o un carroñero puro es tan antiguo como el debate sobre su locomoción. Lambe (1917) describió un buen esqueleto de un pariente cercano del Tyrannosaurus , el Gorgosaurus , y concluyó que éste y, por lo tanto, también el Tyrannosaurus eran carroñeros puros, porque los dientes del Gorgosaurus apenas mostraban desgaste. [126] Este argumento ya no se toma en serio, porque los terópodos reemplazaban sus dientes con bastante rapidez. Desde el primer descubrimiento del Tyrannosaurus, la mayoría de los científicos han estado de acuerdo en que era un depredador, aunque, al igual que los grandes depredadores modernos, habría estado feliz de carroñear o robar la presa de otro depredador si hubiera tenido la oportunidad. [127] [128]
El conocido experto en hadrosaurios Jack Horner es actualmente el principal defensor de la idea de que el Tyrannosaurus era exclusivamente un carroñero y no se dedicaba a la caza activa en absoluto. [66] [129] [130] Horner ha presentado varios argumentos para apoyar la hipótesis del carroñero puro. La presencia de grandes bulbos olfatorios y nervios olfativos sugiere un sentido del olfato altamente desarrollado para olfatear cadáveres a grandes distancias. Los dientes podían triturar el hueso y, por lo tanto, podían extraer la mayor cantidad de alimento ( médula ósea ) posible de los restos de cadáveres, generalmente las partes menos nutritivas. Al menos algunas de sus presas potenciales podían moverse rápidamente, mientras que la evidencia sugiere que el Tyrannosaurus caminaba en lugar de correr. [129] [131]
Otra evidencia sugiere un comportamiento de caza en Tyrannosaurus . Las cuencas oculares de los tiranosaurios están posicionadas de manera que los ojos apunten hacia adelante, lo que les da una visión binocular ligeramente mejor que la de los halcones modernos . Se han encontrado daños infligidos por tiranosaurios en esqueletos de hadrosaurios y Triceratops que parecían haber sobrevivido a los ataques iniciales. [132] [133] [134] Algunos investigadores argumentan que si Tyrannosaurus fuera un carroñero, otro dinosaurio tenía que ser el principal depredador en el Cretácico Superior amerasiático. Las principales presas eran los marginocéfalos y ornitópodos más grandes . Los otros tiranosáuridos comparten tantas características con Tyrannosaurus que solo los pequeños dromaeosaurios permanecen como posibles depredadores superiores. En vista de esto, los partidarios de la hipótesis del carroñero han sugerido que el tamaño y el poder de los tiranosaurios les permitían robar presas a depredadores más pequeños. [131]

En 2023, se informó de un Gorgosaurus juvenil con su contenido estomacal in situ que contenía dos juveniles de Citipes de aproximadamente un año intactos en la Formación Dinosaur Park . Este juvenil habría tenido entre 5 y 7 años en el momento de la muerte, midiendo unos 4 metros (13 pies) de largo y pesando alrededor de 335 kilogramos (739 libras). Es mucho más grande que los dos juveniles de Citipes que pesan alrededor de 9-12 kilogramos (20-26 libras), contrariamente a la suposición de que los tiranosáuridos se alimentaban de presas de su tamaño una vez que alcanzaban los 16-32 kilogramos (35-71 libras), lo que indica que los tiranosáuridos juveniles todavía consumían presas mucho más pequeñas después de superar un cierto umbral de tamaño. El descubrimiento de este espécimen indicó que los tiranosáuridos probablemente no cazaban en manadas multigeneracionales, ya que el tamaño de sus presas es demasiado pequeño para compartir con los congéneres. También es una evidencia dietética directa que refuerza la teoría del "cambio dietético ontogénico" de los tiranosáuridos, como se infirió previamente mediante modelos ecológicos y características anatómicas entre diferentes grupos de edad. Solo los restos de las extremidades traseras y las vértebras caudales de Citipes juveniles estaban presentes en la cavidad estomacal del tiranosáurido, lo que sugiere que un Gorgosaurus juvenil puede haber tenido un consumo preferencial de las extremidades traseras musculares. [135]
La evidencia también sugiere fuertemente que los tiranosáuridos eran al menos ocasionalmente caníbales. El propio Tyrannosaurus tiene evidencia sólida que apunta a que fue caníbal al menos en una capacidad de carroñeo basada en marcas de dientes en los huesos del pie, el húmero y los metatarsianos de un espécimen. [136] Los fósiles de la Formación Fruitland , la Formación Kirtland (ambas del Campaniense) y la Formación Ojo Alamo del Maastichtiense sugieren que el canibalismo estaba presente en varios géneros de tiranosáuridos de la Cuenca de San Juan. La evidencia reunida de los especímenes sugiere un comportamiento de alimentación oportunista en tiranosáuridos que canibalizaban a miembros de su propia especie. [137]

Los restos de tiranosaurio más antiguos que se conocen se encontraron hace entre 129,4 y 125 millones de años en Ishikawa, Japón, cerca de Lat 36.166668 lng 136.633331. Marsh los encontró en 1881 e HF Osborne los identificó en 1906; la edad de los restos fue determinada por H. Matsuoka et al. en 2002.
Aunque los primeros tiranosáuridos se encuentran en los tres continentes del norte, los fósiles de tiranosáuridos sólo se conocen en América del Norte y Asia. A veces, los restos fragmentarios descubiertos en el hemisferio sur se han descrito como "tiranosáuridos del hemisferio sur", aunque parece que se trata de fósiles de abelisáuridos mal identificados . [138]
Los restos de tiranosáuridos nunca han sido recuperados del este de América del Norte, mientras que tiranosáuridos más basales, como Dryptosaurus y Appalachiosaurus , persistieron allí hasta el final del Cretácico, lo que indica que los tiranosáuridos deben haber evolucionado o dispersado en el oeste de América del Norte después de que el continente fuera dividido en dos por el Mar Interior Occidental a mediados del Cretácico. [16] Se han encontrado fósiles de tiranosáuridos en Alaska , que pueden haber proporcionado una ruta para la dispersión entre América del Norte y Asia. [139] Se encontró que Alioramus y Tarbosaurus están relacionados en un análisis cladístico, formando una rama asiática única de la familia. [18] Esto fue refutado más tarde con el descubrimiento de Qianzhousaurus y la descripción de la familia de tiranosáuridos Alioramini. Los dientes de tiranosáuridos de una gran especie de variedad desconocida fueron descubiertos en la península de Nagasaki por investigadores del Museo de Dinosaurios de la Prefectura de Fukui, expandiendo aún más el rango del grupo. Se estima que los dientes tienen una antigüedad de 81 millones de años ( Edad Campaniana ). [140]

De las dos subfamilias, los tiranosaurios parecen haber estado más extendidos. Los albertosaurinos son desconocidos en Asia, que fue el hogar de los tiranosaurios, como Tarbosaurus y Zhuchengtyrannus , y Qianzhousaurus y Alioramus de los Alioramini. Tanto las subfamilias Tyrannosaurinae como Albertosaurinae estuvieron presentes en las etapas Campaniense y Maastrichtiano temprano de América del Norte, con tiranosaurios como Daspletosaurus distribuyéndose por todo el interior occidental, mientras que los albertosaurinos Albertosaurus y Gorgosaurus actualmente se conocen solo de la parte noroeste del continente. [141]
A finales del Maastrichtiano, los albertosaurinos parecen haberse extinguido, mientras que el tiranosaurino Tyrannosaurus vagó desde Saskatchewan hasta Texas . Este patrón se refleja en otros taxones de dinosaurios de América del Norte. Durante el Campaniense y principios del Maastrichtiano, los hadrosaurios lambeosaurinos y los ceratopsianos centrosaurinos son comunes en el noroeste, mientras que los hadrosaurinos y los chasmosaurinos eran más comunes en el sur. A finales del Cretácico, los centrosaurinos son desconocidos y los lambeosaurinos son raros, mientras que los hadrosaurinos y los chasmosaurinos eran comunes en todo el interior occidental. [10] Un estudio publicado en la revista Scientific Reports el 2 de febrero de 2016, por Steve Brusatte, Thomas Carr et al . indica que durante el Maastrichtiano tardío, el propio Tyrannosaurus podría haber sido parcialmente responsable de la extinción de los otros tiranosáuridos en la mayor parte del oeste de América del Norte. El estudio indica que Tyrannosaurus podría haber sido un inmigrante de Asia en lugar de haber evolucionado en América del Norte (posiblemente un descendiente del estrechamente relacionado Tarbosaurus ) que suplantó y superó a otros tiranosáuridos. Esta teoría se ve apoyada además por el hecho de que pocos o ningún otro tipo de tiranosáurido se encuentran dentro del área de distribución conocida de Tyrannosaurus . [142]
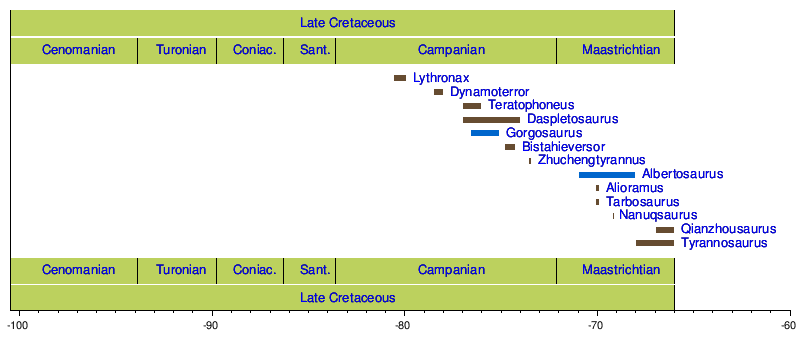
{{cite journal}}: CS1 maint: varios nombres: lista de autores ( enlace )Las longevidades muestreadas para T. rex oscilaron entre 2 y 28 años.(Fe de erratas: doi : 10.1038/nature16487, PMID 26675726, Retraction Watch ) y las estimaciones de masa corporal correspondientes oscilaron entre 29,9 y 5654 kg.
{{cite journal}}: CS1 maint: varios nombres: lista de autores ( enlace ){{cite journal}}: CS1 maint: varios nombres: lista de autores ( enlace ){{cite web}}: CS1 maint: bot: estado de URL original desconocido ( enlace ){{cite journal}}: CS1 maint: varios nombres: lista de autores ( enlace ){{cite web}}: CS1 maint: copia archivada como título ( enlace ){{cite web}}: CS1 maint: nombres numéricos: lista de autores ( enlace )[1]