Los protistas unicelulares comprenden un vasto y diverso grupo de organismos que cubre prácticamente todos los ambientes y hábitats, mostrando una variedad de formas y figuras. Cientos de especies del género ciliado Paramecium [3] o del flagelado Euglena [4] se encuentran en reservorios marinos, salobres y de agua dulce; el alga verde Chlamydomonas se distribuye en el suelo y el agua dulce en todo el mundo; [5] los parásitos del género Giardia colonizan los intestinos de varias especies de vertebrados. [6] Una de las características compartidas de estos organismos es su motilidad, crucial para la adquisición de nutrientes y la evitación de peligros. [7] En el proceso de evolución, los organismos unicelulares se han desarrollado en una variedad de direcciones, y por lo tanto su rica morfología da como resultado un amplio espectro de modos de natación. [8] [2]
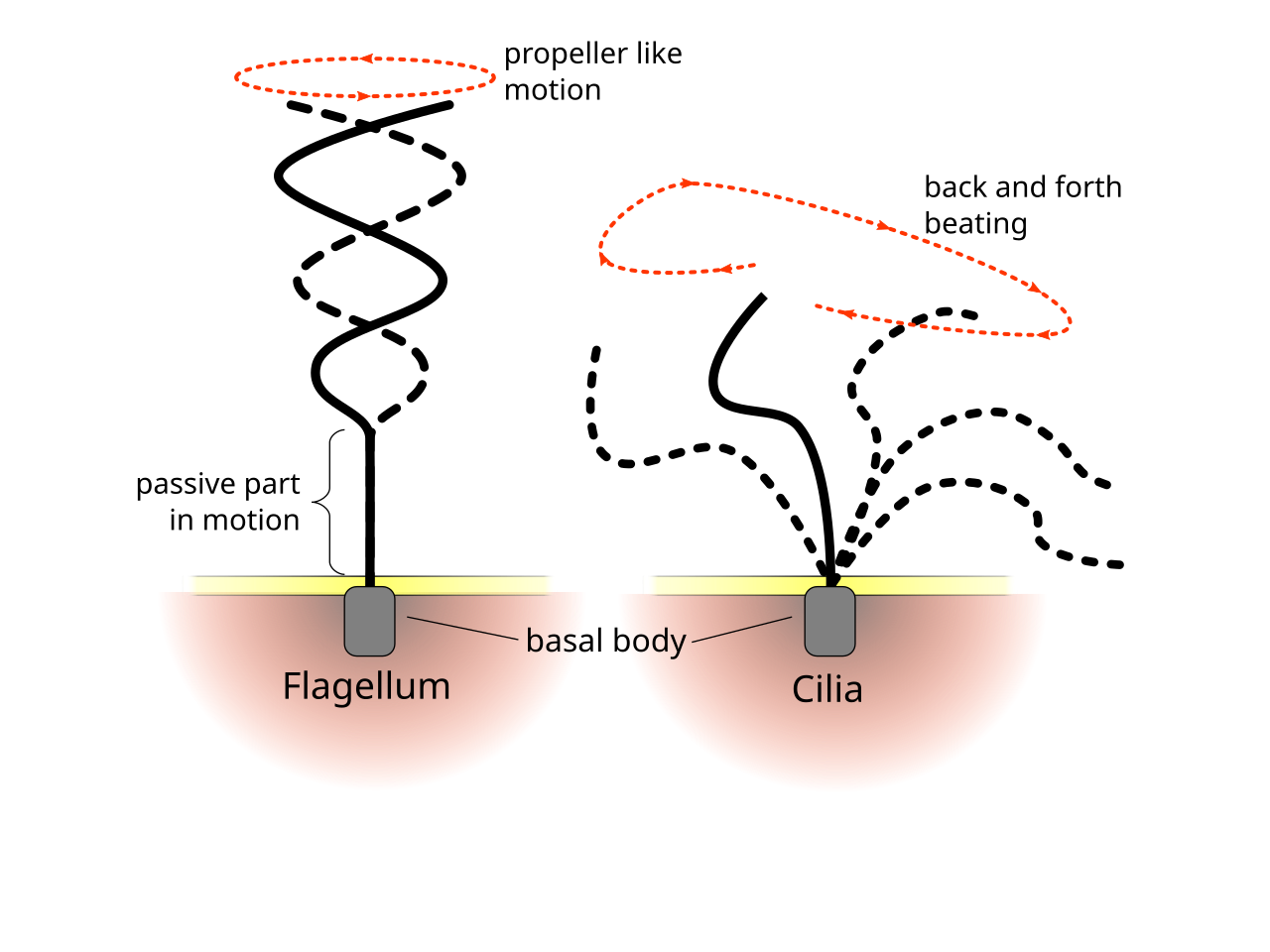
Aunque los flagelos protistas tienen una diversidad de formas y funciones, [11] dos grandes familias, flagelados y ciliados , se pueden distinguir por la forma y el patrón de batido de sus flagelos. [2]
En el árbol filogenético de la derecha, los organismos acuáticos (que viven en ambientes marinos, salobres o de agua dulce) tienen sus ramas dibujadas en azul, mientras que los organismos parásitos tienen sus ramas dibujadas en rojo. Los ciliados están indicados con un asterisco después de sus nombres. Para cada filo marcado en negrita, se ha dibujado un organismo representativo junto a su nombre. [2]
Modos de locomoción
Flagelados
Diferencia en el patrón de batido del flagelo y el cilio
Los flagelos se utilizan tanto en procariotas (arqueas y bacterias) como en protistos. Además, tanto los flagelos como los cilios se utilizan ampliamente en células eucariotas (vegetales y animales), aparte de los protistos.
Los patrones regulares de batido de los cilios y flagelos eucariotas generan movimiento a nivel celular. Los ejemplos van desde la propulsión de células individuales, como la natación de los espermatozoides , hasta el transporte de líquido a lo largo de una capa estacionaria de células, como en un tracto respiratorio . Aunque los flagelos eucariotas y los cilios móviles son ultraestructuralmente idénticos, el patrón de batido de los dos orgánulos puede ser diferente. En el caso de los flagelos, el movimiento suele ser plano y ondulatorio, mientras que los cilios móviles suelen realizar un movimiento tridimensional más complicado con un movimiento de potencia y recuperación.
Los flagelos eucariotas (de células animales, vegetales y protistas) son proyecciones celulares complejas que se mueven de un lado a otro. Los flagelos eucariotas se clasifican junto con los cilios móviles eucariotas como undulipodios [17] para enfatizar su papel distintivo de apéndice ondulado en la función celular o la motilidad . Los cilios primarios son inmóviles y no son undulipodios.
Flagelados marinos de los géneros (de izquierda a derecha) Cryptaulax , Abollifer , Bodo , Rhynchomonas , Kittoksia , Allas y Metromonas [18]
Los flagelados suelen tener una pequeña cantidad de flagelos largos distribuidos a lo largo de sus cuerpos, y los activan para generar impulso. El conjunto de secuencias de movimiento observadas incluye ondas ondulatorias planas y ondas helicoidales que viajan desde la base hasta la punta, o en la dirección opuesta. [19] [20] Los flagelos unidos al mismo cuerpo pueden seguir diferentes patrones de batido, lo que conduce a una estrategia de locomoción compleja que a menudo depende también de la resistencia que el cuerpo celular plantea al fluido. [2]
Ciliados
Cilia realiza potentes movimientos hacia adelante con un flagelo rígido seguido de un movimiento de recuperación relativamente lento con un flagelo relajado.
A diferencia de los flagelados, la propulsión de los ciliados se deriva del movimiento de una capa de cilios densamente agrupados y en movimiento colectivo, que son flagelos cortos similares a pelos que cubren sus cuerpos. El artículo de revisión seminal de Brennen y Winet (1977) enumera algunos ejemplos de ambos grupos, destacando su forma, forma de batir, características geométricas y propiedades de natación. [20] Los cilios también pueden usarse para el transporte del fluido circundante, y su cooperatividad puede conducir a la generación de flujo dirigido. En organismos superiores, esto puede ser crucial para los procesos de transporte interno, como en el flujo citoplasmático dentro de las células vegetales, [21] o el transporte de óvulos desde el ovario hasta el útero en los mamíferos hembra. [22] [2]
Los ciliados generalmente tienen cientos o miles de cilios que están densamente empaquetados en formaciones. Al igual que los flagelos, los cilios son impulsados por motores moleculares especializados . Un movimiento eficiente hacia adelante se realiza con un flagelo rígido, seguido de un movimiento ineficiente hacia atrás realizado con un flagelo relajado. Durante el movimiento, un cilio individual se deforma a medida que utiliza los movimientos de potencia de alta fricción y los movimientos de recuperación de baja fricción. Dado que hay múltiples cilios empaquetados juntos en un organismo individual, muestran un comportamiento colectivo en un ritmo metacrónico . Esto significa que la deformación de un cilio está en fase con la deformación de su vecino, lo que causa ondas de deformación que se propagan a lo largo de la superficie del organismo. Estas ondas de propagación de cilios son las que permiten al organismo utilizar los cilios de manera coordinada para moverse. Un ejemplo típico de un microorganismo ciliado es el Paramecium , un protozoo ciliado unicelular cubierto por miles de cilios. Los cilios que se mueven juntos permiten al Paramecium impulsarse a través del agua a velocidades de 500 micrómetros por segundo. [23]
Cilios latiendo en un micronadador multiciliado [24]
Dos imágenes en momentos diferentes, una después de la otra, que muestran diferentes disposiciones de los cilios en movimiento. El color variable de los cilios indica la etapa instantánea del ciclo de movimiento. El movimiento del nadador es de izquierda a derecha.
La tercera forma predominante de movilidad celular protista es la migración celular dependiente de actina . La evolución de la natación basada en flagelos ha sido bien estudiada y hay evidencia sólida que sugiere que el origen evolutivo único del flagelo eucariota ocurrió antes de la diversificación de los eucariotas modernos. Por otro lado, el reptar dependiente de actina utiliza muchos mecanismos moleculares diferentes y el estudio de cómo evolucionaron recién está comenzando. [25]
(a) Colonia de dieciséis células. Cada célula tiene dos flagelos de 30–40 μm de largo. La barra de escala mide 10 μm. (b) Esquema de una colonia de radio a: dieciséis células (verdes) cada una con una mancha ocular (punto naranja). El flagelo cis es el más cercano a la mancha ocular, el flagelo trans es el más alejado. [27] Los flagelos de las células centrales baten en una brazada opuesta, mientras que los flagelos periféricos baten en paralelo. La organización en rueda de molino de los flagelos periféricos conduce a una rotación del cuerpo hacia la izquierda a una velocidad ω3.
Gonium es un género de algas coloniales perteneciente a la familia Volvocaceae . Las colonias típicas tienen de 4 a 16 células, todas del mismo tamaño, dispuestas en una placa plana, sin diferenciación anteroposterior. En una colonia de 16 células, cuatro están en el centro y las otras 12 en los cuatro lados, tres en cada uno. [28]
Desde el trabajo de August Weismann sobre la teoría del plasma germinal en biología [29] y de Julian Huxley sobre la naturaleza del individuo en la teoría evolutiva, [30] las diversas especies de algas verdes pertenecientes a la familia Volvocaceae han sido reconocidas como importantes en el estudio de las transiciones evolutivas de la vida unicelular a la multicelular. En una visión biológica moderna, [31] esta importancia surge de una serie de características específicas de estas algas, incluido el hecho de que son una familia existente (evitando la necesidad de estudiar microfósiles ), son fácilmente obtenibles en la naturaleza, han sido estudiadas desde una variedad de perspectivas (bioquímica, de desarrollo, genética) y han tenido estudios ecológicos significativos. Desde una perspectiva de dinámica de fluidos, [32] su tamaño relativamente grande y las condiciones fáciles de cultivo permiten estudios precisos de su motilidad, los flujos que crean con sus flagelos y las interacciones entre organismos, mientras que su alto grado de simetría simplifica las descripciones teóricas de esos mismos fenómenos. [33] [26]
Como son fotosintéticas, la capacidad de estas algas para ejecutar fototaxis es central para su vida. Debido a que el linaje abarca desde unicelulares hasta grandes formas coloniales, se puede utilizar para estudiar la evolución de la coordinación multicelular de la motilidad. La motilidad y la fototaxis de las algas verdes móviles han sido objeto de una extensa literatura en los últimos años, [34] [ 35 ] [36] [37] [38] [39] [40] centrándose principalmente en los dos casos extremos: Chlamydomonas unicelular y Volvox mucho más grande , con especies compuestas de 1000–50,000 células. Chlamydomonas nada típicamente mediante la activación de sus dos flagelos en una brazada, combinando propulsión y rotación lenta del cuerpo. Posee una mancha ocular, una pequeña área altamente sensible a la luz, [41] [42] que activa los dos flagelos de manera diferente. [43] Estas respuestas son adaptativas, en una escala de tiempo que coincide con el período de rotación del cuerpo celular, [44] [45] [46] y permiten a las células explorar el entorno y nadar hacia la luz. [47] El Volvox multicelular muestra un mayor nivel de complejidad, con diferenciación entre células germinales internas y células somáticas dedicadas a la propulsión. A pesar de carecer de un sistema nervioso central para coordinar sus células, el Volvox exhibe una fototaxis precisa. Esto también se logra mediante una respuesta adaptativa a los niveles cambiantes de luz, con un tiempo de respuesta ajustado al período de rotación de la colonia que crea una respuesta diferencial entre los lados claro y oscuro del esferoide. [35] [48] [26]
A la luz de lo anterior, una pregunta natural es la siguiente: ¿cómo logra la fototaxis el organismo diferenciado más simple? En el linaje de los volvocinos, la especie de interés es Gonium. Esta colonia de 8 o 16 células representa uno de los primeros pasos hacia la verdadera multicelularidad, [49] se presume que evolucionó a partir del ancestro común unicelular antes que otras algas volvocinas. [50] También es la primera en mostrar diferenciación celular. [26]
La colonia de 16 células de Gonium que se muestra en el diagrama de la derecha está organizada en dos cuadrados concéntricos de 4 y 12 células respectivamente, cada una biflagelada, unidas por una matriz extracelular. [51] Todos los flagelos apuntan hacia el mismo lado: exhibe una simetría mucho menor que Volvox, careciendo de simetría anteroposterior. Sin embargo, realiza funciones similares a sus contrapartes unicelulares y de colonias grandes, ya que mezcla propulsión y rotación corporal y nada eficientemente hacia la luz. [34] [52] [53] La organización flagelar de las células internas y periféricas difiere profundamente: [54] [55] Las células centrales son similares a Chlamydomonas, con los dos flagelos batiendo en una brazada opuesta, y contribuyen principalmente a la propulsión hacia adelante de la colonia. Las células en la periferia, sin embargo, tienen flagelos batiendo en paralelo, de una manera similar a las células Volvox . [27] Esto minimiza las interacciones estéricas y evita que los flagelos se crucen entre sí. [34] Además, estos flagelos están implantados con un ligero ángulo y organizados en forma de rueda de molino [ver Fig. 1(b)]: [54] Su movimiento induce una rotación hacia la izquierda de la colonia, resaltada en las Figs. 1(c) y 1(d) y en la Película suplementaria 1 [29]. Por lo tanto, la estructura de los flagelos de Gonium refuerza su posición clave como intermediario en la evolución hacia la multicelularidad y la diferenciación celular. [26]
Estos pequeños conjuntos planos muestran una natación intrigante a lo largo de trayectorias helicoidales (con su plano corporal casi normal a la dirección de natación) que han atraído la atención de los naturalistas desde el siglo XVIII. [52] [53] [56] Sin embargo, la forma en que las colonias de Gonium sesgan su natación hacia la luz sigue siendo poco clara. Las primeras observaciones microscópicas han identificado la actividad flagelar diferencial entre los lados iluminados y sombreados de la colonia como la fuente de la reorientación fototáctica. [52] [53] Sin embargo, aún falta una descripción completa de la dinámica de fluidos, que vincule cuantitativamente la respuesta flagelar a las variaciones de luz y las fuerzas y pares hidrodinámicos que actúan sobre la colonia. Desde una perspectiva evolutiva, la fototaxis en Gonium plantea cuestiones fundamentales como el grado en que la estrategia fototáctica del ancestro unicelular se conserva en la forma colonial, cómo la reacción fototáctica de los flagelos se adaptó a la geometría y simetría de la colonia y cómo conduce a una reorientación efectiva. [26]
Taxis protistas: movimiento dirigido
Fototaxis
Distribución de la fototaxis tridimensional en el árbol de eucariotas [57]
Las flechas rojas indican el punto de origen probable de la fototaxis en un grupo determinado. Los signos de interrogación indican incertidumbres respecto de su origen independiente o común.
Algunos protistas pueden moverse hacia o desde un estímulo, un movimiento conocido como taxis . Por ejemplo, el movimiento hacia la luz, denominado fototaxis , se logra acoplando su estrategia de locomoción con un órgano sensor de luz. [58] Los eucariotas desarrollaron por primera vez en la historia de la vida la capacidad de seguir la dirección de la luz en tres dimensiones en aguas abiertas. La estrategia de integración sensorial eucariota, el procesamiento sensorial y la velocidad y mecánica de las respuestas tácticas es fundamentalmente diferente de la que se encuentra en los procariotas. [59] [57]
Tanto los organismos fototácticos eucariotas unicelulares como los multicelulares tienen una forma fija, están polarizados, nadan en espiral y utilizan cilios para nadar y dirigirse fototácticamente. La señalización puede ocurrir a través de corrientes iónicas activadas por luz directa , adenilil ciclasas o proteínas G triméricas . Los fotorreceptores utilizados también pueden ser muy diferentes (ver más abajo). Sin embargo, la señalización en todos los casos eventualmente modifica la actividad de batido de los cilios. [57] La mecánica de la orientación fototáctica es análoga en todos los eucariotas. Un fotosensor con un ángulo de visión restringido gira para escanear el espacio y envía señales periódicamente a los cilios para alterar su batido, lo que cambiará la dirección de la trayectoria de natación helicoidal . La fototaxis tridimensional se puede encontrar en cinco de los seis grupos eucariotas principales ( opistocontos , amoebozoos , plantas , cromalveolados , excavatos , rizaria ). [57]
La fototaxis pelágica está presente en las algas verdes – no está presente en las algas glaucófitas o algas rojas . [57] Las algas verdes tienen un "estigma" ubicado en la porción más externa del cloroplasto , directamente debajo de las dos membranas del cloroplasto . El estigma está formado por decenas a varios cientos de glóbulos lipídicos , que a menudo forman matrices hexagonales y pueden estar dispuestos en una o más filas. Los glóbulos lipídicos contienen una mezcla compleja de pigmentos carotenoides , que proporcionan la función de cribado y el color rojo anaranjado, [60] así como proteínas que estabilizan los glóbulos. [61] El estigma está ubicado lateralmente, en un plano fijo en relación con los cilios, pero no directamente adyacente a los cuerpos basales. [62] [63] La posición fija está asegurada por la unión del cloroplasto a una de las raíces ciliares. [64] El estigma pigmentado no debe confundirse con el fotorreceptor. El estigma sólo proporciona sombreado direccional para los fotorreceptores adyacentes insertados en la membrana (por lo tanto, el término "mancha ocular" es engañoso). Los estigmas también pueden reflejar y enfocar la luz como un espejo cóncavo, mejorando así la sensibilidad. [57]
En el alga verde mejor estudiada, Chlamydomonas reinhardtii , la fototaxis está mediada por un pigmento de rodopsina , como se demostró por primera vez con la restauración del fotocomportamiento normal en un mutante ciego por análogos del cromóforo retiniano . [65] Dos rodopsinas de tipo arqueobacteriano, canalrodopsina -1 y -2, [66] [67] fueron identificadas como receptores de fototaxis en Chlamydomonas . [68] Ambas proteínas tienen una porción transmembrana 7-terminal N, similar a las rodopsinas arqueobacterianas, seguida de una porción asociada a la membrana C-terminal de aproximadamente 400 residuos. CSRA y CSRB actúan como canales de cationes activados por luz y desencadenan fotocorrientes despolarizantes. [68] [69] Se demostró que CSRA se localiza en la región del estigma utilizando análisis de inmunofluorescencia (Suzuki et al. 2003). La eliminación individual de la interferencia de ARN de CSRA y CSRB modificó las corrientes inducidas por la luz y reveló que CSRA media una corriente rápida y de alta saturación, mientras que CSRB una corriente lenta y de baja saturación. Ambas corrientes pueden desencadenar respuestas fotofóbicas y pueden tener un papel en la fototaxis, [70] [69] aunque la contribución exacta de los dos receptores aún no está clara. [57]
Como en todos los bikontes (plantas, cromalveoladas, excavadas, rizarias), las algas verdes tienen dos cilios, que no son idénticos. El cilio anterior siempre es más joven que el posterior . [71] [72] En cada ciclo celular, una célula hija recibe el cilio anterior y lo transforma en uno posterior. La otra hija hereda el cilio posterior maduro. Ambas hijas luego desarrollan un nuevo cilio anterior. [57]
Al igual que todos los demás nadadores ciliares, las algas verdes siempre nadan en espiral. La lateralidad de la espiral es robusta y está garantizada por la quiralidad de los cilios. Los dos cilios de las algas verdes tienen diferentes patrones de batimiento y funciones. En Chlamydomonas, la cascada de fototransducción altera el patrón de batimiento y la velocidad de batimiento de los dos cilios de manera diferencial en un patrón complejo. [44] [45] Esto da como resultado la reorientación de la trayectoria de natación helicoidal siempre que el eje de natación helicoidal no esté alineado con el vector de luz. [57]
La temperatura es un factor ambiental clave para los organismos vivos porque las tasas de reacción química y las características físicas de los materiales biológicos pueden cambiar sustancialmente con la temperatura. Los organismos vivos se aclimatan al estrés por frío y calor utilizando mecanismos adquiridos, incluida la capacidad de migrar a un entorno con temperaturas adecuadas para la habitabilidad. Una de las formas más simples del comportamiento para migrar a un entorno térmico adecuado es la termotaxis . La termotaxis se ha encontrado en organismos multicelulares, como Caenorhabditis elegans y Drosophila melanogaster , así como en organismos unicelulares, como Paramecium caudatum , Dictyostelium discoideum , Physarum polycephalum y Escherichia coli . [74] Las células individuales dentro de los organismos multicelulares también muestran termotaxis. Por ejemplo, los espermatozoides de los mamíferos migran a través del oviducto hasta el sitio de fertilización guiados por un aumento de la temperatura. [75] [76]
La investigación de cómo los organismos unicelulares migran hacia temperaturas preferidas comenzó hace más de 100 años. [74] En particular, el comportamiento termotáctico de las células de Paramecium ha sido bien estudiado. Las células de Paramecium se acumulan en sitios que están cerca de la temperatura de cultivo, es decir, la temperatura a la que se cultivan las células. [74] La acumulación en estos sitios ocurre porque las células con frecuencia invierten su dirección de nado cuando encuentran un cambio de temperatura que se desvía de la temperatura de cultivo y aumentan su velocidad de nado cuando experimentan un cambio de temperatura que se acerca a la temperatura de cultivo. [77] [78] La inversión en la dirección de nado es inducida por un potencial receptor despolarizante, que desencadena un potencial de acción en los cilios. [79] Estos estudios sobre células de Paramecium destacaron la termotaxis en organismos unicelulares hace más de 30 años, pero los mecanismos moleculares para la termorrecepción y la transducción de señales aún no se comprenden. [76]
La comprensión de los mecanismos moleculares de la termotaxis ha avanzado mucho en los últimos años, a partir de investigaciones sobre los espermatozoides de mamíferos . Los espermatozoides humanos migran hacia temperaturas más cálidas, que van desde los 29 °C a los 41 °C. [75] Los espermatozoides pueden detectar un gradiente de temperatura tan pequeño como 0,014 °C/mm, lo que sugiere que los espermatozoides detectan cambios temporales de temperatura en lugar de diferencias espaciales. [75] Se ha propuesto que varias moléculas sean moléculas sensoras, incluidas las opsinas y los canales de potencial receptor transitorio (TRP) como TRPV1 , TRPV4 y TRPM8 . [80] [81] [82] Los canales TRP son sensores multimodales para estímulos térmicos, químicos y mecánicos, pero la función de las opsinas como termosensores aún está por establecerse. [76]
La temperatura es un factor ambiental crítico también para las células de Chlamydomonas , que producen pequeñas proteínas de choque térmico , chaperoninas y proteínas de choque térmico HSP70 , y también experimentan otras respuestas de choque térmico para hacer frente al estrés térmico. [83] [84] [85] [86] En respuesta a un choque frío de 4 °C, las células detienen la proliferación y acumulan almidón y azúcar. [87] Se espera que las respuestas conductuales para evitar ambientes cálidos o fríos estresantes estén presentes en Chlamydomonas . Aunque se informa que las células de C. moewusii migran hacia temperaturas más cálidas en un gradiente de 10 °C a 15 °C, [88] no ha habido ningún informe en el que el rango de temperatura se haya manipulado sistemáticamente para examinar una relación con la temperatura de cultivo. Un estudio de 2019 demostró termotaxis en Chlamydomonas reinhardtii y encontró que entre 10 °C y 30 °C, las células de Chlamydomonas migraron hacia temperaturas más bajas independientemente de la temperatura de cultivo. [76]
A diferencia de la termotaxis, la fototaxis ha sido ampliamente estudiada en Chlamydomonas . Dos flagelos de Chlamydomonas se mueven en un patrón similar al de una brazada durante la natación hacia adelante y, durante la fototaxis, las células de Chlamydomonas realizan un giro hacia o desde una fuente de luz controlando el equilibrio de las fuerzas propulsoras generadas por los dos flagelos. [41] [89] El equilibrio depende de la concentración de iones de calcio intraflagelar; por lo tanto, la pérdida del control dependiente del calcio en mutantes ptx1 resulta en un defecto de fototaxis. [43] [90] [91] La dirección de la fototaxis en Chlamydomonas depende de la intensidad de la luz, pero también se ve afectada por las condiciones de reducción-oxidación (redox) intracelular. [92] Las células migran hacia una fuente de luz cuando la intensidad de la luz es débil, pero la dirección se invierte en condiciones reductoras. En contraste, las células nadan lejos de las fuentes de luz con fuerte intensidad, pero la dirección se invierte en condiciones oxidantes. [76]
Velocidades de natación
Velocidades de natación de flagelados y ciliados [2] Distribución lineal de datos de velocidad de natación
Curvas log-normales ajustadas para velocidades de natación de flagelados y ciliados
Las velocidades de natación se normalizaron dividiéndolas por la mediana de su distribución. Las distribuciones normalizadas muestran una similitud notable, tanto en las curvas ajustadas como en la incertidumbre de la estimación. [2]
En las algas flageladas , los cambios abruptos en la intensidad de la luz o los estímulos fóticos intensos inducen una rápida inversión flagelar y una natación hacia atrás transitoria. [94] [95] En las algas verdes , esta acción puede estar mediada por la fibra contráctil de la raíz que altera el ángulo entre los cuerpos basales . [96] Las células también pueden reaccionar rápidamente a estímulos mecánicos inesperados. Las contracciones de todo o nada en el ciliado pedunculado Vorticella pueden ocurrir a velocidades de 8 cm/s. [97] En algunas especies de heliozoos , los axópodos pueden retraerse completamente en 20 ms para atraer a la presa atrapada para la fagocitosis . [97] [98] [93]
Estas reacciones rápidas suelen ser inducidas por potenciales de acción (pulsos eléctricos unidireccionales que implican cambios rápidos y regenerativos en el potencial de membrana ). Si bien todas las células muestran cierta actividad eléctrica, la evidencia filogenética sugiere que la capacidad de propagar potenciales de acción puede haber sido un rasgo ancestral eucariota respaldado por el último ancestro común eucariota. Estos pueden haber surgido en respuesta a un daño accidental de la membrana y una entrada repentina de calcio. [99] La señalización bioeléctrica en forma de potenciales de acción ocurre órdenes de magnitud más rápido que cualquier otra modalidad de señalización, por ejemplo, difusión química , fosforilación de proteínas , etc. [93]
Para iniciar respuestas de escape rápidas , estas pueden haber sido acopladas directamente al aparato de motilidad, particularmente a estructuras flexibles y continuas de membrana como los cilios y los pseudópodos . La pérdida de canales de sodio / calcio dependientes del voltaje se correlaciona además con la pérdida de cilios en muchos taxones. En los protistas, los potenciales de acción de todo o nada ocurren casi exclusivamente en asociación con las membranas ciliares, [100] [101] [102] con la excepción de algunas diatomeas no ciliadas . [103] [104] Los potenciales graduados ocurren en las amebas , también para el control del movimiento. [105] [93]
En Chlamydomonas , las corrientes flagelares similares a potenciales de acción inducen respuestas fotofóbicas e inversión de los flagelos (a través del canal de calcio dependiente de voltaje Cav2), mientras que las corrientes de los fotorreceptores provocan respuestas mucho más suaves. [106] Aquí, un canal mecanosensorial de la familia del potencial receptor transitorio se localiza en la base ciliar, mientras que Cav2 se localiza solo en las regiones distales de los cilios. [107] [108] En Paramecium , las hiperpolarizaciones aumentan la frecuencia del batido ciliar, mientras que las despolarizaciones tienen el efecto opuesto y eventualmente conducen a una inversión ciliar. Las despolarizaciones por encima de un cierto umbral dan como resultado potenciales de acción, debido a la apertura de los canales Cav ubicados exclusivamente en la membrana ciliar. [109] [110] Los canales de potasio , que también residen en la membrana, ayudan a restaurar el potencial de membrana en reposo . [93]
Los eucariotas manipulan su potencial de membrana para lograr transiciones entre diferentes comportamientos. Se han registrado secuencias bioeléctricas complejas en asociación con comportamientos integrados de alimentación y depredación en Favella . [111] Los comportamientos repetitivos surgen de picos rítmicos. En los ciliados, las despolarizaciones rítmicas controlan el caminar rápido y lento mediante cilios compuestos similares a tentáculos llamados cirros , [112] lo que permite escapar de callejones sin salida [113] y rituales de cortejo en gametos conjugados . [114] [115] En Stentor , los potenciales de acción producen contracciones de todo el cuerpo. [116] Finalmente, los sistemas excitables que operan cerca de bifurcaciones pueden admitir ciclos límite, que se manifiestan como picos eléctricos repetitivos o rítmicos y comportamientos repetitivos. En última instancia, esto puede conducir a la habituación. [117] [118] [93]
Arriba: Esquemas de los pasos de producción para el biohíbrido C. reinhardtii . Abajo: Imágenes SEM de microalgas desnudas (izquierda) y microalgas biohíbridas (derecha) recubiertas con nanopartículas de óxido de hierro recubiertas de quitosano (CSIONPs). Las imágenes fueron pseudocoloreadas. Un color verde más oscuro en la imagen SEM de la derecha representa el recubrimiento de quitosano en la pared celular de la microalga. Las partículas de color naranja representan CSIONPs.
Los micronadadores biohíbridos pueden definirse como micronadadores que consisten en componentes tanto biológicos como artificiales, por ejemplo, uno o varios microorganismos vivos unidos a una o varias partes sintéticas. [120] [121] En 1999, Montemagno y Bachand publicaron un artículo que identificaba estrategias de unión específicas de moléculas biológicas a sustratos nanofabricados, lo que permitió la preparación de sistemas nanoelectromecánicos inorgánicos/orgánicos híbridos (NEMS). [122] Describieron la producción de grandes cantidades de F1-ATPasa a partir de la bacteria termófila Bacillus PS3 para la preparación de motores biomoleculares F1-ATPasa inmovilizados en un patrón de nanoarreglo de oro, cobre o níquel producido por litografía de haz de electrones . Estas proteínas se unieron a microesferas de un micrón etiquetadas con un péptido sintético . En consecuencia, lograron la preparación de una plataforma con sitios químicamente activos y el desarrollo de dispositivos biohíbridos capaces de convertir la energía de los motores biomoleculares en trabajo útil. [121]
Durante la última década, los microrobots biohíbridos, en los que microorganismos móviles vivos están físicamente integrados con estructuras artificiales no atadas, han ganado un creciente interés para permitir la locomoción activa y la entrega de carga a un destino objetivo. [123] [ 124] [125] [126] Además de la motilidad, las capacidades intrínsecas de detectar y provocar una respuesta apropiada a los cambios artificiales y ambientales hacen que los microrobots biohíbridos basados en células sean atractivos para el transporte de carga a las cavidades inaccesibles del cuerpo humano para la entrega activa local de agentes de diagnóstico y terapéuticos. [127] [128] [129] La locomoción activa, la orientación y la dirección de agentes terapéuticos y de diagnóstico concentrados integrados en microrobots móviles al sitio de acción pueden superar los desafíos existentes de las terapias convencionales. [130] [131] [132] Para este fin, se han utilizado comúnmente bacterias con perlas adheridas y cuerpos de células fantasma. [133] [134] [135] [136] [137] [138] [139] [140] [119]
Chlamydomonas reinhardtii es una microalga verde unicelular. La C. reinhardtii de tipo salvajetiene una forma esférica que promedia alrededor de 10 μm de diámetro. [141] Este microorganismo puede percibir la luz visible y ser dirigido por ella (es decir, fototaxis ) con altas velocidades de nado en el rango de 100-200 μm/s. [129] Tiene autofluorescencia natural que permite imágenes fluorescentes sin etiquetas. [141] C. reinhardtii ha sido explorado activamente como el componente vivo de microrobots biohíbridos para la administración activa de terapias. [129] Son biocompatibles con células de mamíferos sanos, no dejan toxinas conocidas, son móviles en los medios fisiológicamente relevantes y permiten la modificación de la superficie para transportar carga en la pared celular. [129] [142] [143] [144] [145] Se han propuestoestrategias de unión alternativas para C. reinhardtii para el ensamblaje mediante la modificación de las superficies interactuantes mediante interacciones electrostáticas [129] [142] y enlaces covalentes. [146] [119]
^ Hinchliff, Cody E.; Smith, Stephen A.; Allman, James F.; Burleigh, J. Gordon; Chaudhary, Ruchi; Coghill, Lyndon M.; Crandall, Keith A.; Deng, Jiabin; Drew, Bryan T.; Gazis, Romina; Gude, Karl; Hibbett, David S.; Katz, Laura A.; Laughinghouse, H. Dail; McTavish, Emily Jane; Midford, Peter E.; Owen, Christopher L.; Ree, Richard H.; Rees, Jonathan A.; Soltis, Douglas E.; Williams, Tiffani; Cranston, Karen A. (2015). "Síntesis de filogenia y taxonomía en un árbol de la vida completo". Actas de la Academia Nacional de Ciencias . 112 (41): 12764–12769. Código Bibliográfico :2015PNAS..11212764H. doi : 10.1073/pnas.1423041112 . PMC 4611642 . PMID 26385966.
^ abcdefghi Lisicki, Maciej; Velho Rodrigues, Marcos F.; Goldstein, Raymond E.; Lauga, Eric (2019). "Los microorganismos eucariotas nadadores exhiben una distribución de velocidad universal". eLife . 8 . arXiv : 1907.00906 . doi : 10.7554/eLife.44907 . PMC 6634970 . PMID 31310238.El material fue copiado de esta fuente, que está disponible bajo una Licencia Creative Commons Atribución 4.0 Internacional.
^ Wichterman, R. (6 de diciembre de 2012). La biología del paramecio. Saltador. ISBN9781475703726.
^ Buetow, Dennis E. (2011). "Euglena". eLS . doi : 10.1002/9780470015902.a0001964.pub3. ISBN978-0470016176.
^ Harris, Elizabeth H. (7 de marzo de 2009). The Chlamydomonas Sourcebook: Introduction to Chlamydomonas and Its Laboratory Use: Volume 1 (Libro de consulta sobre Chlamydomonas: Introducción a Chlamydomonas y su uso en laboratorio: Volumen 1). Academic Press. ISBN97-80080919553.
^ Adam, Rodney D. (2001). "Biología de Giardia lamblia". Clinical Microbiology Reviews . 14 (3): 447–475. doi :10.1128/CMR.14.3.447-475.2001. PMC 88984 . PMID 11432808.
^ Bray, Dennis (2 de noviembre de 2000). Movimientos celulares: de las moléculas a la motilidad. Garland Science. ISBN9781136844355.
^ Cappuccinelli, P. (1980). "El movimiento de las células eucariotas". Motilidad de las células vivas . págs. 59–74. doi :10.1007/978-94-009-5812-8_4. ISBN978-0-412-15770-7.
^ Sleigh, MA (1974) Cilia y flagelos . Prensa académica.
^ Nicastro, D.; McIntosh, JR; Baumeister, W. (2005). "Estructura 3D de flagelos eucariotas en estado quiescente revelada por tomografía crioelectrónica". Actas de la Academia Nacional de Ciencias . 102 (44): 15889–15894. Bibcode :2005PNAS..10215889N. doi : 10.1073/pnas.0508274102 . PMC 1276108 . PMID 16246999.
^ Moran, Jonathan; McKean, Paul G.; Ginger, Michael L. (2014). "Flagelos eucariotas: variaciones en forma, función y composición durante la evolución". BioScience . 64 (12): 1103–1114. doi : 10.1093/biosci/biu175 . ISSN 1525-3244.
^ Dawson, Scott C; Paredez, Alexander R (2013). "Paisajes alternativos del citoesqueleto: novedad y evolución del citoesqueleto en protistos excavados basales". Current Opinion in Cell Biology . 25 (1): 134–141. doi :10.1016/j.ceb.2012.11.005. PMC 4927265 . PMID 23312067.
^ ab Atkinson, A.; Polimene, L.; Fileman, ES; Widdicombe, CE; McEvoy, AJ; Smyth, TJ; Djeghri, N.; Sailley, SF; Cornwell, LE (2018). ""Comentario. ¿Qué impulsa la estacionalidad del plancton en un mar de plataforma estratificado? Algunas teorías en competencia y complementarias"]" (PDF) . Limnología y Oceanografía . 63 (6): 2877–2884. Bibcode :2018LimOc..63.2877A. doi :10.1002/lno.11036. S2CID 91380765.
^ Singleton, Paul (2006). Diccionario de microbiología y biología molecular, 3.ª edición, revisada . Chichester, Reino Unido: John Wiley & Sons. pp. 32. ISBN978-0-470-03545-0.
^ David J. Patterson. "Amebas: protistas que se mueven y se alimentan utilizando seudópodos". Proyecto web El árbol de la vida.
^ "Las amebas". Universidad de Edimburgo. Archivado desde el original el 10 de junio de 2009.
^ Diccionario de biología, 2004, consultado el 1 de enero de 2011.
^ Patterson, David J. (2000) "Flagelados: Protistas heterotróficos con flagelos" Árbol de la vida .
^ Jahn, TL; Votta, JJ (1972). "Locomoción de los protozoos". Revisión anual de mecánica de fluidos . 4 : 93–116. Código Bibliográfico :1972AnRFM...4...93J. doi :10.1146/annurev.fl.04.010172.000521.
^ ab Brennen, C.; Winet, H. (1977). "Mecánica de fluidos de la propulsión por cilios y flagelos". Revista anual de mecánica de fluidos . 9 : 339–398. Código Bibliográfico :1977AnRFM...9..339B. doi :10.1146/annurev.fl.09.010177.002011.
^ Allen, NS; Allen, RD (1978). "Transmisión citoplasmática en plantas verdes". Revista anual de biofísica y bioingeniería . 7 : 497–526. doi :10.1146/annurev.bb.07.060178.002433. PMID 352247.
^ Lyons, RA; Saridogan, E.; Djahanbakhch, O. (2006). "La importancia reproductiva de los cilios de las trompas de Falopio humanas". Human Reproduction Update . 12 (4): 363–372. doi : 10.1093/humupd/dml012 . PMID 16565155.
^ Lauga, Eric; Thomas R Powers (25 de agosto de 2009). "La hidrodinámica de los microorganismos nadadores". Informes sobre el progreso en física . 72 (9): 096601. arXiv : 0812.2887 . Bibcode :2009RPPh...72i6601L. doi :10.1088/0034-4885/72/9/096601. S2CID 3932471.
^ ab Rode, Sebastian; Elgeti, Jens; Gompper, Gerhard (2021). "Micronadadores multiciliados: coordinación metacronal y natación helicoidal". The European Physical Journal E . 44 (6): 76. arXiv : 2103.11447 . doi :10.1140/epje/s10189-021-00078-x. PMC 8187229 . PMID 34101070.
^ Fritz-Laylin, Lillian K. (2020). "La evolución de la motilidad celular animal". Current Biology . 30 (10): R477–R482. Código Bibliográfico :2020CBio...30.R477F. doi : 10.1016/j.cub.2020.03.026 . PMID 32428485. S2CID 218711237.
^ abcdef De Maleprade, Hélène; Moisy, Frédéric; Ishikawa, Takuji; Goldstein, Raymond E. (2020). "Motilidad y fototaxis de Gonium, el alga colonial diferenciada más simple". Physical Review E . 101 (2): 022416. arXiv : 1911.08837 . Bibcode :2020PhRvE.101b2416D. doi :10.1103/PhysRevE.101.022416. PMID 32168596. S2CID 211858528.El material fue copiado de esta fuente, que está disponible bajo una Licencia Creative Commons Atribución 4.0 Internacional.
^ ab Coleman, AW (2012). "Un análisis comparativo de Volvocaceae (Chlorophyta)1". Revista de fisiología . 48 (3): 491–513. Código Bibliográfico :2012JPcgy..48..491C. doi :10.1111/j.1529-8817.2012.01168.x. PMID 27011065. S2CID 422091.
^ Pennak, Robert W (1978). Invertebrados de agua dulce de los Estados Unidos (segunda edición). John Wiley & Sons . pp. 43. ISBN0-471-04249-8.
^ Weismann, August (1889). "Ensayos sobre la herencia y problemas biológicos afines: por el Dr. August Weismann... Ed. por Edward B. Poulton... Selmar Schönland... y Arthur E. Shipley... Traducción autorizada".
^ Huxley, Julian S. (22 de marzo de 2012). El individuo en el reino animal. Cambridge University Press. ISBN9781107606074.
^ Kirk, David L. (8 de septiembre de 2005). Volvox: una búsqueda de los orígenes moleculares y genéticos de la multicelularidad y la diferenciación celular. Cambridge University Press. ISBN9780521019149.
^ Goldstein, Raymond E. (2015). "Las algas verdes como organismos modelo para la dinámica de fluidos biológicos". Revista anual de mecánica de fluidos . 47 : 343–375. doi :10.1146/annurev-fluid-010313-141426. PMC 4650200. PMID 26594068 .
^ Goldstein, Raymond E. (2016). "Conferencia del Premio Bachelor Dinámica de fluidos a escala celular". Journal of Fluid Mechanics . 807 : 1–39. Bibcode :2016JFM...807....1G. doi : 10.1017/jfm.2016.586 . S2CID 55745525.
^ abc Hoops, HJ (1997). "Motilidad en los volvocales coloniales y multicelulares: estructura, función y evolución". Protoplasma . 199 (3–4): 99–112. doi :10.1007/BF01294499. S2CID 22315728.
^ ab Drescher, K.; Goldstein, RE; Tuval, I. (2010). "Fidelidad de la fototaxis adaptativa". Actas de la Academia Nacional de Ciencias . 107 (25): 11171–11176. doi : 10.1073/pnas.1000901107 . PMC 2895142 . PMID 20534560.
^ Drescher, Knut; Goldstein, Raymond E.; Michel, Nicolas; Polin, Marco; Tuval, Idan (2010). "Medición directa del campo de flujo alrededor de microorganismos nadadores". Physical Review Letters . 105 (16): 168101. arXiv : 1008.2681 . Código Bibliográfico :2010PhRvL.105p8101D. doi :10.1103/PhysRevLett.105.168101. PMID 21231017. S2CID 8306079.
^ Guasto, Jeffrey S.; Johnson, Karl A.; Gollub, JP (2010). "Flujos oscilatorios inducidos por microorganismos que nadan en dos dimensiones". Physical Review Letters . 105 (16): 168102. arXiv : 1008.2535 . Bibcode :2010PhRvL.105p8102G. doi :10.1103/PhysRevLett.105.168102. PMID 21231018. S2CID 9533722.
^ Bennett, Rachel R.; Golestanian, Ramin (2015). "Un mecanismo de dirección para la fototaxis en Chlamydomonas". Journal of the Royal Society Interface . 12 (104). doi :10.1098/rsif.2014.1164. PMC 4345482 . PMID 25589576.
^ Arrieta, Jorge; Barreira, Ana; Chioccioli, Mauricio; Polin, Marco; Tuval, Idan (2017). "Fototaxis más allá del giro: acumulación persistente y aclimatación de respuesta de la microalga Chlamydomonas reinhardtii". Informes científicos . 7 (1): 3447. arXiv : 1611.08224 . Código Bib : 2017NatSR...7.3447A. doi :10.1038/s41598-017-03618-8. PMC 5471259 . PMID 28615673.
^ Tsang, Alan CH; Lam, Amy T.; Riedel-Kruse, Ingmar H. (2018). "Movimiento poligonal y fototaxis adaptable a través del cambio de ritmo flagelar en la micronadadora Euglena gracilis". Nature Physics . 14 (12): 1216–1222. Código Bibliográfico :2018NatPh..14.1216T. doi :10.1038/s41567-018-0277-7. S2CID 126294173.
^ ab Foster, KW; Smyth, RD (1980). "Antenas de luz en algas fototácticas". Microbiological Reviews . 44 (4): 572–630. doi :10.1128/mr.44.4.572-630.1980. PMC 373196 . PMID 7010112.
^ Hegemann, Peter (2008). "Fotorreceptores sensoriales de algas". Revisión anual de biología vegetal . 59 : 167–189. doi :10.1146/annurev.arplant.59.032607.092847. PMID 18444900.
^ ab Kamiya, R.; Witman, GB (1984). "Los niveles submicromolares de calcio controlan el equilibrio de los latidos entre los dos flagelos en modelos desmembranados de Chlamydomonas". Journal of Cell Biology . 98 (1): 97–107. doi :10.1083/jcb.98.1.97. PMC 2112995 . PMID 6707098.
^ ab Josef, Keith; Saranak, Jureepan; Foster, Kenneth W. (2005). "Comportamiento ciliar de una Chlamydomonas reinhardtii fototáctica negativa". Motilidad celular y citoesqueleto . 61 (2): 97–111. doi :10.1002/cm.20069. PMID 15849714.
^ ab Josef, Keith; Saranak, Jureepan; Foster, Kenneth W. (2006). "Análisis de sistemas lineales del comportamiento de dirección ciliar asociado con la fototaxis negativa en Chlamydomonas reinhardtii". Motilidad celular y citoesqueleto . 63 (12): 758–777. doi :10.1002/cm.20158. PMID 16986140.
^ Yoshimura, Kenjiro; Kamiya, Ritsu (2001). "La sensibilidad del fotorreceptor de Chlamydomonas está optimizada para la frecuencia de rotación del cuerpo celular". Fisiología vegetal y celular . 42 (6): 665–672. doi : 10.1093/pcp/pce084 . PMID 11427687.
^ Leptos, Kyriacos C.; Chioccioli, Maurizio; Furlan, Silvano; Pesci, Adriana I .; Goldstein, Raymond E. (2018). "Una fotorrespuesta flagelar adaptativa determina la dinámica de la dirección fototáctica precisa en Chlamydomonas ". doi :10.1101/254714. S2CID 90374721.{{cite journal}}: Requiere citar revista |journal=( ayuda )
^ Kirk, David L. (2004). "Volvox". Current Biology . 14 (15): R599–R600. Código Bibliográfico :2004CBio...14.R599K. doi : 10.1016/j.cub.2004.07.034 . PMID 15296767. S2CID 235312006.
^ Arakaki, Yoko; Kawai-Toyooka, Hiroko; Hamamura, Yuki; Higashiyama, Tetsuya; Noga, Akira; Hirono, Masafumi; Olson, Bradley JSC; Nozaki, Hisayoshi (2013). "Revelado el organismo multicelular integrado más simple". MÁS UNO . 8 (12): e81641. Código Bib : 2013PLoSO...881641A. doi : 10.1371/journal.pone.0081641 . PMC 3859500 . PMID 24349103.
^ Herron, Matthew D.; Michod, Richard E. (2008). "Evolución de la complejidad en las algas volvocinas: transiciones en la individualidad a través de la mirada de Darwin". Evolution . 62 (2): 436–451. doi :10.1111/j.1558-5646.2007.00304.x. PMID 18031303. S2CID 12139760.
^ Nozaki, Hisayoshi (1990). "Ultraestructura de la matriz extracelular de Gonium (Volvocales, Chlorophyta)". Phycologia . 29 (1): 1–8. Bibcode :1990Phyco..29....1N. doi :10.2216/i0031-8884-29-1-1.1.
^ abc Moore, AR (1916). "El mecanismo de orientación en Gonium". Revista de zoología experimental . 21 (3): 431–432. Código Bibliográfico :1916JEZ....21..431M. doi :10.1002/jez.1400210306.
^ abc Mast, SO (1916). "El proceso de orientación en el organismo colonial, Gonium pectorale, y un estudio de la estructura y función de la mancha ocular". Revista de zoología experimental . 20 (1): 1–17. Bibcode :1916JEZ....20....1M. doi :10.1002/jez.1400200102.
^ ab Greuel, Brian T.; Floyd, Gary L. (1985). "Desarrollo del aparato flagelar y orientación flagelar en el alga verde colonial Gonium pectorale (Volvocales)1". Revista de fisiología . 21 (3): 358–371. Código Bibliográfico :1985JPcgy..21..358G. doi :10.1111/j.0022-3646.1985.00358.x. S2CID 85760904.
^ Harper, RA (1912). "La estructura y el desarrollo de la colonia en Gonium". Transactions of the American Microscopical Society . 31 (2): 65–83. doi :10.2307/3221328. JSTOR 3221328.
^ Müller OF (1782) Kleine Schriften Aus Der Naturhistorie , Dessau, herausgegeben von JAE Goeze, págs.
^ abcdefghij Jékely, Gáspár (2009). "Evolución de la fototaxis". Philosophical Transactions of the Royal Society B: Biological Sciences . 364 (1531): 2795–2808. doi :10.1098/rstb.2009.0072. PMC 2781859 . PMID 19720645.El material fue copiado de esta fuente, que está disponible bajo una Licencia Creative Commons Atribución 4.0 Internacional.
^ Clark, MA, Choi, J. y Douglas, M. (2018) Características de los protistas Biología 2e. OpenStax . ISBN 9781947172951 . El material fue copiado de esta fuente, que está disponible bajo una Licencia Creative Commons Atribución 4.0 Internacional
^ Häder, D. -P; Lebert, M. (19 de junio de 2001). Fotomovimiento. Elsevier. ISBN9780080538860.
^ Grung, Merete; Kreimer, Georg; Calenberg, Michael; Melkonian, Michael; Liaaen-Jensen, Synnøve (1994). "Carotenoides en el aparato ocular del alga verde flagelada Spermatozopsis similis: Adaptación al fotorreceptor basado en la retina". Planta . 193 . doi :10.1007/BF00191604. S2CID 29443649.
^ Renninger, S.; Backendorf, E.; Kreimer, G. (2001). "Subfraccionamiento de los aparatos de la mancha ocular del alga verde Spermatozopsis similis: aislamiento y caracterización de los glóbulos de la mancha ocular". Planta . 213 (1): 51–63. Bibcode :2001Plant.213...51R. doi :10.1007/s004250000473. PMID 11523656. S2CID 24880210.
^ Arnott, Howard J.; Brown, R. Malcolm (1967). "Ultraestructura de la mancha ocular y su posible significado en la fototaxis de Tetracystis excentrica*†". Revista de protozoología . 14 (4): 529–539. doi :10.1111/j.1550-7408.1967.tb02038.x.
^ Melkonian, M.; Robenek, H. (1979). "La mancha ocular del flagelado Tetraselmis cordiformis stein (Chlorophyceae): especialización estructural de la membrana externa del cloroplasto y su posible significado en la fototaxis de las algas verdes". Protoplasma . 100 (2): 183–197. doi :10.1007/BF01283929. S2CID 24606055.
^ Melkonian, Michael (1978). "Estructura y significado de los sistemas de raíces flagelares cruciformes en algas verdes: Investigaciones comparativas en especies de Chlorosarcinopsis (Chlorosarcinales)". Plant Systematics and Evolution . 130 (3–4): 265–292. Bibcode :1978PSyEv.130..265M. doi :10.1007/BF00982810. S2CID 22938771.
^ Fomentar, Kenneth W.; Saranak, Jureepan; Patel, Nayana; Zarilli, Gerald; Okabe, Masami; Kline, Toni; Nakanishi, Koji (1984). "Una rodopsina es el fotorreceptor funcional para la fototaxis en el eucariota unicelular Chlamydomonas". Naturaleza . 311 (5988): 756–759. Código Bib :1984Natur.311..756F. doi :10.1038/311756a0. PMID 6493336. S2CID 4263301.
^ Nagel, G.; Ollig, D.; Fuhrmann, M.; Kateriya, S.; Musti, AM; Bamberg, E.; Hegemann, P. (2002). "Canal rodopsina-1: un canal de protones activado por luz en algas verdes". Ciencia . 296 (5577): 2395–2398. Código bibliográfico : 2002 Ciencia... 296.2395N. doi : 10.1126/ciencia.1072068. PMID 12089443. S2CID 206506942.
^ Nagel, G.; Szellas, T.; Huhn, W.; Kateriya, S.; Adeishvili, N.; Berthold, P.; Ollig, D.; Hegemann, P.; Bamberg, E. (2003). "Channelrhodopsin-2, un canal de membrana selectivo de cationes activado directamente por luz". Actas de la Academia Nacional de Ciencias . 100 (24): 13940–13945. Bibcode :2003PNAS..10013940N. doi : 10.1073/pnas.1936192100 . PMC 283525 . PMID 14615590.
^ ab Sineshchekov, OA; Jung, K.-H.; Spudich, JL (2002). "Dos rodopsinas median la fototaxis a luz de baja y alta intensidad en Chlamydomonas reinhardtii". Actas de la Academia Nacional de Ciencias . 99 (13): 8689–8694. doi : 10.1073/pnas.122243399 . PMC 124360 . PMID 12060707.
^ ab Berthold, Peter; Tsunoda, Satoshi P.; Ernst, Oliver P.; Mages, Wolfgang; Gradmann, Dietrich; Hegemann, Peter (2008). "La canalrodopsina-1 inicia la fototaxis y las respuestas fotofóbicas en Chlamydomonas mediante despolarización inmediata inducida por luz". La célula vegetal . 20 (6): 1665–1677. doi :10.1105/tpc.108.057919. PMC 2483371 . PMID 18552201.
^ Govorunova, Elena G.; Jung, Kwang-Hwan; Sineshchekov, Oleg A.; Spudich, John L. (2004). "Rodopsinas sensoriales a y B de Chlamydomonas: contenido celular y función en las respuestas fotofóbicas". Revista biofísica . 86 (4): 2342–2349. Código Bibliográfico :2004BpJ....86.2342G. doi :10.1016/S0006-3495(04)74291-5. PMC 1304083 . PMID 15041672.
^ Cavalier-Smith, T. (2002). "El origen fagotrófico de los eucariotas y la clasificación filogenética de los protozoos". Revista internacional de microbiología sistemática y evolutiva . 52 (2): 297–354. doi :10.1099/00207713-52-2-297. PMID 11931142.
^ Cavalier-Smith, Thomas (2009). "Megafilogenia, planes corporales celulares, zonas adaptativas: causas y cronología de las radiaciones basales eucariotas". Journal of Eukaryotic Microbiology . 56 (1): 26–33. doi : 10.1111/j.1550-7408.2008.00373.x . PMID 19340985. S2CID 10205240.
^ Harris, Elizabeth H. (2009) "El género Chlamydomonas" en The Chlamydomonas Sourcebook (segunda edición), capítulo 1, volumen 1, páginas 1-24. ISBN 9780080919553 doi :10.1016/B978-0-12-370873-1.00001-0
^ abc Jennings HS (1907) "Comportamiento de los organismos inferiores" The American Naturalist , 41 (481): 42-44.
^ abc Bahat, Anat; Tur-Kaspa, Ilan; Gakamsky, Anna; Giojalas, Laura C.; Breitbart, Haim; Eisenbach, Michael (2003). "Termotaxis de los espermatozoides de los mamíferos: un mecanismo de navegación potencial en el tracto genital femenino". Nature Medicine . 9 (2): 149–150. doi :10.1038/nm0203-149. hdl : 11336/66658 . PMID 12563318. S2CID 36538049.
^ abcde Sekiguchi, Masaya; Kameda, Shigetoshi; Kurosawa, Satoshi; Yoshida, Megumi; Yoshimura, Kenjiro (2018). "La termotaxis en Chlamydomonas se produce mediante la excitación de la membrana y está controlada por condiciones redox". Informes científicos . 8 (1): 16114. Código bibliográfico : 2018NatSR...816114S. doi :10.1038/s41598-018-34487-4. PMC 6208428 . PMID 30382191.El material fue copiado de esta fuente, que está disponible bajo una Licencia Creative Commons Atribución 4.0 Internacional.
^ Tawada, K.; Oosawa, F. (1972). "Respuestas de los paramecios a los cambios de temperatura". Revista de protozoología . 19 (1): 53–57. doi :10.1111/j.1550-7408.1972.tb03412.x. PMID 5008849.
^ Nakaoka, Yasuo; Oosawa, Fumio (1977). "Comportamiento sensible a la temperatura de Paramecium caudatum". La Revista de Protozoología . 24 (4): 575–580. doi :10.1111/j.1550-7408.1977.tb01018.x.
^ Hennessey, Todd M.; Saimi, Yoshiro; Kung, Ching (1983). "Una despolarización inducida por calor de Paramecium y su relación con el comportamiento de evitación térmica". Journal of Comparative Physiology A . 153 : 39–46. doi :10.1007/BF00610340. S2CID 7152549.
^ Pérez-Cerezales, Serafín; Boryshpolets, Sergii; Afanzar, Oshri; Brandis, Alexander; Nevo, Reinat; Kiss, Vladimir; Eisenbach, Michael (2015). "Participación de las opsinas en la termotaxis de los espermatozoides de mamíferos". Scientific Reports . 5 : 16146. Bibcode :2015NatSR...516146P. doi :10.1038/srep16146. PMC 4633616 . PMID 26537127.
^ Hamano, Koh-Ichi; Kawanishi, Tae; Mizuno, Atsuko; Suzuki, Makoto; Takagi, Yuji (2016). "Participación del receptor de potencial transitorio vanilloide (TRPV) 4 en la termotaxis del esperma de ratón". Revista de reproducción y desarrollo . 62 (4): 415–422. doi :10.1262/jrd.2015-106. PMC 5004798 . PMID 27180924.
^ De Blas, Gerardo A.; Darszón, Alberto; Ocampo, Ana Y.; Serrano, Carmen J.; Castellano, Laura E.; Hernández-González, Enrique O.; Chirinos, Mayel; Larrea, Fernando; Beltrán, Carmen; Treviño, Claudia L. (2009). "TRPM8, un canal versátil en el esperma humano". MÁS UNO . 4 (6): e6095. Código Bib : 2009PLoSO...4.6095D. doi : 10.1371/journal.pone.0006095 . PMC 2705237 . PMID 19582168.
^ Kloppstech, Klaus; Meyer, Gabriele; Schuster, Gadi; Ohad, Itzhak (1985). "Síntesis, transporte y localización de una proteína de choque térmico de 22 kd con código nuclear en las membranas de los cloroplastos de guisantes y Chlamydomonas reinhardi". The EMBO Journal . 4 (8): 1901–1909. doi :10.1002/j.1460-2075.1985.tb03869.x. PMC 554439 . PMID 16453628.
^ Van Lis, Robert; Atteia, Ariane; Mendoza-Hernández, Guillermo; GonzáLez-Halphen, Diego (2003). "Identificación de nuevos componentes proteicos mitocondriales de Chlamydomonas reinhardtii. Un enfoque proteómico". Fisiología vegetal . 132 (1): 318–330. doi :10.1104/pp.102.018325. PMC 166977 . PMID 12746537.
^ von Gromoff, ED; Treier, U.; Beck, CF (1989). "Tres genes de choque térmico inducibles por luz de Chlamydomonas reinhardtii". Biología molecular y celular . 9 (9): 3911–3918. doi :10.1128/mcb.9.9.3911-3918.1989. PMC 362453 . PMID 2779571.
^ Schroda, Michael; Hemme, Dorothea; Mühlhaus, Timo (2015). "La respuesta de Chlamydomonas al estrés térmico". The Plant Journal . 82 (3): 466–480. doi : 10.1111/tpj.12816 . PMID 25754362.
^ Valledor, Luis; Furuhashi, Takeshi; Hanak, Anne-Mette; Weckwerth, Wolfram (2013). "Adaptación sistémica al estrés por frío de Chlamydomonas reinhardtii". Molecular & Cellular Proteomics . 12 (8): 2032–2047. doi : 10.1074/mcp.M112.026765 . PMC 3734567 . PMID 23564937.
^ Clegg, Mark R.; Maberly, Stephen C.; Jones, Roger I. (2003). "Respuestas conductuales de los flagelados fitoplanctónicos de agua dulce a un gradiente de temperatura". Revista Europea de Ficología . 38 (3): 195–203. Bibcode :2003EJPhy..38..195C. doi : 10.1080/0967026031000121697 . S2CID 85353895.
^ Isogai, Nahoko; Kamiya, Ritsu; Yoshimura, Kenjiro (2000). "Dominancia entre los dos flagelos durante el giro fototáctico en Chlamydomonas". Zoological Science . 17 (9): 1261–1266. doi : 10.2108/zsj.17.1261 . S2CID 84890095.
^ Horst, CJ; Witman, GB (1993). "Ptx1, un mutante no fototáctico de Chlamydomonas, carece de control de la dominancia flagelar". Journal of Cell Biology . 120 (3): 733–741. doi :10.1083/jcb.120.3.733. PMC 2119553 . PMID 8425899.
^ Okita, Noriko; Isogai, Nahoko; Hirono, Masafumi; Kamiya, Ritsu; Yoshimura, Kenjiro (2005). "Actividad fototáctica en mutantes no fototácticos de Chlamydomonas deficientes en el control de la dominancia flagelar dependiente de Ca2 + o en la dineína del brazo interno". Revista de ciencia celular . 118 (3): 529–537. doi : 10.1242/jcs.01633 . PMID 15657081. S2CID 2379702.
^ Wakabayashi, K.-i.; Misawa, Y.; Mochiji, S.; Kamiya, R. (2011). "El equilibrio reducción-oxidación regula el signo de la fototaxis en Chlamydomonas reinhardtii". Actas de la Academia Nacional de Ciencias . 108 (27): 11280–11284. Bibcode :2011PNAS..10811280W. doi : 10.1073/pnas.1100592108 . PMC 3131381 . PMID 21690384.
^ abcdef Wan, Kirsty Y.; Jékely, Gáspár (2021). "Orígenes de la excitabilidad eucariota". Philosophical Transactions of the Royal Society B: Biological Sciences . 376 (1820). arXiv : 2007.13388 . doi :10.1098/rstb.2019.0758. PMC 7935092 . PMID 33487111.El material fue copiado de esta fuente, que está disponible bajo una Licencia Creative Commons Atribución 4.0 Internacional.
^ Foster, KW; Smyth, RD (1980). "Antenas de luz en algas fototácticas". Microbiological Reviews . 44 (4): 572–630. doi :10.1128/MR.44.4.572-630.1980. PMC 373196 . PMID 7010112.
^ Holland, EM; Harz, H.; Uhl, R.; Hegemann, P. (1997). "Control de las respuestas conductuales fóbicas mediante fotocorrientes inducidas por rodopsina en Chlamydomonas". Biophysical Journal . 73 (3): 1395–1401. Bibcode :1997BpJ....73.1395H. doi :10.1016/S0006-3495(97)78171-2. PMC 1181038 . PMID 9284306.
^ Hayashi, Masahito; Yagi, Toshiki; Yoshimura, Kenjiro; Kamiya, Ritsu (1998). "Observación en tiempo real de la reorientación del cuerpo basal inducida por Ca2+ en Chlamydomonas ". Motilidad celular y citoesqueleto . 41 (1): 49–56. doi :10.1002/(SICI)1097-0169(1998)41:1<49::AID-CM4>3.0.CO;2-A. PMID 9744298.
^ ab Moriyama, Yasushige; Hiyama, Shigeo; Asai, Hiroshi (1998). "Demostración cinematográfica en video de alta velocidad de la contracción del tallo y del zooide de Vorticella convallaria". Revista biofísica . 74 (1): 487–491. Bibcode :1998BpJ....74..487M. doi :10.1016/s0006-3495(98)77806-3. PMC 1299401 . PMID 9449349.
^ Ando, Motonori; Shigenaka, Yoshinobu (1989). "Estructura y función del citoesqueleto en heliozoos: I. Mecanismo de contracción axopodial rápida en Echinosphaerium ". Motilidad celular y citoesqueleto . 14 (2): 288–301. doi :10.1002/cm.970140214.
^ Brunet, Thibaut; Arendt, Detlev (2016). "De la respuesta al daño a los potenciales de acción: evolución temprana de los módulos neurales y contráctiles en eucariotas madre". Philosophical Transactions of the Royal Society B: Biological Sciences . 371 (1685). doi :10.1098/rstb.2015.0043. PMC 4685582 . PMID 26598726.
^ Eckert, R.; Brehm, P. (1979). "Mecanismos iónicos de excitación en paramecio". Revista anual de biofísica y bioingeniería . 8 : 353–383. doi :10.1146/annurev.bb.08.060179.002033. PMID 383005.
^ Eckert, Roger; Naitoh, Yutaka (1972). "Control bioeléctrico de la locomoción en los ciliados*†". Revista de protozoología . 19 (2): 237–243. doi :10.1111/j.1550-7408.1972.tb03444.x. PMID 4624297.
^ Wood, David C. (1982). "Permeabilidades de membrana que determinan los potenciales de reposo, acción y mecanorreceptor en Stentor coeruleus". Journal of Comparative Physiology A . 146 (4): 537–550. doi :10.1007/bf00609450. S2CID 21083419.
^ Taylor, Alison R. (2009). "Un potencial de acción rápido basado en Na+/Ca2+ en una diatomea marina". PLOS ONE . 4 (3): e4966. Bibcode :2009PLoSO...4.4966T. doi : 10.1371/journal.pone.0004966 . PMC 2654917 . PMID 19305505.
^ Helliwell, Katherine E.; Chrachri, Abdul; Koester, Julie A.; Wharam, Susan; Verret, Frédéric; Taylor, Alison R.; Wheeler, Glen L.; Brownlee, Colin (2019). "Mecanismos alternativos para la señalización rápida de Na+/Ca2+ en eucariotas a través de una nueva clase de canales de dominio único controlados por voltaje". Current Biology . 29 (9): 1503–1511.e6. doi :10.1016/j.cub.2019.03.041. PMC 6509283 . PMID 31006567.
^ Bingley, MS; Thompson, CM (1962). "Potenciales bioeléctricos en relación con el movimiento en amebas". Journal of Theoretical Biology . 2 (1): 16–32. Bibcode :1962JThBi...2...16B. doi :10.1016/s0022-5193(62)80024-1.
^ Harz, Hartman; Hegemann, Peter (1991). "Corrientes de calcio reguladas por rodopsina en Chlamydomonas". Naturaleza . 351 (6326): 489–491. Código Bib :1991Natur.351..489H. doi :10.1038/351489a0. S2CID 4309593.
^ Fujiu, Kenta; Nakayama, Yoshitaka; Yanagisawa, Ayaka; Sokabe, Masahiro; Yoshimura, Kenjiro (2009). "Chlamydomonas CAV2 codifica un canal de calcio dependiente del voltaje necesario para la conversión de la forma de onda flagelar". Current Biology . 19 (2): 133–139. Bibcode :2009CBio...19..133F. doi : 10.1016/j.cub.2008.11.068 . PMID 19167228. S2CID 14063142.
^ Umbach JA (1981) "pH y excitabilidad de la membrana en Paramecium caudatum". Los Ángeles, CA: Universidad de California.
^ Dunlap, K. (1977). "Localización de los canales de calcio en Paramecium caudatum". Revista de Fisiología . 271 (1): 119–133. doi :10.1113/jphysiol.1977.sp011993. PMC 1353610 . PMID 915829.
^ Echevarría, Michael L.; Wolfe, Gordon V.; Taylor, Alison R. (2015). "Festín o huida: regulación bioeléctrica de los comportamientos de alimentación y evasión de depredadores en la especie planctónica alveolada Favella sp. (Spirotrichia)". Revista de biología experimental . 219 (Pt 3): 445–456. doi : 10.1242/jeb.121871 . PMID 26567352. S2CID 37255456.
^ Lueken, Wolfgang; Ricci, Nicola; Krüppel, Thomas (1996). "Las despolarizaciones espontáneas rítmicas determinan un ritmo lento y rápido en la marcha del hipotrico marino Euplotes vannus". Revista Europea de Protistología . 32 : 47–54. doi :10.1016/s0932-4739(96)80038-1.
^ Kunita, Itsuki; Kuroda, Shigeru; Ohki, Kaito; Nakagaki, Toshiyuki (2014). "Intentos de retirada de un largo capilar sin salida nadando hacia atrás en Paramecium". Frontiers in Microbiology . 5 : 270. doi : 10.3389/fmicb.2014.00270 . PMC 4052044 . PMID 24966852.
^ Stock C, KrÜPpel T, Key G, Lueken W (1999) "El comportamiento sexual en Euplotes raikovi está acompañado de modificaciones inducidas por feromonas de las corrientes iónicas". J Exp Biol , 202 (4): 475–483. PMID 9914154.
^ Kimball RF (1942) "La naturaleza y la herencia de los tipos de apareamiento en Euplotes Patella". Genética , 27 (3): 269–285. PMID 17247040, PMC PMC1209158.
^ Wood, DC (1988). "Habituación en Stentor: producida por modificación del canal mecanorreceptor". The Journal of Neuroscience . 8 (7): 2254–2258. doi :10.1523/JNEUROSCI.08-07-02254.1988. PMC 6569508 . PMID 3249223.
^ Jennings, HS (1899). "Estudios sobre reacciones a estímulos en organismos unicelulares. III Reacciones a estímulos localizados en Spirostomum y Stentor". The American Naturalist . 33 (389): 373–389. doi :10.1086/277256. S2CID 85272784.
^ Dexter, José P.; Prabakaran, Sudhakaran; Gunawardena, Jeremy (2019). "Una jerarquía compleja de comportamientos de evitación en un eucariota unicelular". Biología actual . 29 (24): 4323–4329.e2. Código Bib : 2019CBio...29E4323D. doi : 10.1016/j.cub.2019.10.059 . PMID 31813604. S2CID 208652463.
^ abc Akolpoglu, Mukrime Birgul; Dogan, Nihal Olcay; Bozuyuk, Ugur; Ceylan, Hakan; Kizilel, Seda; Sitti, Metin (2020). "Producción de alto rendimiento de microalgas biohíbridas para entrega de carga a pedido". Ciencia avanzada . 7 (16). doi :10.1002/advs.202001256. PMC 7435244 . PMID 32832367.El material fue copiado de esta fuente, que está disponible bajo una Licencia Creative Commons Atribución 4.0 Internacional.
^ Schwarz, Lukas; Medina-Sánchez, Mariana; Schmidt, Oliver G. (2017). "Micromotores híbridos biológicos". Applied Physics Reviews . 4 (3): 031301. Bibcode :2017ApPRv...4c1301S. doi : 10.1063/1.4993441 .
^ ab Bastos-Arrieta, Julio; Revilla-Guarinos, Ainhoa; Uspal, William E.; Simmchen, Juliane (2018). "Micronadadores biohíbridos bacterianos". Fronteras en robótica e inteligencia artificial . 5 : 97. doi : 10.3389/frobt.2018.00097 . PMC 7805739 . PMID 33500976.El material fue copiado de esta fuente, que está disponible bajo una Licencia Creative Commons Atribución 4.0 Internacional.
^ Montemagno, Carlo; Bachand, George (1999). "Construcción de dispositivos nanomecánicos alimentados por motores biomoleculares". Nanotecnología . 10 (3): 225–231. Bibcode :1999Nanot..10..225M. doi :10.1088/0957-4484/10/3/301. S2CID 250910730.
^ Ricotti, Leonardo; Trimmer, Barry; Feinberg, Adam W.; Raman, Ritu; Parker, Kevin K.; Bashir, Rashid; Sitti, Metin; Martel, Sylvain; Dario, Paolo; Menciassi, Arianna (2017). "Actuadores biohíbridos para robótica: una revisión de dispositivos activados por células vivas". Science Robotics . 2 (12): eaaq0495. doi : 10.1126/scirobotics.aaq0495 . PMID 33157905. S2CID 29776467.
^ Alapan, Yunus; Yasa, Oncay; Yigit, Berk; Yasa, I. Ceren; Erkoc, Pelin; Sitti, Metin (2019). "Microrrobótica y microorganismos: robots celulares autónomos biohíbridos". Revisión anual de control, robótica y sistemas autónomos . 2 : 205–230. doi :10.1146/annurev-control-053018-023803. S2CID 139819519.
^ Carlsen, Rika Wright; Sitti, Metin (2014). "Actuadores biohíbridos basados en células para microsistemas". Small . 10 (19): 3831–3851. doi :10.1002/smll.201400384. PMID 24895215.
^ Nguyen, Van Du; Han, Ji-Won; Choi, Young Jin; Cho, Sunghoon; Zheng, Shaohui; Ko, Seong Young; Park, Jong-Oh; Park, Sukho (2016). "Bacterobot liposomal terapéutico tumoral activo que combina un liposoma encapsulado con un fármaco (paclitaxel) con bacterias objetivo (Salmonella Typhimurium)". Sensores y actuadores B: Química . 224 : 217–224. doi :10.1016/j.snb.2015.09.034.
^ Felfoul, Ouajdi; Mohammadi, Mahmood; Taherkhani, Samira; De Lanauze, Dominic; Zhong Xu, Yong; Loghin, Dumitru; Essa, Sherief; Jancik, Sylwia; Houle, Daniel; Lafleur, Michel; Gaboury, Louis; Tabrizian, Maryam; Kaou, Neila; Atkin, Michael; Vuong, Té; Batist, Gerald; Beauchemin, Nicole; Radzioch, Danuta; Martel, Sylvain (2016). "Las bacterias magneto-aerotácticas entregan nanoliposomas que contienen fármacos a las regiones hipóxicas del tumor". Nature Nanotechnology . 11 (11): 941–947. Bibcode :2016NatNa..11..941F. doi :10.1038/nnano.2016.137. PMC 6094936 . Número de modelo: PMID27525475.
^ abcde Yasa, Oncay; Erkoc, Pelin; Alapan, Yunus; Sitti, Metin (2018). "Micronadadores propulsados por microalgas hacia la entrega activa de carga". Materiales avanzados . 30 (45): e1804130. Bibcode :2018AdM....3004130Y. doi :10.1002/adma.201804130. PMID 30252963. S2CID 52823884.
^ Ceylan, Hakan; Giltinan, Joshua; Kozielski, Kristen; Sitti, Metin (2017). "Microrobots móviles para aplicaciones de bioingeniería". Laboratorio en un chip . 17 (10): 1705–1724. doi : 10.1039/C7LC00064B . PMID 28480466.
^ Li, Jinxing; Esteban-Fernández De Ávila, Berta; Gao, Wei; Zhang, Liangfang; Wang, Joseph (2017). "Micro/Nanorobots para biomedicina: administración, cirugía, detección y desintoxicación". Science Robotics . 2 (4): eaam6431. doi :10.1126/scirobotics.aam6431. PMC 6759331 . PMID 31552379.
^ Park, Byung-Wook; Zhuang, Jiang; Yasa, Oncay; Sitti, Metin (2017). "Micronadadores multifuncionales impulsados por bacterias para la administración activa dirigida de fármacos". ACS Nano . 11 (9): 8910–8923. doi :10.1021/acsnano.7b03207. PMID 28873304.
^ Behkam, Bahareh; Sitti, Metin (2007). "Propulsión basada en flagelos bacterianos y control del movimiento de encendido/apagado de objetos a microescala". Applied Physics Letters . 90 (2): 023902. Bibcode :2007ApPhL..90b3902B. doi :10.1063/1.2431454.
^ Behkam, Bahareh; Sitti, Metin (2008). "Efecto de la cantidad y configuración de las bacterias adheridas en la propulsión bacteriana de microesferas". Applied Physics Letters . 93 (22): 223901. Bibcode :2008ApPhL..93v3901B. doi :10.1063/1.3040318.
^ Mostaghaci, Babak; Yasa, Oncay; Zhuang, Jiang; Sitti, Metin (2017). "Micronadadores bacterianos bioadhesivos para la administración dirigida de fármacos en los tractos urinario y gastrointestinal". Ciencia avanzada . 4 (6). doi :10.1002/advs.201700058. PMC 5473323 . PMID 28638787.
^ Schauer, Oliver; Mostaghaci, Babak; Colin, Remy; Hürtgen, Daniel; Kraus, David; Sitti, Metin; Sourjik, Victor (2018). "Motilidad y quimiotaxis de micronadadores impulsados por bacterias fabricados utilizando la visualización de biotina mediada por el antígeno 43". Scientific Reports . 8 (1): 9801. Bibcode :2018NatSR...8.9801S. doi :10.1038/s41598-018-28102-9. PMC 6023875 . PMID 29955099.
^ Singh, Ajay Vikram; Hosseinidoust, Zeinab; Park, Byung-Wook; Yasa, Oncay; Sitti, Metin (2017). "Micronadadores impulsados por bacterias blandas basados en microemulsiones para la entrega activa de carga". ACS Nano . 11 (10): 9759–9769. doi :10.1021/acsnano.7b02082. PMID 28858477.
^ Stanton, Morgan M.; Park, Byung-Wook; Miguel-López, Albert; Ma, Xing; Sitti, Metin; Sánchez, Samuel (2017). "Nadadores de microtubos biohíbridos impulsados por bacterias capturadas individualmente". Small . 13 (19). doi :10.1002/smll.201603679. hdl : 2445/123481 . PMID 28299891.
^ Stanton, Morgan M.; Park, Byung Wook; Vilela, Diana; Bente, Klaas; Faivre, Damián; Sitti, Metin; Sánchez, Samuel (2017). "Biohíbridos impulsados por bacterias magnetotácticas objetivo de biopelículas de E. Coli ". ACS Nano . 11 (10): 9968–9978. doi :10.1021/acsnano.7b04128. hdl : 2445/123493 . PMID 28933815.
^ ab Harris, Elizabeth H. (2001). "Chlamydomonas Amodelorganism". Revisión anual de fisiología vegetal y biología molecular vegetal . 52 : 363–406. doi :10.1146/annurev.arplant.52.1.363. PMID 11337403.
^ ab Weibel, DB; Garstecki, P.; Ryan, D.; Diluzio, WR; Mayer, M.; Seto, JE; Whitesides, GM (2005). "Microbueyes: microorganismos para mover cargas a microescala". Actas de la Academia Nacional de Ciencias . 102 (34): 11963–11967. Bibcode :2005PNAS..10211963W. doi : 10.1073/pnas.0505481102 . PMC 1189341 . PMID 16103369.
^ Hopfner, Úrsula; Schenck, Thilo-Ludwig; Chávez, Myra-Noemí; Machens, Hans-Günther; Bohne, Alexandra-Viola; Nickelsen, Jörg; Giunta, Riccardo-Enzo; Egaña, José-Tomás (2014). "Desarrollo de biomateriales fotosintéticos para ingeniería de tejidos in vitro". Acta Biomaterialia . 10 (6): 2712–2717. doi :10.1016/j.actbio.2013.12.055. PMID 24406198.
^ Centeno-Cerdas, Carolina; Jarquín-Cordero, Montserrat; Chávez, Myra Noemí; Hopfner, Úrsula; Holmes, Cristóbal; Schmauss, Daniel; Machens, Hans-Günther; Nickelsen, Jörg; Egaña, José Tomás (2018). "Desarrollo de suturas fotosintéticas para el suministro local de oxígeno y factores de crecimiento recombinantes en heridas". Acta Biomaterialia . 81 : 184-194. doi :10.1016/j.actbio.2018.09.060. PMID 30287280. S2CID 52922420.
^ Schenck, Thilo Ludwig; Hopfner, Úrsula; Chávez, Myra Noemí; Machens, Hans-Günther; Somlai-Schweiger, Ian; Giunta, Riccardo Enzo; Bohne, Alexandra Viola; Nickelsen, Jörg; Allende, Miguel L.; Egaña, José Tomás (2015). "Biomateriales fotosintéticos: un camino hacia la ingeniería de tejidos autótrofos". Acta Biomaterialia . 15 : 39–47. doi :10.1016/j.actbio.2014.12.012. PMID 25536030.







.jpg/1280px-Chlamydomonas_(10000x).jpg)





