Paranthropus boisei es una especie de australopiteco del Pleistoceno temprano de África Oriental hace unos 2,5 a 1,15 millones de años. [1] El espécimen holotipo , OH 5 , fue descubierto por la paleoantropóloga Mary Leakey en 1959 en Olduvai Gorge, Tanzania y descrito por su esposo Louis un mes después. Originalmente se lo colocó en su propio género como " Zinjanthropus boisei ", pero ahora se lo relega a Paranthropus junto con otros australopitecos robustos. Sin embargo, también se argumenta que Paranthropus es una agrupación inválida y sinónimo de Australopithecus , por lo que la especie también se clasifica a menudo como Australopithecus boisei .
Los australopitecos robustos se caracterizan por tener cráneos de constitución robusta capaces de producir altas tensiones y fuerzas de mordida , y algunos de los molares más grandes con el esmalte más grueso de cualquier simio conocido. P. boisei es el más robusto de este grupo. El tamaño del cerebro era de unos 450-550 cc (27-34 cu in), similar a otros australopitecos. Algunos cráneos son notablemente más pequeños que otros, lo que se toma como evidencia de dimorfismo sexual donde las hembras son mucho más pequeñas que los machos, aunque el tamaño del cuerpo es difícil de estimar dado que solo un espécimen, OH 80, definitivamente proporciona elementos corporales. El presunto macho OH 80 puede haber medido 156 cm (5 pies 1 pulgada) de alto y 61,7 kg (136 libras) de peso, y la presunta hembra KNM-ER 1500 124 cm (4 pies 1 pulgada) de alto (aunque su designación de especie no está clara). Los huesos del brazo y la mano de OH 80 y KNM-ER 47000 sugieren que P. boisei era arbóreo hasta cierto punto.
Originalmente se creía que P. boisei había sido una especie especialista en alimentos duros, como nueces, debido a su cráneo fuertemente construido, pero es más probable que fuera un alimentador generalista de plantas C4 predominantemente abrasivas , como pastos u órganos de almacenamiento subterráneos . Al igual que los gorilas , las adaptaciones aparentemente especializadas del cráneo pueden haber sido utilizadas solo con alimentos de reserva menos deseables, lo que le permitió a P. boisei habitar una gama más amplia de hábitats que los australopitecos gráciles. P. boisei puede haber sido capaz de fabricar herramientas de piedra olduvayenses y de matar cadáveres. P. boisei habitaba principalmente en ambientes húmedos y boscosos, y coexistía con H. habilis , H. rudolfensis y H. ergaster / erectus . Es probable que estos fueran presa de los grandes carnívoros de la época, incluidos los grandes felinos , los cocodrilos y las hienas .

Los paleoantropólogos Mary y Louis Leakey habían realizado excavaciones en Tanzania desde la década de 1930, aunque el trabajo se pospuso con el inicio de la Segunda Guerra Mundial . Regresaron en 1951 y encontraron principalmente herramientas antiguas y fósiles de mamíferos extintos para los siguientes años. [2] En 1955, desenterraron un canino bebé de homínido y un molar grande en Olduvai Gorge , ID de catálogo Olduvai Hominin (OH) 3. [3]
En la mañana del 17 de julio de 1959, Louis se sintió enfermo y se quedó en el campamento mientras Mary salía a Bed I's Frida Leakey Gully. [4] En algún momento alrededor de las 11:00 a. m., notó lo que parecía ser una parte de un cráneo que sobresalía del suelo, OH 5. [5] El equipo de excavación creó una pila de piedras alrededor de la parte expuesta para protegerla de una mayor erosión. [6] La excavación activa comenzó al día siguiente; habían decidido esperar a que el fotógrafo Des Bartlett documentara todo el proceso. [6] El cráneo parcial fue desenterrado por completo el 6 de agosto, aunque tuvo que ser reconstruido a partir de sus fragmentos que estaban esparcidos en el pedregal . [7] Louis publicó un breve resumen del hallazgo y el contexto la semana siguiente. [8]
Louis determinó que OH 5 era un subadulto o adolescente basándose en el desarrollo dental, y él y Mary lo apodaron "Dear Boy". [9] Después de que reconstruyeron el cráneo y las mandíbulas, los periódicos comenzaron a referirse a él como "El hombre cascanueces" debido a los grandes dientes posteriores y mandíbulas que le daban un parecido con los cascanueces antiguos . [10] El paleoantropólogo sudafricano Phillip Tobias , un colega de los Leakey, también recibió atribución por este apodo. [11] El cráneo fue llevado a Kenia después de su descubrimiento y estuvo allí hasta enero de 1965, cuando se exhibió en el Salón del Hombre en el Museo Nacional de Tanzania en Dar es Salaam. [12]
Louis supuso preliminarmente que el OH 5 tenía alrededor de medio millón de años, pero en 1965, los geólogos estadounidenses Garniss Curtis y Jack Evernden dataron el OH 5 en 1,75 millones de años atrás utilizando la datación de potasio-argón de cristales de anortoclasa de un lecho de toba (ceniza volcánica) suprayacente . [13] Tal aplicación de la geocronología no tenía precedentes en ese momento. [14]

La primera mandíbula identificada, Peninj 1 , fue descubierta en el lago Natron, justo al norte de Olduvai Gorge, en 1964. [15] : 107 [16] [17] Especialmente entre 1966 y 1975, se informó de varios especímenes más que revelaban elementos faciales en la Formación Shungura , Etiopía; Koobi Fora y Chesowanja , Kenia; y Omo y Konso , Etiopía. Entre los especímenes notables encontrados se incluye el cráneo bien conservado KNM-ER 406 de Koobi Fora en 1970. [15] : 108–109 En 1997, el primer espécimen con cráneo y mandíbula (y también uno de los especímenes más grandes), KGA10-525, fue descubierto en Konso. [18] En 1999, se recuperó una mandíbula en Malema, Malawi, extendiendo el rango más al sur de la especie a más de 2000 km (1200 mi) desde Olduvai Gorge. [15] : 109 Los primeros elementos corporales definitivos de P. boisei asociados con elementos faciales, OH 80 (dientes aislados con un brazo y una pierna), fueron descubiertos en 2013. Anteriormente, los restos corporales que carecían de elementos craneales diagnósticos inequívocos habían sido asignados dudosamente a la especie, a saber, el esqueleto parcial KNM-ER 1500 asociado con un pequeño fragmento de mandíbula. [19] En 2015, basándose en OH 80, el paleoantropólogo estadounidense Michael Lague recomendó asignar los especímenes de húmero aislados KNM-ER 739, 1504, 6020 y 1591 de Koobi Fora a P. boisei . [20] En 2020, se informaron los primeros huesos de la mano asociados, KNM-ER 47000 (que también incluye un brazo casi completo), de Ileret , Kenia. [21]
Los restos eran claramente australopitecos (no del género Homo ), y en ese momento, los únicos géneros australopitecos descritos eran Australopithecus por Raymond Dart y Paranthropus (el P. robustus sudafricano ) por Robert Broom , y hubo argumentos de que Paranthropus era sinónimo de Australopithecus . Louis creía que el cráneo tenía una mezcla de rasgos de ambos géneros, enumerando brevemente 20 diferencias, y por eso utilizó OH 5 como base para el nuevo género y especie " Zinjanthropus boisei " el 15 de agosto de 1959. El nombre del género deriva del término medieval para África Oriental, " Zanj ", y el nombre específico era en honor a Charles Watson Boise , el benefactor de los Leakey. [22] Inicialmente consideró el nombre " Titanohomo mirabilis " ("maravilloso hombre parecido a Titán"). [23]
Poco después, Louis presentó a " Z. " boisei en el 4º Congreso Panafricano de Prehistoria en Léopoldville, Congo Belga (ahora Kinshasa , República Democrática del Congo). [3] Dart hizo su ahora famoso chiste, "... qué habría pasado si [el espécimen de A. africanus ] la Sra. Ples hubiera conocido a Dear Boy una noche oscura". En el momento del descubrimiento, hubo resistencia a erigir géneros completamente nuevos basados en especímenes únicos, y el Congreso rechazó en gran medida a " Zinjanthropus ". [24] En 1960, el antropólogo estadounidense John Talbot Robinson señaló que las supuestas diferencias entre " Zinjanthropus " y Paranthropus se deben a que OH 5 es ligeramente más grande, por lo que recomendó que la especie se reclasificara como P. boisei . Louis rechazó la propuesta de Robinson. [25] Después de esto, se debatió si P. boisei era simplemente una variante de África Oriental de P. robustus hasta 1967, cuando el paleoantropólogo sudafricano Phillip V. Tobias dio una descripción mucho más detallada de OH 5 en una monografía (editada por Louis). Tobias y Louis todavía conservaron " Zinjanthropus ", pero recomendaron degradarlo al nivel de subgénero como Australopithecus ("Zinjanthropus") boisei , considerando que Paranthropus era sinónimo de Australopithecus . [3] La sinonimización de Paranthropus con Australopithecus fue sugerida por primera vez por los antropólogos Sherwood Washburn y Bruce D. Patterson en 1951, quienes recomendaron limitar los géneros de homínidos solo a Australopithecus y Homo . [26]
El género Paranthropus (también conocido como "australopitecos robustos") incluye típicamente a P. boisei , P. aethiopicus y P. robustus . Se debate si Paranthropus es una agrupación natural válida ( monofilético ) o una agrupación inválida de homínidos de apariencia similar ( parafilético ). Debido a que los elementos esqueléticos son tan limitados en estas especies, sus afinidades entre sí y con otros australopitecos es difícil de medir con precisión. Las mandíbulas son el principal argumento a favor de la monofilia, pero dicha anatomía está fuertemente influenciada por la dieta y el medio ambiente, y con toda probabilidad podría haber evolucionado independientemente en P. boisei y P. robustus . Los defensores de la monofilia consideran que P. aethiopicus es ancestral de las otras dos especies, o está estrechamente relacionado con el ancestro. Los defensores de la parafilia asignan estas tres especies al género Australopithecus como A. boisei , A. aethiopicus y A. robustus . [15] : 117-121
Antes de que se describiera a P. boisei (y P. robustus era el único miembro de Paranthropus ), Broom y Robinson seguían argumentando que P. robustus y A. africanus (los únicos australopitecos conocidos en aquel momento) eran dos linajes distintos. Sin embargo, los restos no estaban firmemente datados y se debatía si había de hecho múltiples linajes de homínidos o si solo había uno que condujera a los humanos. En 1975, se demostró que el cráneo de P. boisei KNM-ER 406 era contemporáneo del cráneo de H. ergaster / erectus KNM ER 3733 , lo que generalmente se considera que demuestra que Paranthropus era un taxón hermano de Homo , ambos desarrollándose a partir de algunas especies de Australopithecus , que en ese momento solo incluían a A. africanus . En 1979, un año después de describir a A. afarensis de África Oriental, los antropólogos Donald Johanson y Tim D. White sugirieron que A. afarensis era en cambio el último ancestro común entre Homo y Paranthropus , y A. africanus era el miembro más antiguo del linaje Paranthropus o al menos era ancestral de P. robustus , porque A. africanus habitó Sudáfrica antes que P. robustus , y A. afarensis era en ese momento la especie de homínido más antigua conocida con aproximadamente 3,5 millones de años. [27] Ahora, el australopiteco sudafricano más antiguo conocido (" Little Foot ") data de hace 3,67 millones de años, contemporáneo con A. afarensis . [28]
Tales argumentos se basan en cómo se dibuja el árbol genealógico de los homínidos, y la clasificación exacta de las especies de Australopithecus entre sí es bastante polémica. Por ejemplo, si se considera al sudafricano A. sediba (que evolucionó a partir de A. africanus ) como el ancestro o estrechamente relacionado con el ancestro de Homo , esto podría permitir que A. africanus se coloque más estrechamente relacionado con Homo que con Paranthropus . Esto dejaría al etíope A. garhi como el ancestro de P. aethiopicus en lugar de A. africanus (asumiendo que Paranthropus es monofilético y que P. aethiopicus evolucionó en un momento en África Oriental cuando solo existía A. garhi allí). [29]
Dado que tanto P. boisei como P. aethiopicus son conocidos en África Oriental y que P. aethiopicus solo se ha identificado con seguridad a partir del cráneo KNM WT 17000 y de unas pocas mandíbulas y dientes aislados, se debate si P. aethiopicus debería incluirse en P. boisei o si las diferencias derivadas de su arcaicidad justifican la distinción entre especies. Los términos P. boisei sensu lato ("en sentido amplio") y P. boisei sensu stricto ("en sentido estricto") pueden utilizarse para incluir y excluir respectivamente a P. aethiopicus de P. boisei cuando se habla del linaje en su conjunto. [15] : 106–107
P. aethiopicus es el miembro más antiguo del género, con los restos más antiguos, de la Formación Omo Kibish etíope , que datan de hace 2,6 millones de años (mya) al final del Plioceno . [30] Es posible que P. aethiopicus evolucionara incluso antes, hasta 3,3 mya, en las extensas llanuras aluviales de Kenia de la época. [31] Los restos más antiguos de P. boisei datan de alrededor de 2,3 mya de Malema. [30] El registro más reciente de P. boisei proviene de Olduvai Gorge (OH 80) hace unos 1,34 mya; [19] sin embargo, debido a una gran brecha en el registro fósil de homínidos, P. boisei puede haber persistido hasta 1 mya. [15] : 109 P. boisei cambió notablemente poco durante su existencia de casi un millón de años. [32]

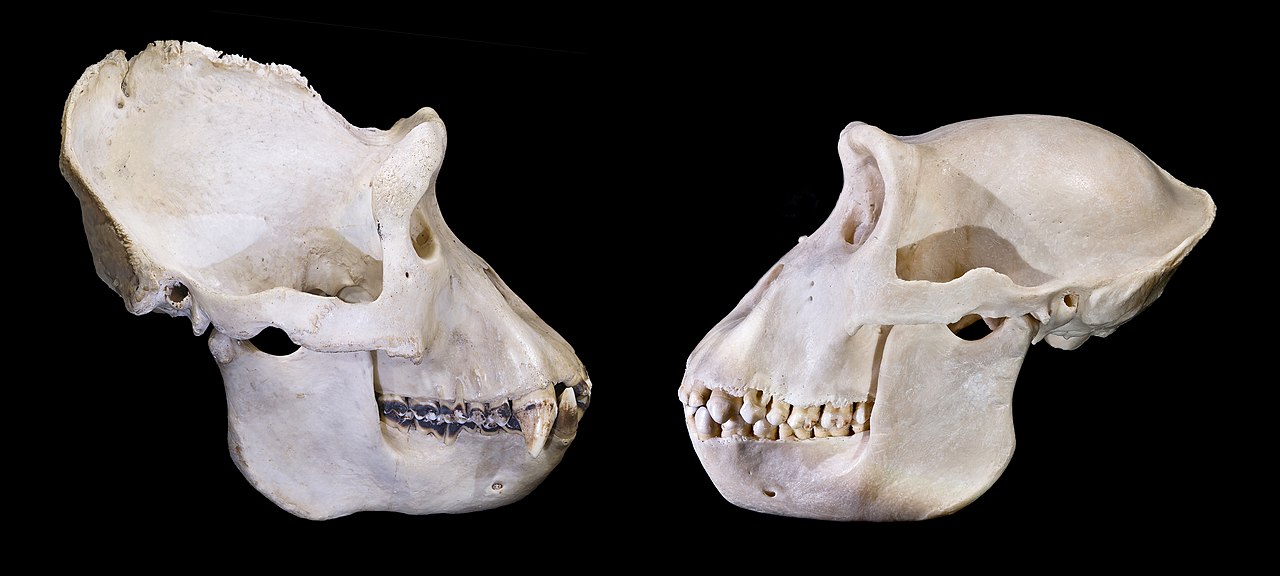
P. boisei es el más robusto de los robustos australopitecos, mientras que el sudafricano P. robustus es más pequeño con rasgos comparativamente más gráciles. [15] : 120 El cráneo de P. boisei es de constitución robusta y presenta una cresta superciliar definida , frente retraída, márgenes inferiores redondeados de las cuencas de los ojos , pómulos inflados y cóncavos , un paladar grueso y una mandíbula robusta y profunda. Esto generalmente se interpreta como que le permitió a P. boisei resistir altas tensiones mientras masticaba, [33] aunque el paladar grueso podría ser en cambio un subproducto del alargamiento facial. [34] El cráneo presenta grandes parches ásperos (rugosidades) en las mejillas y las mandíbulas, y los machos tienen crestas sagitales (en la línea media) y temporonucal (en la espalda) pronunciadas, que indican un músculo masetero masivo (usado para morder) colocado cerca de la parte delantera de la cabeza (aumentando la ventaja mecánica ). Esto generalmente se considera evidencia de una alta fuerza de mordida . [33]
Los incisivos y caninos son reducidos, lo que dificultaría la mordida de trozos grandes de comida. En contraste, los dientes de las mejillas de ambos sexos son enormes ( megadoncia postcanina ), y la mayor área de superficie habría permitido el procesamiento de mayores cantidades de comida a la vez. [33] En la mandíbula superior, el primer molar promedia aproximadamente 250 mm2 ( 0,39 pulgadas cuadradas), el segundo molar 320 mm2 ( 0,50 pulgadas cuadradas) y el tercer molar 315 mm2 ( 0,488 pulgadas cuadradas); en la mandíbula inferior, el primer molar promedia aproximadamente 260 mm2 ( 0,40 pulgadas cuadradas), el segundo molar 315 mm2 ( 0,488 pulgadas cuadradas) y el tercer molar 340 mm2 ( 0,53 pulgadas cuadradas). [35] Los molares son bunodontes , con cúspides bajas y redondeadas . Los premolares se parecen a las muelas (están molarizados), lo que puede indicar que P. boisei necesitaba una superficie de masticación extendida para procesar una gran cantidad de alimentos al mismo tiempo. El esmalte de los molares se encuentra entre los más gruesos de cualquier simio conocido, lo que ayudaría a resistir altas tensiones al morder. [33] : 128–132

En una muestra de 10 especímenes de P. boisei , el tamaño del cerebro varió de 444 a 545 cc (27,1 a 33,3 pulgadas cúbicas) con un promedio de 487,5 cc (29,75 pulgadas cúbicas). [36] Sin embargo, el espécimen de extremo inferior, Omo L338‐y6, es un juvenil, y muchos especímenes de cráneo tienen un hueso frontal muy dañado o faltante , lo que puede alterar las estimaciones del volumen cerebral. [37] El volumen cerebral de los australopitecos generalmente varió de 400 a 500 cc (24 a 31 pulgadas cúbicas), y para el Homo contemporáneo de 500 a 900 cc (31 a 55 pulgadas cúbicas). [38]
En cuanto a los senos venosos durales , en 1983, el neuroantropólogo estadounidense Dean Falk y el antropólogo Glenn Conroy sugirieron que, a diferencia de A. africanus o los humanos modernos, todos los Paranthropus (y A. afarensis ) tenían senos occipitales y marginales (alrededor del foramen magnum ) expandidos, suplantando por completo los senos transversos y sigmoideos . En 1988, Falk y Tobias demostraron que los homínidos pueden tener sistemas occipital/marginal y transverso/sigmoideo simultáneamente o en mitades opuestas del cráneo, como en el espécimen KNM-ER 23000 de P. boisei . [39]

En 1983, el antropólogo francés Roger Saban afirmó que la rama parietal de la arteria meníngea media se originó a partir de la rama posterior en P. boisei y P. robustus en lugar de la rama anterior como en los homínidos anteriores, y consideró que esto era una característica derivada debido a una mayor capacidad cerebral. [40] Desde entonces se ha demostrado que la rama parietal podría originarse de las ramas anterior o posterior, a veces ambas en un solo espécimen en lados opuestos del cráneo como en KNM-ER 23000 y OH 5. [41]
La amplia gama de variación de tamaño en los especímenes de cráneo parece indicar un alto grado de dimorfismo sexual , siendo los machos notablemente más grandes que las hembras. Sin embargo, es difícil predecir con precisión las dimensiones reales de los machos y hembras vivos debido a la falta de restos esqueléticos definitivos de P. boisei , salvo el presunto macho OH 80. Basándonos en una aproximación de 400 mm (1,3 pies) para el fémur antes de que se rompiera y utilizando proporciones similares a las de los humanos modernos (lo que probablemente sea una suposición insegura), OH 80 medía aproximadamente 156,3 cm (5 pies 1,5 pulgadas) de alto en vida. [19] A modo de comparación, los hombres y mujeres humanos modernos en el año 1900 medían en promedio 163 cm (5 pies 4 pulgadas) y 152,7 cm (5,01 pies), respectivamente. [42] Falta la cabeza femoral , el mejor indicador para estimar la masa corporal, pero usando el eje, OH 80 pesaba alrededor de 50 kg (110 lb) asumiendo proporciones humanas, y 61,7 kg (136 lb) usando las proporciones de un simio no humano. [19] Se estima que el fémur femenino KNM-ER 1500, atribuido de forma ambigua y presuntamente femenino, era de un individuo de unos 124 cm (4 pies 1 pulgada) de alto [43], lo que sería consistente con el argumento del dimorfismo sexual, [19] pero si el espécimen pertenece de hecho a P. boisei , mostraría una anatomía de las extremidades bastante similar a la del H. habilis contemporáneo . [15] : 116

En cambio, el fémur OH 80, más parecido al femora de H. erectus , es bastante grueso, presenta un eje aplanado lateralmente e indica líneas glúteas , pectíneas e intertrocantéreas dispuestas de manera similar alrededor de la articulación de la cadera . No obstante, la línea intertrocantérea está mucho más definida en OH 80, la tuberosidad glútea está más hacia la línea media del fémur y el eje medio en la vista lateral es más recto, lo que probablemente refleja alguna diferencia en las capacidades de carga de la pierna. A diferencia de P. robustus , los huesos del brazo de OH 80 están fuertemente construidos y la articulación del codo muestra similitudes con la de los gibones y orangutanes modernos . Esto podría indicar que P. boisei usaba una combinación de caminar terrestre y comportamiento suspensorio , o era completamente bípedo pero conservaba una condición de la parte superior del cuerpo similar a la de los simios de algunas especies ancestrales debido a la falta de presión selectiva para perderlos. Por el contrario, la mano de P. robustus no es compatible con la escalada. [19] La mano de KNM-ER 47000 muestra una anatomía similar a la de Australopithecus, ya que carece del proceso estiloides del tercer metacarpiano (que permite que la mano se bloquee en la muñeca para ejercer más presión), un pulgar débil en comparación con los humanos modernos y falanges curvadas (huesos de los dedos) que generalmente se interpretan como adaptaciones para la escalada. No obstante, a pesar de carecer de un agarre de precisión particularmente fuerte como el Homo , la mano aún era lo suficientemente diestra para manejar y fabricar herramientas simples. [21]
En 1954, Robinson sugirió que el cráneo corpulento de Paranthropus (que en ese momento solo incluía a P. robustus ) era indicativo de una dieta especializada específicamente adaptada para procesar una banda estrecha de alimentos. Debido a esto, el modelo predominante de extinción de Paranthropus durante la segunda mitad del siglo XX fue que no pudo adaptarse al clima volátil del Pleistoceno , a diferencia del mucho más adaptable Homo . [33] También se pensó alguna vez que P. boisei abría nueces y alimentos duros similares con sus poderosos dientes, lo que le dio a OH 5 el apodo de "Hombre Cascanueces". [44]
Sin embargo, en 1981, el antropólogo inglés Alan Walker encontró que los patrones de microdesgaste en los molares eran inconsistentes con una dieta alta en alimentos duros, y eran efectivamente indistinguibles del patrón visto en los molares de mandriles , chimpancés y orangutanes frugívoros . [45] El microdesgaste en los molares de P. boisei es diferente al de los molares de P. robustus , e indica que P. boisei , a diferencia de P. robustus , muy rara vez comía alimentos duros. Los análisis de isótopos de carbono informan una dieta predominantemente de plantas C4 , como pastos y juncos abrasivos y de baja calidad. [46] [47] [48] El esmalte grueso es consistente con la molienda de alimentos abrasivos. [33] Los patrones de microdesgaste en P. robustus se han examinado a fondo, y sugieren que la constitución pesada del cráneo solo era relevante cuando se comían alimentos de reserva menos deseables. Un esquema similar puede haber sido utilizado por P. boisei . [33] Esta estrategia es similar a la utilizada por los gorilas modernos , que pueden mantenerse enteramente con alimentos de reserva de menor calidad durante todo el año, a diferencia de los chimpancés de complexión más ligera (y presumiblemente los australopitecos gráciles) que requieren un acceso constante a alimentos de alta calidad. [49]

En 1980, los antropólogos Tom Hatley y John Kappelman sugirieron que los primeros homínidos ( convergentemente con los osos y los cerdos ) se adaptaron a comer órganos de almacenamiento subterráneo (USO) abrasivos y ricos en calorías, como raíces y tubérculos. [50] Desde entonces, la explotación de USO por parte de los homínidos ha ganado más apoyo. En 2005, los antropólogos biológicos Greg Laden y Richard Wrangham propusieron que Paranthropus dependía de los USO como una fuente de alimento de reserva o posiblemente primaria, y señalaron que puede haber una correlación entre la alta abundancia de USO y la ocupación de los homínidos. [49] En este modelo, P. boisei puede haber sido un alimentador generalista con predilección por los USO, [51] [48] y puede haberse extinguido debido a una tendencia a la aridez y una disminución resultante de los USO junto con la creciente competencia con los babuinos y el Homo . [52] Al igual que los chimpancés y babuinos modernos, los australopitecos probablemente buscaban alimento en las horas más frescas de la mañana y de la tarde en lugar de en el calor del día. [53]
Cuando se descubrió OH 5, los Leakey habían pasado 24 años excavando el área en busca de restos de homínidos tempranos, pero en cambio habían recuperado principalmente otros restos animales, así como la industria de herramientas de piedra de Oldowan . [3] Debido a que OH 5 estaba asociado con las herramientas y los huesos animales procesados, presumieron que era el fabricante de herramientas. La atribución de las herramientas se cambió rápidamente al H. habilis de cerebro más grande tras su descripción en 1964. [3] En 2013, se encontró OH 80 asociado con una masa de herramientas de piedra de Oldowan y huesos de animales que mostraban evidencia de carnicería. Esto podría indicar potencialmente que P. boisei fabricaba esta industria y comía carne en algún grado. [19]
Además, la Edad de Piedra Temprana de África coincide con herramientas de hueso simples . En Sudáfrica, estas se desentierran en la Cuna de la Humanidad y se atribuyen en gran parte a P. robustus . En África Oriental, se han encontrado algunas en los lechos I-IV de la garganta de Olduvai, que se produjeron hace aproximadamente entre 1,7 y 0,8 millones de años, y generalmente están hechas de huesos de extremidades y posiblemente dientes de grandes mamíferos, sobre todo elefantes . La infrecuencia de animales tan grandes en este sitio puede explicar la relativa rareza de las herramientas de hueso. Los fabricantes de herramientas modificaban el hueso de la misma manera que lo hacían con la piedra. Aunque las herramientas de hueso de Olduva normalmente se atribuyen a H. ergaster / erectus , la presencia tanto de P. boisei como de H. habilis confunde la atribución. [54]
En 1979, el antropólogo biológico estadounidense Noel T. Boaz observó que las proporciones relativas entre las grandes familias de mamíferos de la Formación Shungura son bastante similares a las que existen en la actualidad en el África subsahariana. Boaz creía que los homínidos habrían tenido aproximadamente la misma densidad de población que otros grandes mamíferos, lo que equivaldría a 0,006-1,7 individuos por kilómetro cuadrado (0,4 millas cuadradas). Alternativamente, al multiplicar la densidad de bóvidos, elefantes o hipopótamos por el porcentaje de restos de homínidos respecto del total de restos de mamíferos encontrados en la formación, Boaz estimó una densidad de 0,001-2,58 individuos por kilómetro cuadrado. [55] El biólogo Robert A. Martin consideró endebles los modelos de población basados en el número de especímenes conocidos. En 1981, Martin aplicó ecuaciones formuladas por los ecólogos Alton S. Harestad y Fred L. Bunnel en 1979 para estimar el área de distribución y la densidad de población de los grandes mamíferos basándose en el peso y la dieta, y, utilizando un peso de 52,4 kg (116 lb), obtuvo: 130 ha (320 acres) y 0,769 individuos por kilómetro cuadrado si eran herbívoros; 1295 ha (3200 acres) y 0,077 individuos si eran omnívoros; y 287 819 ha (711 220 acres) y 0,0004 individuos si eran carnívoros. A modo de comparación, calculó 953 ha (2350 acres) y 0,104 individuos por kilómetro cuadrado para chimpancés omnívoros de 37,5 kilogramos (83 lb). [56]
Un estudio de 2017 postuló que, debido a que los grandes simios no humanos machos tienen una cresta sagital más grande que las hembras (particularmente los gorilas y los orangutanes), la cresta puede estar influenciada por la selección sexual además de sostener los músculos masticadores. Además, el tamaño de la cresta sagital (y los músculos de los glúteos ) en los gorilas de las tierras bajas occidentales machos se ha correlacionado con el éxito reproductivo. Extendieron su interpretación de la cresta a los machos de especies de Paranthropus , con la cresta y la cabeza resultante más grande (al menos en P. boisei ) siendo utilizadas para algún tipo de exhibición . Esto contrasta con otros primates que exhiben los caninos típicamente congestionados en exhibición agonística (los caninos de Paranthropus son comparativamente pequeños). Sin embargo, también es posible que los gorilas y orangutanes machos requieran músculos temporales más grandes para lograr una abertura más amplia para exhibir mejor los caninos. [57]
En general, se considera que los australopitecos tuvieron una tasa de crecimiento más rápida, similar a la de los simios , que la de los humanos modernos, en gran medida debido a las tendencias de desarrollo dental. En términos generales, la aparición del primer molar permanente en los primeros homínidos se ha estimado de diversas formas entre los 2,5 y los 4,5 años de edad, lo que contrasta notablemente con el promedio de los humanos modernos de 5,8 años. Las puntas de las cúspides mesiales del primer molar (en el lado más cercano al premolar) de KNM-ER 1820 estaban aproximadamente al mismo nivel que el cuello uterino (donde el esmalte se encuentra con el cemento ) de su segundo premolar no permanente. En los babuinos, esta etapa ocurre cuando el primer molar está a punto de erupcionar de las encías. La raíz del diente mide aproximadamente 5 mm (0,20 pulgadas), lo que es similar a la de la mayoría de los demás homínidos en esta etapa. En contraste, la raíz del espécimen SK 62 de P. robustus medía 6 mm (0,24 pulgadas) cuando emergía a través del alvéolo dental (una etapa de desarrollo más temprana que la emergencia de la encía), por lo que, a menos que alguno de los especímenes sea anormal, P. robustus puede haber tenido una tasa de formación de raíz dentaria más alta. El primer molar del espécimen puede haber erupcionado 2-3 meses antes de la muerte, por lo que posiblemente a los 2,7-3,3 años de edad. En los simios modernos (incluidos los humanos), la trayectoria del desarrollo dental está fuertemente correlacionada con la historia de vida y la tasa de crecimiento general, pero es posible que los primeros homínidos simplemente tuvieran una trayectoria dental más rápida y una historia de vida más lenta debido a factores ambientales, como la edad de destete temprana exhibida en los lémures índridos modernos . [58]
Los restos de P. boisei se han encontrado predominantemente en lo que eran ambientes húmedos y boscosos, como humedales a lo largo de lagos y ríos, matorrales boscosos o áridos y bosques semiáridos, [48] con la excepción de los lechos Chiwondo de Malawi dominados por la sabana. [59] Su abundancia probablemente aumentó durante los períodos de humedad relativa impulsados por la precesión, mientras que fue más rara durante los intervalos de aridez. [60] Durante el Pleistoceno, parece haber habido bosques costeros y montañosos en África Oriental. Los valles fluviales más expansivos, a saber, el valle del río Omo , pueden haber servido como refugios importantes para las criaturas que habitan en los bosques. Al estar separados de los bosques de África Central por un corredor de sabana, estos bosques de África Oriental habrían promovido altas tasas de endemismo , especialmente durante épocas de volatilidad climática. [61] Los australopitecos y los primeros Homo probablemente preferían condiciones más frías que los posteriores Homo , ya que no hay yacimientos de australopitecos que estuvieran por debajo de los 1000 m (3300 pies) de altitud en el momento de la deposición. Esto significaría que, al igual que los chimpancés, a menudo habitaban zonas con una temperatura media diurna de 25 °C (77 °F), que descendía a 10 o 5 °C (50 o 41 °F) por la noche. [53]
P. boisei coexistió con H. habilis , H. rudolfensis y H. ergaster / erectus , pero no está claro cómo interactuaron. [3] Para explicar por qué P. boisei se asoció con herramientas olduvayenses a pesar de no ser el fabricante de herramientas, Louis Leakey y sus colegas, al describir a H. habilis en 1964, sugirieron que una posibilidad era que P. boisei fuera asesinado por H. habilis , [62] quizás como alimento. [63] Sin embargo, al describir a P. boisei 5 años antes, dijo: "No hay razón alguna, en este caso, para creer que el cráneo [OH 5] represente a la víctima de un festín caníbal por parte de algún tipo hipotético de hombre más avanzado". [22] OH 80 parece haber sido devorado por un gran felino. [64] La pata OH 35, que pertenece a P. boisei o H. habilis , muestra evidencia de depredación por leopardo . [65] Otros posibles depredadores olduvayenses de los grandes simios incluyen a la hiena cazadora Chasmaporthetes nitidula , los felinos dientes de sable Dinofelis y Megantereon , [66] y el cocodrilo Crocodylus anthropophagus . [67]
La mandíbula fue descubierta por Kamoya Kimeu en 1964, durante una expedición dirigida por Richard Leakey y Glynn Isaac.