
El comportamiento de enjambre , o enjambre , es un comportamiento colectivo que exhiben entidades, particularmente animales, de tamaño similar que se agrupan, tal vez pululando en el mismo lugar o tal vez moviéndose en masa o migrando en alguna dirección. Es un tema altamente interdisciplinario. [1]
Como término, el término enjambre se aplica particularmente a los insectos, pero también puede aplicarse a cualquier otra entidad o animal que exhiba un comportamiento de enjambre. El término bandada o murmuración puede referirse específicamente al comportamiento de enjambre en las aves, manada para referirse al comportamiento de enjambre en los tetrápodos , y cardumen o cardumen para referirse al comportamiento de enjambre en los peces. El fitoplancton también se reúne en enormes enjambres llamados floraciones , aunque estos organismos son algas y no se autopropulsan como lo hacen los animales. Por extensión, el término "enjambre" se aplica también a entidades inanimadas que exhiben comportamientos paralelos, como en un enjambre de robots , un enjambre de terremotos o un enjambre de estrellas.
Desde un punto de vista más abstracto, el comportamiento de enjambre es el movimiento colectivo de un gran número de entidades autopropulsadas . [2] Desde la perspectiva del modelador matemático, es un comportamiento emergente que surge de reglas simples que son seguidas por individuos y no involucra ninguna coordinación central. El comportamiento de enjambre también es estudiado por los físicos de materia activa como un fenómeno que no está en equilibrio termodinámico , y como tal requiere el desarrollo de herramientas más allá de las disponibles en la física estadística de sistemas en equilibrio termodinámico. En este sentido, el enjambre ha sido comparado con las matemáticas de los superfluidos , específicamente en el contexto de las bandadas de estorninos (murmuración). [3]
El comportamiento de los enjambres se simuló por primera vez en un ordenador en 1986 con el programa de simulación boids . [4] Este programa simula agentes simples (boids) a los que se les permite moverse de acuerdo con un conjunto de reglas básicas. El modelo fue diseñado originalmente para imitar el comportamiento de las bandadas de pájaros, pero también se puede aplicar a bancos de peces y otras entidades que forman enjambres.
En las últimas décadas, los científicos han recurrido a modelar el comportamiento de los enjambres para comprender más profundamente su comportamiento.

Los primeros estudios sobre el comportamiento de los enjambres emplearon modelos matemáticos para simular y comprender el comportamiento. Los modelos matemáticos más simples de enjambres de animales generalmente representan a los animales individuales siguiendo tres reglas:
El programa informático Boids , creado por Craig Reynolds en 1986, simula el comportamiento de un enjambre siguiendo las reglas anteriores. [4] Muchos modelos posteriores y actuales utilizan variaciones de estas reglas, a menudo implementándolas mediante "zonas" concéntricas alrededor de cada animal. En la "zona de repulsión", muy cerca del animal, el animal focal intentará distanciarse de sus vecinos para evitar la colisión. Un poco más lejos, en la "zona de alineación", el animal focal intentará alinear su dirección de movimiento con sus vecinos. En la "zona de atracción" más externa, que se extiende tan lejos del animal focal como sea capaz de percibir, el animal focal intentará moverse hacia un vecino.
La forma de estas zonas se verá necesariamente afectada por las capacidades sensoriales de un animal determinado. Por ejemplo, el campo visual de un ave no se extiende detrás de su cuerpo. Los peces dependen tanto de la visión como de las percepciones hidrodinámicas transmitidas a través de sus líneas laterales , mientras que el krill antártico depende tanto de la visión como de las señales hidrodinámicas transmitidas a través de las antenas .
Sin embargo, estudios recientes sobre bandadas de estorninos han demostrado que cada ave modifica su posición en relación con los seis o siete animales que la rodean directamente, sin importar cuán cerca o cuán lejos estén esos animales. [5] Las interacciones entre bandadas de estorninos se basan, por lo tanto, en una regla topológica , en lugar de métrica. Queda por ver si esto se aplica a otros animales. Otro estudio reciente, basado en un análisis de imágenes de bandadas tomadas con cámaras de alta velocidad sobre Roma y asumiendo reglas de comportamiento mínimas, ha simulado de manera convincente varios aspectos del comportamiento de las bandadas. [6] [7] [8] [9]
Para comprender mejor por qué los animales desarrollan comportamientos de enjambre, los científicos han recurrido a modelos evolutivos que simulan poblaciones de animales en evolución. Normalmente, estos estudios utilizan un algoritmo genético para simular la evolución a lo largo de muchas generaciones. Estos estudios han investigado una serie de hipótesis que intentan explicar por qué los animales desarrollan comportamientos de enjambre, como la teoría de la manada egoísta [10] [11] [12] [13] [14] , el efecto de confusión del depredador [15] [16] , el efecto de dilución [17] [18] y la teoría de los muchos ojos [19] .

El concepto de emergencia (que las propiedades y funciones que se encuentran en un nivel jerárquico no están presentes y son irrelevantes en los niveles inferiores) es a menudo un principio básico detrás de los sistemas autoorganizados . [20] Un ejemplo de autoorganización en biología que conduce a la emergencia en el mundo natural ocurre en las colonias de hormigas. La reina no da órdenes directas ni les dice a las hormigas qué hacer. [ cita requerida ] En cambio, cada hormiga reacciona a estímulos en forma de olores químicos de larvas, otras hormigas, intrusos, comida y acumulación de desechos, y deja un rastro químico que, a su vez, proporciona un estímulo a otras hormigas. Aquí cada hormiga es una unidad autónoma que reacciona dependiendo solo de su entorno local y de las reglas codificadas genéticamente para su variedad. A pesar de la falta de toma de decisiones centralizada, las colonias de hormigas exhiben comportamientos complejos e incluso han podido demostrar la capacidad de resolver problemas geométricos. Por ejemplo, las colonias encuentran rutinariamente la distancia máxima desde todas las entradas de la colonia para deshacerse de los cadáveres.
Otro concepto clave en el campo de la inteligencia de enjambre es la estigmergia . [21] [22] La estigmergia es un mecanismo de coordinación indirecta entre agentes o acciones. El principio es que el rastro dejado en el entorno por una acción estimula la realización de una siguiente acción, por el mismo agente o por uno diferente. De esa manera, las acciones posteriores tienden a reforzarse y apoyarse mutuamente, lo que lleva al surgimiento espontáneo de una actividad coherente y aparentemente sistemática. La estigmergia es una forma de autoorganización. Produce estructuras complejas, aparentemente inteligentes, sin necesidad de ninguna planificación, control o incluso comunicación directa entre los agentes. Como tal, apoya la colaboración eficiente entre agentes extremadamente simples, que carecen de memoria, inteligencia o incluso conciencia de los demás. [22]
La inteligencia de enjambre es el comportamiento colectivo de sistemas descentralizados y autoorganizados , naturales o artificiales. El concepto se emplea en trabajos sobre inteligencia artificial . La expresión fue introducida por Gerardo Beni y Jing Wang en 1989, en el contexto de los sistemas robóticos celulares . [23]
Los sistemas de inteligencia de enjambre suelen estar formados por una población de agentes simples , como los boids, que interactúan localmente entre sí y con su entorno. Los agentes siguen reglas muy simples y, aunque no existe una estructura de control centralizada que dicte cómo deben comportarse los agentes individuales, las interacciones locales y, en cierta medida, aleatorias entre dichos agentes conducen al surgimiento de un comportamiento global inteligente, desconocido para los agentes individuales.
La investigación sobre inteligencia de enjambre es multidisciplinaria. Puede dividirse en investigación de enjambre natural que estudia los sistemas biológicos e investigación de enjambre artificial que estudia los artefactos humanos. También existe una corriente científica que intenta modelar los sistemas de enjambre en sí y comprender sus mecanismos subyacentes, y una corriente de ingeniería centrada en aplicar los conocimientos desarrollados por la corriente científica para resolver problemas prácticos en otras áreas. [24]
Los algoritmos de enjambre siguen un enfoque lagrangiano o un enfoque euleriano . [25] El enfoque euleriano considera el enjambre como un campo , trabajando con la densidad del enjambre y derivando propiedades de campo promedio. Es un enfoque hidrodinámico y puede ser útil para modelar la dinámica general de enjambres grandes. [26] [27] [28] Sin embargo, la mayoría de los modelos funcionan con el enfoque lagrangiano, que es un modelo basado en agentes que sigue a los agentes individuales (puntos o partículas) que componen el enjambre. Los modelos de partículas individuales pueden seguir información sobre el rumbo y el espaciamiento que se pierde en el enfoque euleriano. [25] [29]
La optimización de colonias de hormigas es un algoritmo ampliamente utilizado que se inspiró en los comportamientos de las hormigas y ha sido eficaz para resolver problemas de optimización discretos relacionados con la formación de enjambres. [31] El algoritmo fue propuesto inicialmente por Marco Dorigo en 1992, [32] [33] y desde entonces se ha diversificado para resolver una clase más amplia de problemas numéricos. Las especies que tienen múltiples reinas pueden tener una reina que abandona el nido junto con algunas obreras para fundar una colonia en un nuevo sitio, un proceso similar al enjambre de las abejas . [34] [35]
El concepto de partículas autopropulsadas (SPP) fue introducido en 1995 por Tamás Vicsek et al. [37] como un caso especial del modelo de boids introducido en 1986 por Reynolds. [4] Un enjambre SPP está modelado por una colección de partículas que se mueven con una velocidad constante y responden a perturbaciones aleatorias adoptando en cada incremento de tiempo la dirección promedio de movimiento de las otras partículas en su vecindario local. [38]
Las simulaciones demuestran que una "regla del vecino más próximo" adecuada acaba dando lugar a que todas las partículas se agrupen o se muevan en la misma dirección. Esto se produce a pesar de que no existe una coordinación centralizada y de que los vecinos de cada partícula cambian constantemente con el tiempo. [37] Los modelos SPP predicen que los animales que forman enjambres comparten ciertas propiedades a nivel de grupo, independientemente del tipo de animales que formen parte del enjambre. [39] Los sistemas de enjambres dan lugar a comportamientos emergentes que se producen en muchas escalas diferentes, algunos de los cuales son universales y robustos. Se ha convertido en un reto en física teórica encontrar modelos estadísticos mínimos que capturen estos comportamientos. [40] [41]
La optimización de enjambre de partículas es otro algoritmo ampliamente utilizado para resolver problemas relacionados con enjambres. Fue desarrollado en 1995 por Kennedy y Eberhart y primero tuvo como objetivo simular el comportamiento social y la coreografía de bandadas de aves y bancos de peces. [42] [43] El algoritmo se simplificó y se observó que realizaba optimización. El sistema inicialmente siembra una población con soluciones aleatorias. Luego busca en el espacio del problema a través de generaciones sucesivas utilizando optimización estocástica para encontrar las mejores soluciones. Las soluciones que encuentra se denominan partículas . Cada partícula almacena su posición, así como la mejor solución que ha logrado hasta ahora. El optimizador de enjambre de partículas rastrea el mejor valor local obtenido hasta ahora por cualquier partícula en el vecindario local. Las partículas restantes luego se mueven a través del espacio del problema siguiendo el ejemplo de las partículas óptimas. En cada iteración de tiempo, el optimizador de enjambre de partículas acelera cada partícula hacia sus ubicaciones óptimas de acuerdo con reglas matemáticas simples . La optimización de enjambre de partículas se ha aplicado en muchas áreas. Tiene pocos parámetros para ajustar, y una versión que funciona bien para aplicaciones específicas también puede funcionar bien con modificaciones menores en una variedad de aplicaciones relacionadas. [44] Un libro de Kennedy y Eberhart describe algunos aspectos filosóficos de las aplicaciones de optimización de enjambre de partículas y la inteligencia de enjambre. [45] Poli realiza un estudio extenso de las aplicaciones. [46] [47]
En Suiza, unos investigadores han desarrollado un algoritmo basado en la regla de Hamilton para la selección de parentesco. El algoritmo muestra cómo el altruismo en un enjambre de entidades puede, con el tiempo, evolucionar y dar lugar a un comportamiento más eficaz del enjambre. [48] [49]

Las primeras evidencias de comportamiento en grupo en los animales datan de hace unos 480 millones de años. Recientemente se ha descrito a fósiles del trilobite Ampyx priscus agrupados en líneas a lo largo del fondo del océano. Todos los animales eran adultos maduros y estaban todos orientados en la misma dirección como si hubieran formado una conga o un pelotón . Se ha sugerido que se alinean de esta manera para migrar, de forma muy similar a como las langostas espinosas migran en filas de una sola fila; [50] también se ha sugerido que la formación es el precursor del apareamiento, [51] como ocurre con la mosca Leptoconops torrens . Los hallazgos sugieren que el comportamiento colectivo animal tiene orígenes evolutivos muy tempranos. [52]
Se encuentran ejemplos de enjambre biológico en bandadas de aves , [53] bancos de peces , [54] [55] enjambres de insectos , [56] enjambres de bacterias , [57] [58] mohos, [59] motores moleculares , [60] manadas de cuadrúpedos , [61] y personas. [62] [63] [64] [65]
El comportamiento de los insectos sociales (insectos que viven en colonias , como las hormigas, las abejas, las avispas y las termitas) siempre ha sido una fuente de fascinación para niños, naturalistas y artistas. Los insectos individuales parecen hacer lo suyo sin ningún control central, pero la colonia en su conjunto se comporta de una manera altamente coordinada. [66] Los investigadores han descubierto que la cooperación a nivel de colonia es en gran medida autoorganizada . La coordinación grupal que surge es a menudo solo una consecuencia de la forma en que interactúan los individuos de la colonia. Estas interacciones pueden ser notablemente simples, como una hormiga que simplemente sigue el rastro dejado por otra hormiga. Sin embargo, en conjunto, el efecto acumulativo de tales comportamientos puede resolver problemas altamente complejos, como localizar la ruta más corta en una red de posibles caminos hacia una fuente de alimento. El comportamiento organizado que surge de esta manera a veces se llama inteligencia de enjambre , una forma de emergencia biológica . [66]
Las hormigas individuales no exhiben comportamientos complejos, pero una colonia de hormigas colectivamente logra tareas complejas como construir nidos, cuidar a sus crías, construir puentes y buscar comida. Una colonia de hormigas puede seleccionar colectivamente (es decir, enviar a la mayoría de las obreras hacia) la mejor fuente de alimento, o la más cercana, de varias en las cercanías. [67] Tales decisiones colectivas se logran utilizando mecanismos de retroalimentación positiva. La selección de la mejor fuente de alimento se logra mediante hormigas siguiendo dos reglas simples. Primero, las hormigas que encuentran alimento regresan al nido depositando una sustancia química llamada feromona . Se deposita más feromona para las fuentes de alimento de mayor calidad. [68] Por lo tanto, si se encuentran simultáneamente dos fuentes de alimento equidistantes de diferentes calidades, el rastro de feromonas hacia la mejor será más fuerte. Las hormigas en el nido siguen otra regla simple, para favorecer los rastros más fuertes, en promedio. Luego, más hormigas siguen el rastro más fuerte, por lo que más hormigas llegan a la fuente de alimento de alta calidad y se asegura un ciclo de retroalimentación positiva, lo que resulta en una decisión colectiva para la mejor fuente de alimento. Si hay dos caminos desde el hormiguero hasta una fuente de alimento, la colonia suele elegir el más corto, ya que las hormigas que primero regresan al hormiguero desde la fuente de alimento tienen más probabilidades de ser las que tomaron el camino más corto. Luego, más hormigas vuelven a recorrer el camino más corto, lo que refuerza el rastro de feromonas. [69]
Las hormigas legionarias , a diferencia de la mayoría de las especies de hormigas, no construyen nidos permanentes; una colonia de hormigas legionarias se mueve casi incesantemente durante el tiempo que existe, permaneciendo en un estado de enjambre esencialmente perpetuo. Varios linajes han desarrollado de forma independiente el mismo síndrome conductual y ecológico básico, a menudo denominado "comportamiento legionario", y puede ser un ejemplo de evolución convergente . [70]
Las técnicas exitosas que utilizan las colonias de hormigas se han estudiado en informática y robótica para producir sistemas distribuidos y tolerantes a fallos para resolver problemas. Esta área de la biomimética ha dado lugar a estudios sobre la locomoción de las hormigas, los motores de búsqueda que utilizan "rutas de búsqueda de alimentos", el almacenamiento tolerante a fallos y los algoritmos de redes . [71]

En climas templados, las abejas melíferas suelen formar enjambres a finales de la primavera. Un enjambre suele contener aproximadamente la mitad de las obreras junto con la reina vieja, mientras que la nueva reina se queda con las obreras restantes en la colmena original. Cuando las abejas melíferas emergen de una colmena para formar un enjambre, pueden reunirse en una rama de un árbol o en un arbusto a solo unos metros de la colmena. Las abejas se agrupan alrededor de la reina y envían de 20 a 50 exploradoras para encontrar nuevas ubicaciones adecuadas para nidos. Las exploradoras son las recolectoras más experimentadas del grupo. Si una exploradora encuentra una ubicación adecuada, regresa al grupo y lo promueve bailando una versión de la danza del meneo . Esta danza transmite información sobre la calidad, la dirección y la distancia del nuevo sitio. Cuanto más emocionada esté con sus hallazgos, más vigorosamente baila. Si puede convencer a los demás, es posible que se vayan y revisen el sitio que encontró. Si lo aprueban, también pueden promocionarlo. En este proceso de toma de decisiones, las exploradoras revisan varios sitios, a menudo abandonando su propio sitio original para promover el sitio superior de otra exploradora. Al principio, diferentes exploradoras pueden promover varios sitios diferentes. Después de algunas horas y, a veces, días, finalmente surge una ubicación preferida de este proceso de toma de decisiones. Cuando todas las exploradoras están de acuerdo con la ubicación final, todo el grupo despega y se dirige a ella. A veces, si no se llega a una decisión, el enjambre se separará, algunas abejas irán en una dirección y otras, en otra. Esto generalmente resulta en un fracaso, ya que ambos grupos mueren. Una nueva ubicación suele estar a un kilómetro o más de la colmena original, aunque algunas especies, por ejemplo, Apis dorsata , [72] pueden establecer nuevas colonias a tan solo 500 metros del nido natal. Este proceso de toma de decisiones colectiva es notablemente exitoso en la identificación del nuevo sitio de nido más adecuado y en mantener intacto el enjambre. Un buen sitio para una colmena debe ser lo suficientemente grande para acomodar el enjambre (aproximadamente 15 litros de volumen), debe estar bien protegido de los elementos, recibir una cantidad óptima de luz solar, estar a cierta altura sobre el suelo, tener una entrada pequeña y ser capaz de resistir la infestación de hormigas; es por eso que a menudo se seleccionan cavidades en los árboles. [73] [74] [75] [76] [77]
A diferencia de los insectos sociales, los enjambres de insectos no sociales que se han estudiado parecen funcionar principalmente en contextos como el apareamiento, la alimentación, la evitación de depredadores y la migración.
Las polillas pueden exhibir un apareamiento sincronizado, durante el cual las feromonas liberadas por las hembras inician el comportamiento de búsqueda y enjambre en los machos. [78] Los machos detectan las feromonas con antenas sensibles y pueden rastrear a las hembras a varios kilómetros de distancia. [79] El apareamiento en enjambre implica la elección de la hembra y la competencia de los machos. Solo un macho en el enjambre, típicamente el primero, copulará con éxito. [80] Las hembras maximizan los beneficios de la aptitud y minimizan el costo al controlar el inicio y la magnitud de la feromona liberada. Muy poca feromona no atraerá a una pareja, demasiada permite que los machos menos aptos detecten la señal. [81] Después de la cópula, las hembras ponen los huevos en una planta huésped. La calidad de la planta huésped puede ser un factor que influya en la ubicación del enjambre y la puesta de huevos. En un caso, los investigadores observaron polillas de gusano de roble de rayas rosadas ( Anisota virginiensis ) enjambrando en un sitio de carroña , donde la descomposición probablemente aumentó los niveles de nutrientes del suelo y la calidad de la planta huésped. [82]
Los mosquitos, como Tokunagayusurika akamusi , forman enjambres que bailan en el aire. El enjambre tiene múltiples propósitos, incluido el de facilitar el apareamiento al atraer a las hembras para que se acerquen al enjambre, un fenómeno conocido como apareamiento lek . Estos enjambres en forma de nube a menudo se forman a primera hora de la tarde cuando el sol está bajando, en la punta de un arbusto, en la cima de una colina, sobre un estanque de agua o incluso a veces sobre una persona. La formación de estos enjambres no se debe al instinto, sino a un comportamiento adaptativo -un "consenso"- entre los individuos dentro de los enjambres. También se sugiere que el enjambre es un ritual , porque rara vez hay un mosquito macho solo y no en un enjambre. Esto podría haberse formado debido al beneficio de reducir la endogamia al tener machos de varios genes reunidos en un solo lugar. [83] El género Culicoides , también conocido como mosquitos mordedores, ha mostrado un comportamiento de enjambre que se cree que causa confusión en los depredadores. [84]
Las cucarachas dejan rastros químicos en sus heces y emiten feromonas en el aire para aparearse. Otras cucarachas seguirán estos rastros para descubrir fuentes de alimento y agua, y también descubrirán dónde se esconden otras cucarachas. Por lo tanto, los grupos de cucarachas pueden exhibir un comportamiento emergente [85] , en el que el comportamiento grupal o de enjambre surge de un conjunto simple de interacciones individuales.
Las cucarachas son animales principalmente nocturnos y huyen cuando se exponen a la luz. Un estudio puso a prueba la hipótesis de que las cucarachas utilizan sólo dos datos para decidir a dónde ir en esas condiciones: lo oscuro que está y cuántas otras cucarachas hay. El estudio, realizado por José Halloy y sus colegas de la Universidad Libre de Bruselas y otras instituciones europeas, creó un conjunto de pequeños robots que se les aparecen a las cucarachas como otras cucarachas y que pueden así alterar la percepción de masa crítica de las cucarachas . Los robots también estaban perfumados especialmente para que fueran aceptados por las cucarachas reales. [86]

Las langostas son la fase de enjambre de los saltamontes de cuernos cortos de la familia Acrididae . Algunas especies pueden reproducirse rápidamente en condiciones adecuadas y posteriormente volverse gregarias y migratorias. Forman bandas como ninfas y enjambres como adultos, los cuales pueden viajar grandes distancias, despojando rápidamente los campos y dañando enormemente los cultivos . Los enjambres más grandes pueden cubrir cientos de millas cuadradas y contener miles de millones de langostas. Una langosta puede comer su propio peso (alrededor de 2 gramos) en plantas todos los días. Eso significa que un millón de langostas pueden comer más de una tonelada de alimento cada día, y los enjambres más grandes pueden consumir más de 100.000 toneladas cada día. [87]
Se ha descubierto que la enjambre de langostas está asociada con mayores niveles de serotonina , lo que hace que la langosta cambie de color, coma mucho más, se atraiga mutuamente y se reproduzca con mucha más facilidad. Los investigadores proponen que el comportamiento de enjambre es una respuesta al hacinamiento y los estudios han demostrado que el aumento de la estimulación táctil de las patas traseras o, en algunas especies, simplemente el encuentro con otros individuos provoca un aumento en los niveles de serotonina. La transformación de la langosta a la variedad de enjambre puede ser inducida por varios contactos por minuto durante un período de cuatro horas. [88] [89] [90] [91] Cabe destacar que se ha encontrado una predisposición innata a agruparse en las crías de la langosta del desierto, Schistocerca gregaria , independientemente de su fase parental. [92]
La respuesta de una langosta individual a una pérdida de alineación en el grupo parece aumentar la aleatoriedad de su movimiento, hasta que se alcanza nuevamente un estado de alineación. Esta alineación inducida por el ruido parece ser una característica intrínseca del movimiento coherente colectivo. [93]

La migración de insectos es el movimiento estacional de los insectos , en particular de las especies de libélulas , escarabajos , mariposas y polillas . La distancia puede variar de una especie a otra, pero en la mayoría de los casos estos movimientos involucran un gran número de individuos. En algunos casos, los individuos que migran en una dirección pueden no regresar y la siguiente generación puede migrar en la dirección opuesta. Esta es una diferencia significativa con la migración de las aves .
Las mariposas monarca son especialmente conocidas por su larga migración anual. En América del Norte realizan migraciones masivas hacia el sur a partir de agosto hasta la primera helada. Una migración hacia el norte tiene lugar en la primavera. La monarca es la única mariposa que migra tanto al norte como al sur como lo hacen las aves de forma regular. Pero ningún individuo hace el viaje de ida y vuelta completo. Las monarcas hembras depositan huevos para la siguiente generación durante estas migraciones. [94] La duración de estos viajes excede la vida normal de la mayoría de las monarcas, que es de menos de dos meses para las mariposas nacidas a principios del verano. La última generación del verano entra en una fase no reproductiva conocida como diapausa y puede vivir siete meses o más. [95] Durante la diapausa, las mariposas vuelan a uno de los muchos sitios de hibernación. La generación que hiberna generalmente no se reproduce hasta que abandona el sitio de hibernación en algún momento de febrero y marzo. Son la segunda, tercera y cuarta generaciones las que regresan a sus ubicaciones del norte en los Estados Unidos y Canadá en la primavera. Cómo la especie logra regresar a los mismos lugares de hibernación después de un intervalo de varias generaciones es todavía un tema de investigación; los patrones de vuelo parecen ser heredados, basados en una combinación de la posición del sol en el cielo [96] y una brújula solar compensada en el tiempo que depende de un reloj circadiano ubicado en sus antenas. [97] [98]

 Murmullos de estorninos
Murmullos de estorninos[99]

Aproximadamente 1.800 de las 10.000 especies de aves del mundo migran a largas distancias. [100] La principal motivación para la migración parece ser la alimentación; por ejemplo, algunos colibríes optan por no migrar si se alimentan durante el invierno. Además, los días más largos del verano boreal proporcionan más tiempo para que las aves reproductoras alimenten a sus crías. Esto ayuda a las aves diurnas a producir nidadas más grandes que las especies no migratorias relacionadas que permanecen en los trópicos. A medida que los días se acortan en otoño, las aves regresan a regiones más cálidas donde el suministro de alimentos disponible varía poco con la estación. Estas ventajas compensan el alto estrés, los costos del esfuerzo físico y otros riesgos de la migración, como la depredación.
Muchas aves migran en bandadas. En el caso de las aves más grandes, se supone que volar en bandadas reduce los costes energéticos. A menudo se supone que la formación en V aumenta la eficiencia y el alcance de las aves en vuelo, en particular en rutas migratorias largas . Todas las aves, excepto la primera, vuelan en la corriente ascendente de uno de los vórtices de la punta del ala del ave que va delante. La corriente ascendente ayuda a cada ave a soportar su propio peso en vuelo, de la misma forma que un planeador puede ascender o mantener la altura indefinidamente en el aire ascendente. Los gansos que vuelan en formación en V ahorran energía al volar en la corriente ascendente del vórtice de la punta del ala generado por el animal anterior en la formación. Por tanto, las aves que vuelan detrás no necesitan esforzarse tanto para lograr la elevación. Los estudios muestran que las aves en formación en V se colocan aproximadamente a la distancia óptima predicha por la teoría aerodinámica simple. [101] Los gansos en formación en V pueden conservar entre el 12 y el 20 % de la energía que necesitarían para volar solos. [102] [103] En estudios de radar se ha descubierto que los playeros rojizos y los correlimos vuelan 5 km por hora más rápido en bandadas que cuando vuelan solos. [104] Las aves que vuelan en las puntas y en la parte delantera se rotan de manera cíclica y oportuna para distribuir la fatiga del vuelo de manera uniforme entre los miembros de la bandada. La formación también facilita la comunicación y permite que las aves mantengan contacto visual entre sí.
Otros animales pueden utilizar técnicas de desplazamiento similares durante la migración. Las langostas , por ejemplo, migran en formación cerrada en "trenes de langostas", a veces durante cientos de millas.
El Mediterráneo y otros mares presentan un gran obstáculo para las aves planeadoras, que deben cruzar por los puntos más estrechos. Cantidades masivas de grandes rapaces y cigüeñas pasan por áreas como Gibraltar , Falsterbo y el Bósforo en épocas de migración. Especies más comunes, como el busardo abejero europeo , pueden contarse en cientos de miles en otoño. Otras barreras, como las cadenas montañosas, también pueden causar embudos, particularmente de grandes migrantes diurnos. Este es un factor notable en el cuello de botella migratorio centroamericano . Esta concentración de aves durante la migración puede poner en peligro a las especies. Algunas aves migratorias espectaculares ya se han extinguido, la más notable es la paloma migratoria . Durante la migración, las bandadas tenían una milla (1,6 km) de ancho y 300 millas (500 km) de largo, tardaban varios días en pasar y contenían hasta mil millones de aves.

El término "banco" se puede utilizar para describir cualquier grupo de peces, incluidos los grupos de especies mixtas, mientras que "escuela" se utiliza para grupos más unidos de la misma especie que nadan de manera altamente sincronizada y polarizada.
Los peces obtienen muchos beneficios de la pertenencia a cardúmenes, incluida la defensa contra los depredadores (mediante una mejor detección de los depredadores y la dilución de las posibilidades de captura), un mayor éxito en la búsqueda de alimento y un mayor éxito en la búsqueda de pareja. [106] También es probable que los peces se beneficien de la pertenencia a cardúmenes mediante una mayor eficiencia hidrodinámica . [107]
Los peces utilizan muchos rasgos para elegir a sus compañeros de cardumen. Generalmente prefieren cardúmenes más grandes, compañeros de su misma especie, compañeros de cardumen similares en tamaño y apariencia a ellos, peces sanos y parientes (cuando los reconocen). El "efecto de rareza" postula que cualquier miembro del cardumen que se destaque en apariencia será el objetivo preferencial de los depredadores. Esto puede explicar por qué los peces prefieren cardúmenes con individuos que se les parecen. El efecto de rareza tendería, por lo tanto, a homogeneizar los cardúmenes. [108]
Un aspecto desconcertante de la selección de cardúmenes es cómo un pez puede elegir unirse a un cardumen de animales similares a él, dado que no puede conocer su propia apariencia. Los experimentos con peces cebra han demostrado que la preferencia por cardúmenes es una habilidad aprendida, no innata. Un pez cebra tiende a asociarse con cardúmenes que se parecen a los cardúmenes en los que se crió, una forma de impronta . [109]
Otras cuestiones abiertas sobre el comportamiento de los cardúmenes incluyen la identificación de los individuos responsables de la dirección del movimiento de los cardúmenes. En el caso del movimiento migratorio , la mayoría de los miembros de un cardumen parecen saber hacia dónde se dirigen. En el caso del comportamiento de búsqueda de alimento, los cardúmenes cautivos de alburnos dorados (una especie de pececillo ) están liderados por un pequeño número de individuos experimentados que saben cuándo y dónde hay comida disponible. [110]
Radakov estimó que los bancos de arenque en el Atlántico Norte pueden ocupar hasta 4,8 kilómetros cúbicos (1,2 millas cúbicas) con densidades de peces de entre 0,5 y 1,0 pez/metro cúbico, lo que suma un total de varios miles de millones de peces en un banco. [111]
Entre mayo y julio, una gran cantidad de sardinas desovan en las frías aguas del Banco de las Agujas y luego siguen una corriente de agua fría hacia el norte a lo largo de la costa este de Sudáfrica. Esta gran migración, llamada la migración de las sardinas , genera espectaculares frenesíes alimentarios a lo largo de la costa, ya que los depredadores marinos, como delfines, tiburones y alcatraces, atacan los bancos.
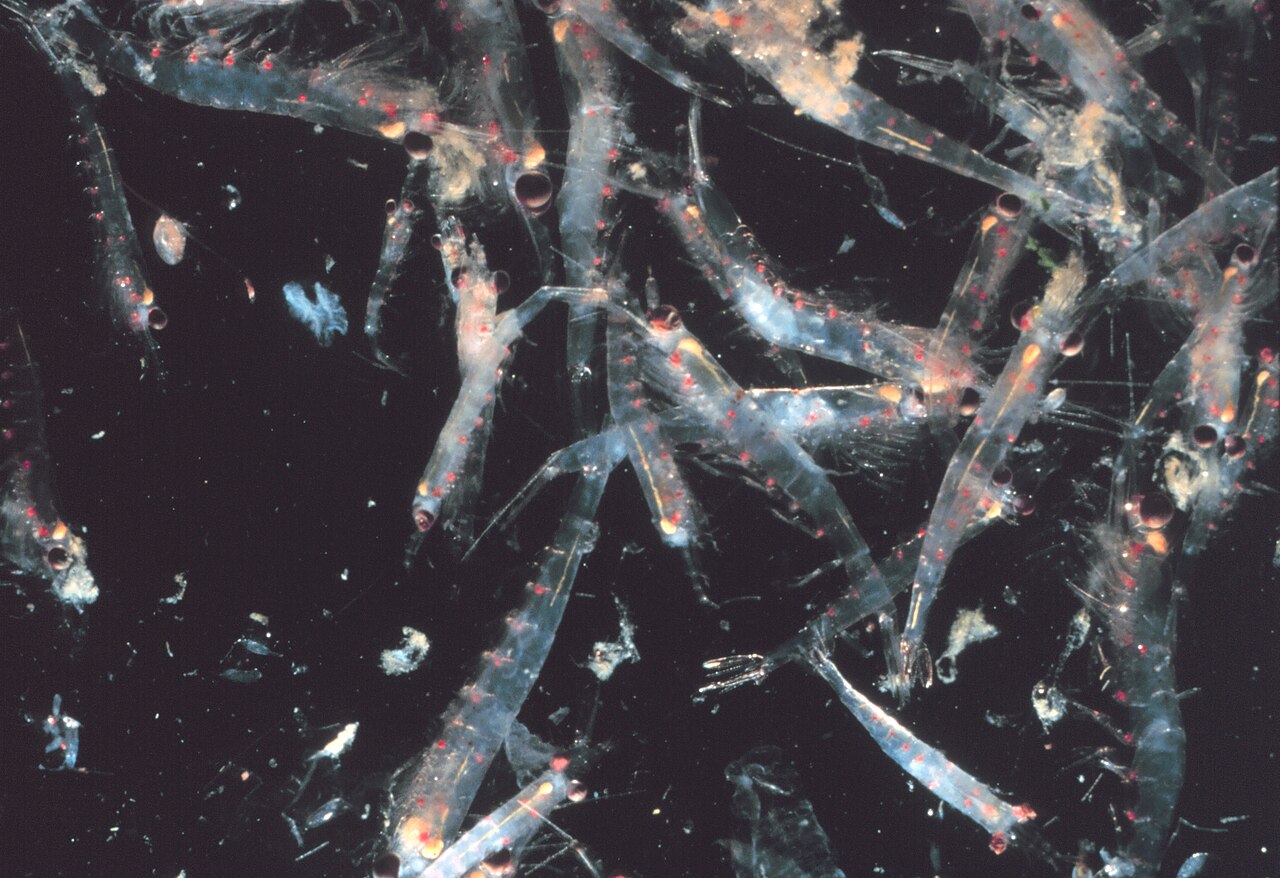
La mayoría de los krill , pequeños crustáceos similares a los camarones , forman grandes enjambres, que a veces alcanzan densidades de 10.000 a 60.000 animales individuales por metro cúbico. [113] [114] [115] La formación de enjambres es un mecanismo defensivo que confunde a los depredadores más pequeños que querrían localizar individuos individuales. Los enjambres más grandes son visibles desde el espacio y se pueden rastrear por satélite. [116] Se observó que un enjambre cubría un área de 450 kilómetros cuadrados (175 millas cuadradas) de océano, hasta una profundidad de 200 metros (650 pies) y se estimó que contenía más de 2 millones de toneladas de krill. [117] Investigaciones recientes sugieren que el krill no simplemente se desplaza pasivamente en estas corrientes, sino que en realidad las modifica. [117] El krill normalmente sigue una migración vertical diurna . Al desplazarse verticalmente por el océano en ciclos de 12 horas, los enjambres desempeñan un papel importante en la mezcla de aguas más profundas, ricas en nutrientes, con aguas pobres en nutrientes en la superficie. [117] Hasta hace poco se suponía que pasaban el día a mayores profundidades y subían durante la noche hacia la superficie. Se ha descubierto que cuanto más profundo descienden, más reducen su actividad, [118] aparentemente para reducir los encuentros con depredadores y conservar energía.
Trabajos posteriores sugirieron que la actividad de natación del krill variaba con la saciedad del estómago. Los animales saciados que habían estado alimentándose en la superficie nadaban menos activamente y, por lo tanto, se hundían debajo de la capa mixta. [119] A medida que se hunden, producen heces, lo que puede significar que tienen un papel importante que desempeñar en el ciclo del carbono antártico. Se descubrió que el krill con el estómago vacío nadaba más activamente y, por lo tanto, se dirigía hacia la superficie. Esto implica que la migración vertical puede ocurrir dos o tres veces al día. Algunas especies forman enjambres superficiales durante el día con fines de alimentación y reproducción, aunque este comportamiento es peligroso porque los hace extremadamente vulnerables a los depredadores. [120] Los enjambres densos pueden provocar un frenesí alimentario entre los peces, las aves y los mamíferos depredadores, especialmente cerca de la superficie. Cuando se les molesta, un enjambre se dispersa, e incluso se ha observado que algunos individuos mudan instantáneamente, dejando atrás la exuvia como señuelo. [121] En 2012, Gandomi y Alavi presentaron lo que parece ser un algoritmo estocástico exitoso para modelar el comportamiento de los bancos de krill. El algoritmo se basa en tres factores principales: " (i) movimiento inducido por la presencia de otros individuos (ii) actividad de alimentación, y (iii) difusión aleatoria". [122]

Los copépodos son un grupo de diminutos crustáceos que se encuentran en el mar y los lagos. Muchas especies son planctónicas (flotan en las aguas marinas) y otras son bentónicas (viven en el fondo del océano). Los copépodos suelen tener entre 1 y 2 milímetros (0,04 y 0,08 pulgadas) de largo, un cuerpo en forma de lágrima y grandes antenas . Aunque, al igual que otros crustáceos, tienen un exoesqueleto acorazado , son tan pequeños que en la mayoría de las especies esta fina armadura, y todo el cuerpo, es casi totalmente transparente. Los copépodos tienen un único ojo compuesto, mediano, generalmente de color rojo brillante, en el centro de la cabeza transparente.
Los copépodos también forman enjambres. Por ejemplo, se han observado regularmente enjambres monoespecíficos alrededor de los arrecifes de coral y las praderas marinas , y en lagos. Las densidades de enjambres eran de alrededor de un millón de copépodos por metro cúbico. Los enjambres típicos tenían uno o dos metros de diámetro, pero algunos superaban los 30 metros cúbicos. Los copépodos necesitan contacto visual para mantenerse juntos y se dispersan por la noche. [123]
La primavera produce floraciones de fitoplancton enjambre que proporcionan alimento a los copépodos. Los copépodos planctónicos suelen ser los miembros dominantes del zooplancton y, a su vez, son los principales organismos alimentarios de muchos otros animales marinos. En particular, los copépodos son presa de peces forrajeros y medusas , que pueden reunirse en enormes enjambres de millones de ejemplares. Algunos copépodos tienen respuestas de escape extremadamente rápidas cuando detectan un depredador y pueden saltar a gran velocidad unos pocos milímetros (ver la imagen animada a continuación).

Los copépodos planctónicos son importantes para el ciclo del carbono . Algunos científicos dicen que forman la biomasa animal más grande de la Tierra. [124] Compiten por este título con el krill antártico . Sin embargo, debido a su menor tamaño y tasas de crecimiento relativamente más rápidas, y debido a que están distribuidos de manera más uniforme en la mayor parte de los océanos del mundo, los copépodos casi con certeza contribuyen mucho más a la productividad secundaria de los océanos del mundo y al sumidero global de carbono oceánico que el krill , y quizás más que todos los demás grupos de organismos juntos. Actualmente se cree que las capas superficiales de los océanos son el sumidero de carbono más grande del mundo, absorbiendo alrededor de 2 mil millones de toneladas de carbono al año, el equivalente a quizás un tercio de las emisiones de carbono humanas , lo que reduce su impacto. Muchos copépodos planctónicos se alimentan cerca de la superficie por la noche, luego se hunden en aguas más profundas durante el día para evitar depredadores visuales. Sus exoesqueletos mudados, bolitas fecales y respiración en profundidad llevan carbono a las profundidades marinas.
En los océanos y lagos viven muchos organismos unicelulares llamados fitoplancton . Cuando se dan ciertas condiciones, como niveles elevados de nutrientes o de luz, estos organismos se reproducen de forma explosiva. El enjambre denso de fitoplancton resultante se denomina floración de algas . Las floraciones pueden cubrir cientos de kilómetros cuadrados y se ven fácilmente en imágenes satelitales. El fitoplancton individual rara vez vive más de unos pocos días, pero las floraciones pueden durar semanas. [125] [126]
Los científicos han atribuido el comportamiento de enjambre a las plantas durante cientos de años. En su libro de 1800, Phytologia: or, The philosophy of farming and gardening (Phytología: o la filosofía de la agricultura y la jardinería) , Erasmus Darwin escribió que el crecimiento de las plantas se parecía a los enjambres observados en otras partes de la naturaleza. [127] Si bien se refería a observaciones más amplias de la morfología de las plantas y se centraba tanto en el comportamiento de las raíces como de los brotes, investigaciones recientes han respaldado esta afirmación.
Las raíces de las plantas , en particular, muestran un comportamiento de enjambre observable, creciendo en patrones que exceden el umbral estadístico de probabilidad aleatoria e indican la presencia de comunicación entre los ápices de las raíces individuales . La función principal de las raíces de las plantas es la absorción de nutrientes del suelo , y es este propósito el que impulsa el comportamiento de enjambre. Las plantas que crecen en estrecha proximidad han adaptado su crecimiento para asegurar la disponibilidad óptima de nutrientes. Esto se logra creciendo en una dirección que optimiza la distancia entre las raíces cercanas, aumentando así sus posibilidades de explotar las reservas de nutrientes sin explotar. La acción de este comportamiento toma dos formas: maximización de la distancia desde los ápices de las raíces vecinas y repulsión por ellos. [128] La zona de transición de la punta de una raíz es en gran medida responsable de monitorear la presencia de hormonas transmitidas por el suelo, señalando patrones de crecimiento de respuesta según corresponda. Las respuestas de las plantas a menudo son complejas, integrando múltiples entradas para informar una respuesta autónoma. Las entradas adicionales que informan el crecimiento del enjambre incluyen la luz y la gravedad, las cuales también se monitorean en la zona de transición del ápice de una raíz. [129] Estas fuerzas actúan para informar a cualquier número de raíces "principales" en crecimiento, que exhiben sus propias liberaciones independientes de sustancias químicas inhibidoras para establecer el espaciamiento apropiado, contribuyendo así a un patrón de comportamiento de enjambre. El crecimiento horizontal de las raíces, ya sea en respuesta al alto contenido mineral en el suelo o debido al crecimiento de estolones , produce un crecimiento ramificado que se establece para formar también sus propios enjambres de raíces independientes. [130]
El enjambre también describe las agrupaciones de algunos tipos de bacterias depredadoras , como las mixobacterias . Las mixobacterias se agrupan en "manadas de lobos", moviéndose activamente mediante un proceso conocido como deslizamiento bacteriano y manteniéndose juntas con la ayuda de señales moleculares intercelulares . [57] [131]


Un grupo de personas también puede mostrar un comportamiento de enjambre, como los peatones [134] o los soldados que se agolpan en los parapetos [ dudoso – discutir ] . En Colonia, Alemania, dos biólogos de la Universidad de Leeds demostraron un comportamiento de bandada en humanos. El grupo de personas exhibió un patrón de comportamiento similar al de una bandada, donde si el cinco por ciento de la bandada cambiaba de dirección, los demás la seguían. Si una persona era designada como depredador y todos los demás debían evitarla, la bandada se comportaba de manera muy similar a un banco de peces. [135] [136] Comprender cómo interactúan los humanos en multitudes es importante si se pretende que la gestión de multitudes evite de manera efectiva las bajas en los campos de fútbol, los conciertos de música y las estaciones de metro. [137]
El modelado matemático del comportamiento de las multitudes es una tecnología común y ha encontrado usos en la animación. Las simulaciones de multitudes se han utilizado en muchas películas [138] para generar multitudes que se mueven de manera realista. Batman Returns de Tim Burton fue la primera película en hacer uso de la tecnología de enjambre para la representación, representando de manera realista los movimientos de un grupo de murciélagos utilizando el sistema boids . La trilogía cinematográfica de El Señor de los Anillos hizo uso de una tecnología similar, conocida como Massive , durante las escenas de batalla. La tecnología de enjambre es particularmente atractiva porque es barata, robusta y simple.
También se ha utilizado una simulación informática basada en hormigas que utiliza sólo seis reglas de interacción para evaluar el comportamiento de embarque en los aviones. [139] Las aerolíneas también han utilizado el enrutamiento basado en hormigas para asignar las llegadas de los aviones a las puertas del aeropuerto. Un sistema de aerolíneas desarrollado por Douglas A. Lawson utiliza la teoría de enjambre, o inteligencia de enjambre , la idea de que una colonia de hormigas funciona mejor que una sola. Cada piloto actúa como una hormiga que busca la mejor puerta del aeropuerto. "El piloto aprende de su experiencia qué es lo mejor para él, y resulta que esa es la mejor solución para la aerolínea", explica Lawson. Como resultado, la "colonia" de pilotos siempre va a las puertas en las que puede llegar y salir rápidamente. El programa puede incluso avisar a un piloto de los atascos de aviones antes de que se produzcan. "Podemos anticipar que va a ocurrir, por lo que tendremos una puerta disponible", dice Lawson. [140]
El comportamiento de enjambre también se produce en la dinámica del flujo de tráfico , como en la ola de tráfico . El tráfico bidireccional se puede observar en los rastros de hormigas . [141] [142] En los últimos años, este comportamiento se ha investigado para obtener información sobre los modelos de peatones y tráfico. [143] [144] Las simulaciones basadas en modelos de peatones también se han aplicado a multitudes que se precipitan en estampida debido al pánico. [145]
El comportamiento gregario en el marketing se ha utilizado para explicar las dependencias del comportamiento mutuo de los clientes. The Economist informó sobre una conferencia reciente en Roma sobre el tema de la simulación del comportamiento humano adaptativo. [146] En ella se compartieron mecanismos para aumentar las compras impulsivas y hacer que la gente "compre más jugando con el instinto gregario". La idea básica es que la gente comprará más productos que se consideran populares, y se mencionan varios mecanismos de retroalimentación para hacer llegar a los consumidores información sobre la popularidad de los productos, incluida la tecnología de tarjetas inteligentes y el uso de la tecnología de etiquetas de identificación por radiofrecuencia . Un investigador del Instituto Tecnológico de Florida presentó un modelo de "movimientos de enjambre" , que resulta atractivo para los supermercados porque puede "aumentar las ventas sin necesidad de ofrecer descuentos a la gente".

La aplicación de los principios de enjambre a los robots se denomina robótica de enjambre , mientras que la inteligencia de enjambre se refiere al conjunto más general de algoritmos.
Parcialmente inspirados por colonias de insectos como hormigas y abejas, los investigadores están modelando el comportamiento de enjambres de miles de pequeños robots que juntos realizan una tarea útil, como encontrar algo escondido, limpiar o espiar. Cada robot es bastante simple, pero el comportamiento emergente del enjambre es más complejo. [1] El conjunto completo de robots puede considerarse como un único sistema distribuido, de la misma manera que una colonia de hormigas puede considerarse un superorganismo , que exhibe inteligencia de enjambre . El enjambre más grande creado hasta ahora es el enjambre de 1024 robots Kilobot. [148] Otros enjambres grandes incluyen el enjambre iRobot , el proyecto SRI International / ActivMedia Robotics Centibots , [149] y el enjambre Open-source Micro-robotic Project, que se están utilizando para investigar comportamientos colectivos. [150] [151] Los enjambres también son más resistentes al fracaso. Mientras que un robot grande puede fallar y arruinar una misión, un enjambre puede continuar incluso si fallan varios robots. Esto podría hacerlos atractivos para misiones de exploración espacial, donde el fracaso normalmente es extremadamente costoso. [152] Además de los vehículos terrestres, la robótica de enjambre también incluye la investigación de enjambres de robots aéreos [147] [153] y equipos heterogéneos de vehículos terrestres y aéreos. [154] [155]
A diferencia de los robots macroscópicos, las partículas coloidales a microescala también pueden adoptarse como agentes para realizar comportamientos colectivos para llevar a cabo tareas utilizando enfoques mecánicos y físicos, como un microenjambre reconfigurable tipo tornado [156] que imita a los peces en cardúmenes, [157] especies de partículas jerárquicas [158] que imitan el comportamiento depredador de los mamíferos, y la manipulación de microobjetos utilizando un microenjambre transformable. [159] La fabricación de dichas partículas coloidales generalmente se basa en la síntesis química.

El enjambre militar es un comportamiento en el que unidades de acción autónomas o parcialmente autónomas atacan a un enemigo desde varias direcciones diferentes y luego se reagrupan. El pulso , donde las unidades cambian el punto de ataque, también es una parte del enjambre militar. El enjambre militar implica el uso de una fuerza descentralizada contra un oponente, de una manera que enfatiza la movilidad, la comunicación, la autonomía de la unidad y la coordinación o sincronización. [160] Históricamente, las fuerzas militares usaban principios de enjambre sin realmente examinarlos explícitamente, pero ahora la investigación activa examina conscientemente las doctrinas militares que extraen ideas del enjambre.
El mero hecho de que varias unidades converjan sobre un objetivo no implica necesariamente que se trate de enjambres. Las operaciones de asedio no implican enjambres, porque no hay maniobras; hay convergencia, pero sobre la fortificación sitiada. Las emboscadas de guerrillas tampoco constituyen enjambres, porque son de tipo "golpe y fuga". Aunque la emboscada puede tener varios puntos de ataque sobre el enemigo, las guerrillas se retiran cuando han infligido el daño adecuado o cuando se encuentran en peligro.
En 2014, la Oficina de Investigación Naval de Estados Unidos publicó un vídeo que mostraba pruebas de un enjambre de pequeñas embarcaciones de ataque con drones autónomos que pueden dirigirse y tomar acciones ofensivas coordinadas como grupo. [161]

![Las personas que se agolpan en una salida no siempre se comportan como un fluido.[163][164]](http://upload.wikimedia.org/wikipedia/commons/thumb/5/54/Crowd_04378.JPG/1280px-Crowd_04378.JPG)


{{cite book}}: CS1 maint: varios nombres: lista de autores ( enlace ){{cite web}}: CS1 maint: URL no apta ( enlace )


![Las salpas dispuestas en cadenas forman enormes enjambres.[162]](http://upload.wikimedia.org/wikipedia/commons/thumb/6/63/Salp.jpg/1280px-Salp.jpg)

