Stegoceras es un género de dinosaurio paquicefalosáurido (con cabeza en forma de cúpula)que vivió en lo que hoy es América del Norte durante el período Cretácico Superior , hace unos 77,5 a 74 millones de años (mya). Los primeros especímenes de Alberta , Canadá, fueron descritos en 1902, y la especie tipo Stegoceras validum se basó en estos restos. El nombre genérico significa "techo de cuerno", y el nombre específico significa "fuerte". Varias otras especies se han incluido en el género a lo largo de los años, pero desde entonces se han movido a otros géneros o se han considerado sinónimos menores . Actualmente solo quedan S. validum y S. novomexicanum , nombrados en 2011 a partir de fósiles encontrados en Nuevo México . La validez de esta última especie también ha sido debatida, y es posible que ni siquiera pertenezca al género Stegoceras.
Stegoceras era un dinosaurio bípedo pequeño de entre 2 y 2,5 metros de largo y que pesaba entre 10 y 40 kilogramos. El cráneo era aproximadamente triangular con un hocico corto y tenía una cúpula gruesa, ancha y relativamente lisa en la parte superior. La parte posterior del cráneo tenía una "plataforma" gruesa sobre el occipucio y una cresta gruesa sobre los ojos. Gran parte del cráneo estaba adornado por tubérculos (o "excrecencias" redondas) y nódulos (o "protuberancias"), muchos en filas, y los más grandes formaban pequeños cuernos en la plataforma. Los dientes eran pequeños y aserrados. Se cree que el cráneo era plano en los animales juveniles y que se convirtió en una cúpula con la edad. Tenía una columna vertebral rígida y una cola rígida. La región pélvica era amplia, tal vez debido a un intestino extendido.
Stegoceras , que originalmente solo se conocía por las cúpulas craneales, fue uno de los primeros paquicefalosaurios conocidos, y la incompletitud de estos restos iniciales dio lugar a muchas teorías sobre las afinidades de este grupo. En 1924 se describió un cráneo completo de Stegoceras con partes asociadas del esqueleto, lo que arrojó más luz sobre estos animales. Hoy en día, los paquicefalosaurios se agrupan con los ceratopsianos con cuernos en el grupo Marginocephalia . El propio Stegoceras ha sido considerado basal (o "primitivo") en comparación con otros paquicefalosaurios. Lo más probable es que Stegoceras fuera herbívoro y probablemente tuviera un buen sentido del olfato. La función de la cúpula ha sido debatida y las teorías en competencia incluyen el uso en el combate intraespecífico (cabezazos o golpes en los flancos), la exhibición sexual o el reconocimiento de especies . S. validum se conoce de la Formación Dinosaur Park y la Formación Oldman , mientras que el controvertido S. novomexicanum se conoce de la Formación Fruitland y Kirtland .

Los primeros restos conocidos de Stegoceras fueron recolectados por el paleontólogo canadiense Lawrence Lambe del Belly River Group , en el distrito del río Red Deer de Alberta, Canadá . Estos restos consistían en dos cúpulas craneales parciales (ejemplares CMN 515 y CMN 1423 en el Museo Canadiense de la Naturaleza ) de dos animales de diferentes tamaños recolectados en 1898, y una tercera cúpula parcial (CMN 1594) recolectada en 1901. Basándose en estos especímenes, Lambe describió y nombró el nuevo género y especie monotípicos Stegoceras validus en 1902. [1] [ 2 ] El nombre genérico Stegoceras proviene del griego stegè /στέγη, que significa "techo" y keras /κέρας que significa "cuerno". El nombre específico validus significa "fuerte" en latín, posiblemente en referencia al grueso techo del cráneo. [3] Debido a que la especie se basó en múltiples especímenes (una serie sintipo ), CMN 515 fue designado como el espécimen lectotipo por John Bell Hatcher en 1907. [4] [2]
Como no se habían encontrado restos similares en la zona antes, Lambe no estaba seguro de qué tipo de dinosaurio eran y si representaban una especie o más; sugirió que las cúpulas eran "prenasales" situadas antes de los huesos nasales en la línea media de la cabeza, y notó su similitud con el núcleo del cuerno nasal de un espécimen de Triceratops . [1] En 1903, el paleontólogo húngaro Franz Nopcsa von Felső-Szilvás sugirió que las cúpulas fragmentarias de Stegoceras eran de hecho huesos frontales y nasales, y que el animal, por lo tanto, habría tenido un solo cuerno no apareado. Lambe simpatizaba con esta idea de un nuevo tipo de "dinosaurio unicornio" en una revisión de 1903 del artículo de Nopscsa. En ese momento, todavía había incertidumbre sobre a qué grupo de dinosaurios pertenecía Stegoceras , con tanto los ceratopsianos (dinosaurios con cuernos ) como contendientes. [5] [6] Hatcher dudaba de que los especímenes de Stegoceras pertenecieran a la misma especie y de que fueran dinosaurios en absoluto, y sugirió que las cúpulas consistían en los huesos frontal, occipital y parietal del cráneo. [2] En 1918, Lambe atribuyó otra cúpula (CMN 138) a S. validus y nombró una nueva especie, S. brevis , basándose en el espécimen CMN 1423 (que originalmente incluyó en S. validus ). En ese momento, consideró a estos animales como miembros de Stegosauria (entonces compuesta por ambas familias de dinosaurios acorazados, Stegosauridae y Ankylosauridae ), en una nueva familia que llamó Psalisauridae (nombrada por el techo abovedado o en forma de cúpula del cráneo). [7]
En 1924, el paleontólogo estadounidense Charles W. Gilmore describió un cráneo completo de S. validus con restos postcraneales asociados, por entonces los restos más completos de un dinosaurio con cabeza en forma de cúpula. Fue descubierto en el Belly River Group por el paleontólogo estadounidense George F. Sternberg en 1926, y catalogado como espécimen UALVP 2 en el Laboratorio de Paleontología de Vertebrados de la Universidad de Alberta . Este hallazgo confirmó la interpretación de Hatcher de que las cúpulas consistían en el área frontoparietal del cráneo. UALVP 2 fue encontrado con pequeños elementos óseos desarticulados, que entonces se pensaba que eran gastralia (costillas abdominales), que no se conocen en otros dinosaurios ornitisquios (uno de los dos grupos principales de dinosaurios). Gilmore señaló que los dientes de S. validus eran muy similares a los de la especie Troodon formosus (nombrada en 1856 y para entonces conocida solo a partir de dientes aislados), y describió una cúpula craneal descubierta cerca de la localidad donde se encontró Troodon . Por lo tanto, Gilmore consideró a Stegoceras un sinónimo menor inválido de Troodon , por lo que renombró a S. validus como T. validus , y sugirió que incluso las dos especies podrían ser la misma. Además, encontró que S. brevis era idéntico a S. validus , y por lo tanto un sinónimo menor de este último. También colocó estas especies en la nueva familia Troodontidae (ya que Lambe no había seleccionado un género tipo para su Psalisauridae), que consideró más cercana a los dinosaurios ornitópodos . [8] [9] Debido a que el cráneo parecía tan especializado en comparación con el esqueleto de aspecto más bien "primitivo", Nopcsa dudó de si estas partes realmente pertenecían juntas, y sugirió que el cráneo pertenecía a un nodosaurio , el esqueleto a un ornitópodo y la supuesta gastralia (costillas del vientre) a un pez. Esta afirmación fue refutada por Gilmore y Loris S. Russell en la década de 1930. [10]
La clasificación de Gilmore fue apoyada por los paleontólogos estadounidenses Barnum Brown y Erich Maren Schlaikjer en su revisión de 1943 de los dinosaurios con cabeza en forma de cúpula, para entonces conocidos a partir de 46 cráneos. A partir de estos especímenes, Brown y Schlaikjer nombraron las nuevas especies T. sternbergi y T. edmontonensis (ambos de Alberta), así como también trasladaron la especie grande T. wyomingensis (que fue nombrada en 1931) al nuevo género Pachycephalosaurus , junto con otras dos especies. Encontraron que T. validus era distinto de T. formosus , pero consideraron a S. brevis la forma femenina de T. validus y, por lo tanto, un sinónimo menor. En ese momento, los dinosaurios con cabeza en forma de cúpula eran considerados parientes de los ornitópodos o de los anquilosaurios. [10] En 1945, después de examinar moldes de dientes de T. formosus y S. validus , el paleontólogo estadounidense Charles M. Sternberg demostró diferencias entre los dos, y en su lugar sugirió que Troodon era un dinosaurio terópodo , y que los dinosaurios con cabeza en forma de cúpula deberían ubicarse en su propia familia. Aunque Stegoceras fue el primer miembro de esta familia en ser nombrado, Sternberg nombró al grupo Pachycephalosauridae en honor al segundo género, ya que encontró que ese nombre (que significa "lagarto de cabeza gruesa") era más descriptivo. También consideró a T. sternbergi y T. edmontonensis miembros de Stegoceras , encontró válido a S. brevis y nombró una nueva especie, S. lambei , basándose en un espécimen anteriormente referido como S. validus . [3] [11] La separación de Troodon fue apoyada por Russell en 1948, quien describió un dentario de terópodo con dientes casi idénticos a los de T. formosus . [12]
En 1953, Birger Bohlin nombró a Troodon bexelli basándose en un hueso parietal de China. [13] En 1964, Oskar Kuhn consideró esto como una especie inequívoca de Stegoceras ; S. bexelli . [14] En 1974, los paleontólogos polacos Teresa Maryańska y Halszka Osmólska concluyeron que la "gastralia" de Stegoceras eran tendones osificados, después de identificar tales estructuras en la cola del paquicefalosaurio Homalocephale . [9] En 1979, William Patrick Wall y Peter Galton nombraron a la nueva especie Stegoceras browni , basándose en una cúpula aplanada, anteriormente descrita como una hembra de S. validus por Galton en 1971. El nombre específico honra a Barnum Brown, quien encontró el espécimen holotipo (espécimen AMNH 5450 en el Museo Americano de Historia Natural ) en Alberta. [15] En 1983, Galton y Hans-Dieter Sues trasladaron a S. browni a su propio género, Ornatotholus ( ornatus en latín significa "adornado" y tholus significa "cúpula"), y lo consideraron el primer miembro americano conocido de un grupo de paquicefalosaurios de "cabeza plana", previamente conocidos de Asia. [16] En una revisión de los paquicefalosaurios de 1987, Sues y Galton enmendaron el nombre específico validus a validum , que posteriormente se ha utilizado en la literatura científica. Estos autores sinonimizaron a S. brevis , S. sternbergi y S. lambei con S. validum , encontraron que S. bexelli difería de Stegoceras en varias características y lo consideraron un paquicefalosaurio indeterminado. [3] [4] En 1998, Goodwin y sus colegas consideraron a Ornatotholus un S. validum juvenil , por lo tanto un sinónimo menor. [17]

En 2000, Robert M. Sullivan refirió a S. edmontonensis y S. brevis al género Prenocephale (hasta entonces sólo conocido de la especie mongola P. prenes ), y encontró más probable que S. bexelli perteneciera a Prenocephale que a Stegoceras , pero lo consideró un nomen dubium (nombre dudoso, sin caracteres distintivos) debido a su incompletitud, y notó que su espécimen holotipo parecía estar perdido. [18] En 2003, Thomas E. Williamson y Thomas Carr consideraron a Ornatotholus un nomen dubium , o quizás un Stegoceras juvenil . [19] En una revisión de 2003 de Stegoceras , Sullivan estuvo de acuerdo en que Ornatotholus era un sinónimo menor de Stegoceras , movió a S. lambei al nuevo género Colepiocephale , y a S. sternbergi a Hanssuesia . Afirmó que el género Stegoceras se había convertido en un taxón basura para los paquicefalosaurios norteamericanos de tamaño pequeño a mediano hasta ese momento. En ese momento, docenas de especímenes habían sido referidos a S. validum , incluidos muchos domos demasiado incompletos para ser identificados como Stegoceras con certeza. UALVP 2 sigue siendo el espécimen más completo de Stegoceras , en el que se basa la mayor parte del conocimiento científico del género. [4] S. brevis fue trasladado al nuevo género Foraminacephale en 2016 por Ryan K. Schott Schott y David C. Evans, [20] y S. bexelli a Sinocephale en 2021 por Evans y colegas. [21] En 2023, Aaron D. Dyer y colegas analizaron suturas y elementos individuales en los cráneos de los paquicefalosaurios Gravitholus y Hanssuesia , y no encontraron ninguna distinción significativa entre ellos y Stegoceras validum . Consideraron a ambos como sinónimos menores, y Gravitholus representaba la etapa final en el crecimiento de Stegoceras . [22]
En 2002, Williamson y Carr describieron un domo (espécimen NMMNH P-33983 en el Museo de Historia Natural y Ciencia de Nuevo México ) de la Cuenca de San Juan , Nuevo México , que consideraron un paquicefalosaurio juvenil de especie incierta (aunque quizás Sphaerotholus goodwini ). En 2006, Sullivan y Spencer G. Lucas lo consideraron un S. validum juvenil , lo que ampliaría considerablemente el rango de la especie. [23] [24] En 2011 , Steven E. Jasinski y Sullivan consideraron al espécimen un adulto, y lo convirtieron en el holotipo de la nueva especie Stegoceras novomexicanum , con otros dos especímenes (SMP VP-2555 y SMP VP-2790) como paratipos. [25] Un análisis filogenético de 2011 por Watabe y colegas no colocó a las dos especies de Stegoceras cerca una de la otra. [26]
En 2016, Williamson y Stephen L. Brusatte volvieron a estudiar el holotipo de S. novomexicanum y descubrieron que los paratipos no pertenecían al mismo taxón que el holotipo, y que todos los especímenes involucrados eran juveniles. Además, no pudieron determinar si el espécimen holotipo representaba la especie distinta S. novomexicanum , o si era un juvenil de S. validum o Sphaerotholus goodwini , u otro paquicefalosaurio previamente conocido. [27] En 2016, Jasinski y Sullivan defendieron la validez de S. novomexicanum ; estuvieron de acuerdo en que algunas características utilizadas para diagnosticar la especie eran indicativas de una etapa subadulta, pero presentaban características diagnósticas adicionales en el holotipo que distinguen a la especie. También señalaron algunas características adultas, que pueden indicar heterocronía (diferencia en el momento de los cambios ontogenéticos entre taxones relacionados) en la especie. Ellos aceptaron que los paratipos y otros especímenes asignados diferían del holotipo en tener cráneos más abovedados, refiriéndose en cambio a ellos como cf. S. novomexicanum (difícil de identificar), pero encontraron probable que todos ellos pertenecieran al mismo taxón (siendo los especímenes asignados adultos), debido al intervalo estratigráfico restringido y al rango geográfico. [28] Dyer y colegas encontraron que el holotipo de S. novomexicanum podría ser un Sphaerotholus goodwini inmaduro , porque los rasgos únicos propuestos de S. novomexicanum desaparecieron a través de la ontogenia en S. validum . [22]
En 2024, se describió un espécimen de Stegoceras de la Formación Aguja , y se asignó a Stegoceras con base en análisis morfométricos. Era un juvenil, muy comparable a los juveniles de S. validum , pero diferente en algunos aspectos. Lo consideraron un posible representante de una nueva especie sureña de Stegoceras , pero no de S. novomexicanum, ya que el estudio concluyó que era muy diferente de otros especímenes de Stegoceras y, por lo tanto, probablemente no referible a Stegoceras. La descripción también incluyó el holotipo de la dudosa especie Texacephale langstoni en su análisis morfométrico , donde también se encontró que era muy similar a S. validum , pero no al grado en que los autores del estudio lo referirían directamente a esa especie. Sin embargo, los autores del estudio consideraron que el holotipo de Texacephale probablemente era un espécimen adulto del género Stegoceras. [29]

Stegoceras es uno de los paquicefalosaurios norteamericanos mejor conocidos y uno de los pocos conocidos a partir de restos postcraneales ; el espécimen UALVP 2 de S. validum es el individuo de Stegoceras más completo conocido hasta la fecha. Se estima que su longitud era de unos 2 a 2,5 metros (6,6 a 8,2 pies), comparable al tamaño de una cabra . [30] [31] [32] Se ha estimado que su peso era de unos 10 a 40 kilogramos (22 a 88 libras). [33] Stegoceras era de tamaño pequeño a mediano en comparación con otros paquicefalosaurios. [3] S. novomexicanum parece haber sido más pequeño que S. validum , pero se discute si los especímenes conocidos (cráneos incompletos) son adultos o juveniles. [25] [27]

El cráneo de Stegoceras tenía una forma aproximadamente triangular cuando se lo observaba de lado, con un hocico relativamente corto. Los huesos frontal y parietal eran muy gruesos y formaban una cúpula elevada. La sutura entre estos dos elementos estaba obliterada (solo ligeramente visible en algunos especímenes), y se los denomina colectivamente "frontoparietal". La cúpula frontoparietal era amplia y tenía una superficie relativamente lisa, con solo los lados rugosos (arrugados). Se estrechaba por encima y entre las órbitas (cuencas de los ojos). El frontoparietal se estrechaba en la parte posterior, estaba encajado entre los huesos escamosos y terminaba en una depresión por encima del occipucio en la parte posterior del cráneo. Los huesos parietal y escamoso formaban una plataforma gruesa sobre el occipucio denominada plataforma parietoescamosa, cuya extensión variaba entre los especímenes. El escamoso era grande, no formaba parte de la cúpula, y la parte posterior estaba hinchada. Estaba adornado por tubérculos (o excrecencias redondeadas) espaciados irregularmente, y una fila de nodos (protuberancias) se extendía a lo largo de sus bordes superiores, terminando en un tubérculo puntiagudo (o cuerno pequeño) a cada lado en la parte posterior del cráneo. Una fila interna de tubérculos más pequeños corría paralela a la más grande. A excepción de la superficie superior de la cúpula, gran parte del cráneo estaba adornado con nodos, muchos dispuestos en filas. [3]
La gran órbita tenía forma de elipse imperfecta (con el eje más largo de adelante hacia atrás), y miraba hacia un lado y ligeramente hacia adelante. La fenestra infratemporal (abertura) detrás del ojo era estrecha e inclinada hacia atrás, y la fenestra supratemporal en la parte superior trasera del cráneo era muy reducida en tamaño, debido al engrosamiento del frontoparietal. El basicráneo (suelo de la caja craneana ) estaba acortado y distanciado de las regiones debajo de las órbitas y alrededor del paladar . El occipucio estaba inclinado hacia atrás y hacia abajo, y el cóndilo occipital estaba desviado en la misma dirección. El hueso lagrimal formaba el margen frontal inferior de la órbita, y su superficie tenía filas de ornamentación en forma de nódulos. Los huesos prefrontal y palpebral estaban fusionados y formaban una cresta gruesa sobre la órbita. El hueso yugal relativamente grande formaba el margen inferior de la órbita, extendiéndose mucho hacia adelante y hacia abajo hacia la articulación de la mandíbula. Estaba ornamentado con crestas y nudos en disposición radial. [3]
Las aberturas nasales eran grandes y miraban hacia adelante. El hueso nasal era grueso, muy esculpido y tenía un perfil convexo. Formaba un protuberancia (escudo) en la parte superior media del cráneo junto con el hueso frontal. La parte frontal inferior del premaxilar (hueso frontal de la mandíbula superior) era rugosa y engrosada. Había un pequeño foramen (agujero) en la sutura entre los premaxilares, que conducía a la cavidad nasal y posiblemente estaba conectado al órgano de Jacobson (un órgano del sentido del olfato ). El maxilar era corto y profundo, y probablemente contenía un seno . El maxilar tenía una serie de agujeros que se correspondían con la posición de cada diente allí, y estos funcionaban como pasajes para la erupción de los dientes de reemplazo. La mandíbula se articulaba con el cráneo debajo de la parte posterior de la órbita. La parte de la mandíbula inferior que contenía los dientes era larga, y la parte posterior era bastante corta. Aunque no se conservó, la presencia de un hueso predentario está indicada por facetas en la parte frontal de la mandíbula inferior. [3] Al igual que otros paquicefalosaurios, habría tenido un pico pequeño. [34]

Stegoceras tenía dientes heterodontes ( diferenciados) y tecodontos (colocados en alvéolos). Tenía filas marginales de dientes relativamente pequeños, y las filas no formaban un borde cortante recto. Los dientes estaban dispuestos oblicuamente a lo largo de las mandíbulas y se superponían ligeramente entre sí de adelante hacia atrás. En cada lado, el espécimen más completo (UALVP 2) tenía tres dientes en el premaxilar, dieciséis en el maxilar (ambos parte de la mandíbula superior) y diecisiete en el dentario de la mandíbula inferior. Los dientes en el premaxilar estaban separados de los dientes detrás del maxilar por un diastema (espacio) corto, y las dos filas en el premaxilar estaban separadas por un espacio sin dientes en la parte delantera. Los dientes en la parte delantera de la mandíbula superior (premaxilar) y la mandíbula inferior delantera eran similares; estos tenían coronas más altas, puntiagudas y recurvadas, y un "talón" en la parte posterior. Los dientes delanteros en la mandíbula inferior eran más grandes que los de la mandíbula superior. Los bordes frontales de las coronas tenían ocho dentículos (dentaduras) y el borde posterior tenía entre nueve y once. Los dientes en la parte posterior de la mandíbula superior (maxilar) e inferior eran triangulares en vista lateral y comprimidos en vista frontal. Tenían raíces largas que eran ovaladas en sección, y las coronas tenían un cíngulo marcado en sus bases. Los dentículos aquí estaban comprimidos y dirigidos hacia la parte superior de las coronas. Tanto el lado externo como el interno de las coronas de los dientes tenían esmalte , y ambos lados estaban divididos verticalmente por una cresta. Cada borde tenía alrededor de siete u ocho dentículos, y el borde frontal generalmente tenía la mayoría. [3]
El cráneo de Stegoceras se puede distinguir de los de otros paquicefalosaurios por características tales como su pronunciada plataforma parietoescamosa (aunque esta se hizo más pequeña con la edad), la "incipiente" cúpula de su frontoparietal (aunque la cúpula aumentó con la edad), sus huesos nasales inflados, su ornamentación de tubérculos en los lados y la parte posterior de los huesos escamosos, filas de hasta seis tubérculos en el lado superior de cada escamoso y hasta dos nodos en la proyección posterior del parietal. También se distingue por su falta de ornamentación nasal y por tener un diastema reducido. [4] [35] El cráneo de S. novomexicanum se puede distinguir del de S. validum en características tales como la extensión posterior del hueso parietal que es más reducida y triangular, teniendo fenestras supratemporales más grandes (aunque esto puede deberse al posible estado juvenil de los especímenes), y teniendo contactos de sutura aproximadamente paralelos entre el escamoso y el parietal. También parece haber tenido un protuberancia frontal más pequeña que S. validum , [25] [27] y parece haber sido más grácil en general. [28]
La columna vertebral de Stegoceras no se conoce completamente. La articulación entre las zigofisis (procesos articulares) de las sucesivas vértebras dorsales (espalda) parece haber impedido el movimiento lateral de la columna vertebral, lo que la hacía muy rígida, y se fortalecía aún más mediante tendones osificados . [3] Aunque no se conocen las vértebras del cuello, el cóndilo occipital hacia abajo (que se articula con la primera vértebra del cuello) indica que el cuello se mantenía en una postura curva, como la forma de "S" o "U" de la mayoría de los cuellos de los dinosaurios. [36] Según su posición en Homalocephale , los tendones osificados encontrados con UALVP 2 habrían formado una intrincada " cesta caudal " en la cola, que consta de filas paralelas, con las extremidades de cada tendón en contacto con el siguiente sucesivamente. Estas estructuras se denominan miorhabdoi y, por lo demás, solo se conocen en peces teleósteos ; Esta característica es exclusiva de los paquicefalosaurios entre los animales tetrápodos (de cuatro extremidades) y puede haber funcionado para endurecer la cola. [9]
La escápula (omóplato) era más larga que el húmero (hueso del brazo superior); su hoja era delgada y estrecha, y ligeramente torcida, siguiendo el contorno de las costillas. La escápula no se expandía en el extremo superior, pero estaba muy expandida en la base. El coracoides era principalmente delgado y en forma de placa. El húmero tenía un eje delgado, estaba ligeramente torcido a lo largo de su longitud y estaba ligeramente arqueado. La cresta deltopectoral (donde se unían los músculos deltoides y pectorales ) estaba débilmente desarrollada. Los extremos del cúbito estaban expandidos y las crestas se extendían a lo largo del eje. El radio era más robusto que el cúbito, lo cual es inusual. Cuando se ve desde arriba, la cintura pélvica era muy ancha para un arcosaurio bípedo , y se ensanchaba hacia la parte trasera. La amplitud de la región pélvica puede haber acomodado una extensión posterior del intestino. El íleon era alargado y el isquion era largo y delgado. Aunque no se conoce el pubis , probablemente era de tamaño reducido como el de Homalocephale . El fémur (hueso del muslo) era delgado y curvado hacia adentro, la tibia era delgada y retorcida, y el peroné era delgado y ancho en el extremo superior. El metatarso del pie parece haber sido estrecho, y el único ungueal (hueso de la garra) conocido de un dedo del pie era delgado y ligeramente curvado. [3] Aunque no se conocen completamente las extremidades de Stegoceras , lo más probable es que fueran como otros paquicefalosaurios en tener manos de cinco dedos y cuatro dedos en los pies. [34]

Durante la década de 1970, se describieron más géneros de paquicefalosaurios a partir de fósiles asiáticos, lo que proporcionó más información sobre el grupo. En 1974, Maryańska y Osmólska concluyeron que los paquicefalosaurios son lo suficientemente distintos como para justificar su propio suborden dentro de Ornithischia, Pachycephalosauria. En 1978, el paleontólogo chino Dong Zhiming dividió a Pachycephalosauria en dos familias: los Pachycephalosauridae de cabeza en forma de cúpula (que incluía a Stegoceras ) y los Homalocephalidae de cabeza plana (originalmente escritos Homalocephaleridae). [37] Wall y Galton no encontraron que el estatus de suborden para los paquicefalosaurios estuviera justificado en 1979. [15] En la década de 1980, las afinidades de los paquicefalosaurios dentro de Ornithischia estaban sin resolver. Las principales opiniones en pugna eran que el grupo estaba más próximo a los ornitópodos o a los ceratopsianos, esta última opinión debido a las similitudes entre el esqueleto de Stegoceras y el ceratopsiano "primitivo" Protoceratops . En 1986, el paleontólogo estadounidense Paul Sereno apoyó la relación entre los paquicefalosaurios y los ceratopsianos, y los unió en el grupo Marginocephalia , basándose en características craneales similares, como la estructura en "plataforma" sobre el occipucio. Reconoció que la evidencia para esta agrupación no era abrumadora, pero la validez del grupo fue apoyada por Sues y Galton en 1987. [3]
A principios del siglo XXI, se conocían pocos géneros de paquicefalosaurios a partir de restos postcraneales, y muchos taxones solo se conocían a partir de domos, lo que dificultaba la clasificación dentro del grupo. Por lo tanto, los paquicefalosaurios se definen principalmente por características craneales, como el frontoparietal plano a abovedado, la barra ancha y aplanada a lo largo de los huesos postorbital y escamoso, y los huesos escamosos que son placas profundas en el occipucio. [32] En 1986, Sereno había dividido a los paquicefalosaurios en diferentes grupos según la extensión de la cúpula de sus cráneos (agrupados en taxones ahora inválidos como "Tholocephalidae" y "Domocephalinae"), y en 2000 consideró al "parcialmente" abovedado Stegoceras una transición entre los géneros supuestamente "primitivos" de cabeza plana y los avanzados "totalmente" abovedados (como Pachycephalosaurus ). [38] La división de cabezas abombadas/cabezas planas de los paquicefalosaurios fue abandonada en los años siguientes, ya que las cabezas planas se consideraron rasgos pedomórficos (similares a los juveniles) o derivados en la mayoría de las revisiones, pero no un rasgo sexualmente dimórfico . En 2006, Sullivan argumentó en contra de la idea de que el grado de abombamiento fuera útil para determinar las afinidades taxonómicas entre los paquicefalosaurios. [32] En 2003, Sullivan encontró que el propio Stegoceras era más basal (o "primitivo") que los miembros "completamente abombados" de la subfamilia Pachycephalosaurinae, y elaboró las conclusiones a las que llegó Sereno en 1986. [4]
Un análisis filogenético de 2013 realizado por Evans y colegas encontró que algunos géneros de paquicefalosaurios de cabeza plana estaban más estrechamente relacionados con taxones "totalmente" abovedados que con los Stegoceras "incompletamente" abovedados , lo que sugiere que representan juveniles de taxones abovedados, y que las cabezas planas no indican afinidades taxonómicas. [39] El cladograma a continuación muestra la ubicación de Stegoceras dentro de Pachycephalosauridae según Schott y colegas, 2016: [20]

La biogeografía y la historia evolutiva temprana de los paquicefalosaurios son poco conocidas y solo pueden ser aclaradas por nuevos descubrimientos. Los paquicefalosaurios aparecen abruptamente en el registro fósil y están presentes tanto en América del Norte como en Asia, por lo que se desconoce cuándo se originaron por primera vez y desde qué dirección se dispersaron. Los miembros más antiguos conocidos del grupo (como Acrotholus ) son "totalmente abovedados" y se los conoce desde la etapa Santoniana del período Cretácico Superior (hace unos 84 millones de años). Esto es anterior a los supuestamente más primitivos Stegoceras del Campaniano Medio (hace 77 millones de años) y Homalocephale del Maastrichtiano Inferior (hace 70 millones de años), por lo que la abovedamiento del cráneo puede ser un rasgo homoplástico (una forma de evolución convergente ). La aparición tardía de los paquicefalosaurios en comparación con los ceratopsianos relacionados indica un largo linaje fantasma (inferido, pero que falta en el registro fósil) que abarca 66 millones de años, desde el Jurásico Superior hasta el Cretácico. Dado que los paquicefalosaurios eran principalmente pequeños, esto puede deberse a un sesgo tafonómico ; los animales más pequeños tienen menos probabilidades de conservarse mediante fosilización. Los huesos más delicados también tienen menos probabilidades de conservarse, por lo que los paquicefalosaurios se conocen principalmente por sus cráneos robustos. [4] [39]

No se sabe con certeza qué comían los paquicefalosaurios; al tener dientes muy pequeños y estriados, no podrían haber masticado plantas duras y fibrosas con tanta eficacia como otros dinosaurios del mismo período. Se supone que sus dientes afilados y dentados eran ideales para una dieta mixta de hojas, semillas, frutas e insectos. [40] Stegoceras puede haber tenido una dieta completamente herbívora, ya que las coronas de los dientes eran similares a las de los lagartos iguánidos . Los dientes premaxilares muestran facetas de desgaste por el contacto con el hueso predentario, y los dientes maxilares tienen facetas de desgaste dobles similares a las observadas en otros dinosaurios ornitisquios. Cada tercer diente maxilar de UALVP 2 son dientes de reemplazo en erupción , y el reemplazo de dientes ocurrió en progresión regresiva en tres secuenciales. La región occipital de Stegoceras estaba bien delimitada para la inserción de los músculos y se cree que el movimiento de la mandíbula de Stegoceras y otros paquicefalosaurios se limitaba principalmente a movimientos de arriba a abajo con solo una ligera capacidad para la rotación de la mandíbula. Esto se basa en la estructura de la mandíbula y el microdesgaste dental y las facetas de desgaste de los dientes indican que la fuerza de mordida se usaba más para cortar que para aplastar. [3] [41]
En 2021, el paleontólogo canadiense Michael N. Hudgins y sus colegas examinaron los dientes de Stegoceras y Thescelosaurus y descubrieron que, si bien ambos tenían dientes heterodontes, podían distinguirse estadísticamente entre sí. Debido a su amplio rostro y dientes más uniformes, Stegoceras era un devorador indiscriminado de grandes cantidades de vegetación, mientras que los dientes y el rostro estrecho de Thescelosaurus indican que era un devorador selectivo. Los paquicefalosaurios y los tescelosáuridos se encuentran en las mismas formaciones de América del Norte, y parece que su coexistencia fue posible porque ocupaban ecomorfoespacios diferentes (aunque Stegoceras y Thescelosaurus no eran contemporáneos). [42]
En 1989, Emily B. Griffin descubrió que Stegoceras y otros paquicefalosaurios tenían un buen sentido del olfato (olfato), basándose en el estudio de endomoldes craneales que mostraban grandes bulbos olfatorios en el cerebro. [43] En 2014, Jason M. Bourke y sus colegas descubrieron que Stegoceras habría necesitado cornetes nasales cartilaginosos en la parte delantera de los conductos nasales para que el flujo de aire llegara a la región olfativa. La evidencia de la presencia de esta estructura es una cresta ósea a la que podría haberse adherido. El tamaño de la región olfativa también indica que Stegoceras tenía un agudo sentido del olfato. Los investigadores descubrieron que el dinosaurio podría haber tenido un cornete en forma de pergamino (como en un pavo ) o uno ramificado (como en un avestruz ), ya que ambos podrían haber dirigido el aire a la región olfativa. El sistema de vasos sanguíneos en los conductos también sugiere que los cornetes servían para enfriar la sangre arterial caliente del cuerpo que se dirigía al cerebro. El cráneo del espécimen UALVP 2 de S. validum era adecuado para un estudio de este tipo debido a su excepcional conservación; tiene tejido blando osificado en la cavidad nasal, que de otro modo sería cartilaginoso y, por lo tanto, no se conservaría mediante mineralización. [44]

Históricamente se han propuesto varias explicaciones para la variación observada en los cráneos de Stegoceras y otros paquicefalosaurios. Brown y Schlaikjer sugirieron que había dimorfismo sexual en el grado de abombamiento, y plantearon la hipótesis de que los especímenes de cabeza plana como AMNH 5450 ( Ornatotholus ) representaban la forma femenina de Stegoceras . Esta idea fue apoyada por un estudio morfométrico de 1981 realizado por Champan y colegas, que encontró que los machos tenían cúpulas más grandes y gruesas. [35] [45] Después de que se descubrieron otros paquicefalosaurios de cabeza plana, se propuso que el grado de abombamiento fuera una característica con importancia taxonómica, y por lo tanto AMNH 5450 se consideró un taxón distinto a partir de 1979. En 1998, Goodwin y sus colegas propusieron que la inflación de la cúpula era una característica ontogénica que cambiaba con la edad, basándose en un estudio histológico de un cráneo de S. validum que mostraba que la cúpula estaba formada por hueso vascular de rápido crecimiento, lo que era coherente con un aumento de la formación de la cúpula con la edad. Estos autores descubrieron que las características supuestamente distintivas de Ornatotholus podrían ser fácilmente el resultado de la ontogenia. [35] [17]
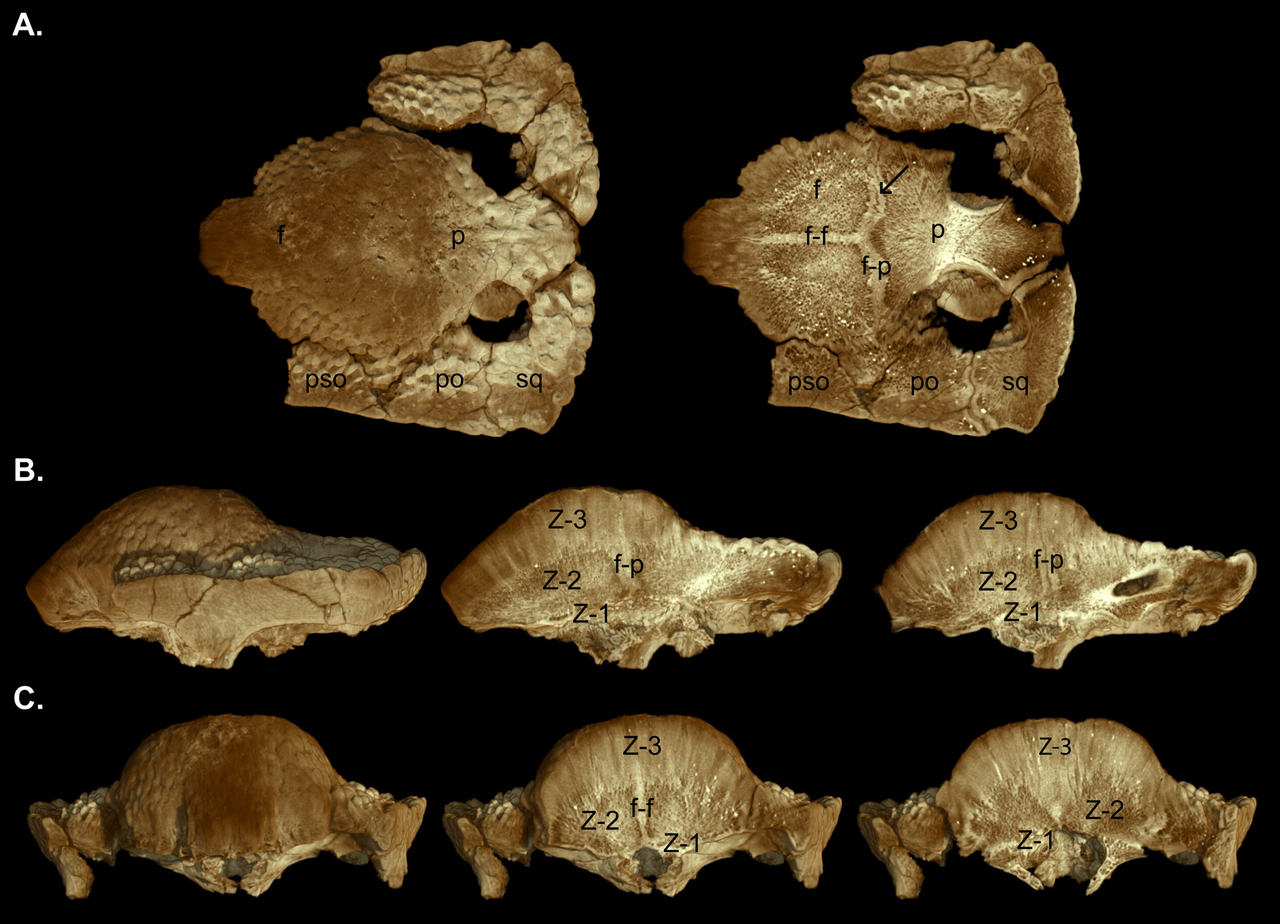
En 2003, Williamson y Carr publicaron una serie hipotética de crecimiento de S. validum , mostrando a Ornatotholus como la etapa juvenil. Sugirieron que los juveniles se caracterizaban por un techo frontoparietal plano y engrosado, con fenestras supratemporales más grandes y tachonadas de tubérculos y nodos muy espaciados. La plataforma parietoescamosa no se redujo en tamaño y la sutura frontoparietal estaba abierta. Los subadultos tenían cúpulas en forma de montículo, con la parte posterior del parietal y el techo del cráneo planas. Las fenestras supratemporales mostraron asimetría en tamaño y el cierre de la sutura frontoparietal fue variable. Los nodos se estiraron o casi se borraron a medida que la cúpula se expandía durante el crecimiento, con una superficie teselada restante. El patrón a menudo se borraba en el punto más alto (ápice) de la cúpula, el área donde se producía la máxima expansión. Los tubérculos del cráneo se estiraban en diferentes direcciones, y los que se encontraban en el margen de la plataforma parietoescamosa pueden haber sido tubérculos hipertrofiados (agrandados). La parte posterior y los lados de los cráneos de los subadultos y adultos estaban ornamentados por tubérculos menos modificados. Antes de ser incorporados a la cúpula en expansión, los huesos del cráneo se expandieron, lo que dio lugar a uniones entre estos huesos. La cúpula de los adultos era amplia y convexa, e incorporaba la mayor parte de la plataforma, que se redujo en tamaño y sobresalía del occipucio como un "labio" grueso. Las fenestras supratemporales estaban cerradas, pero la sutura entre los huesos frontoparietales y los huesos del cráneo conectados no siempre estaba cerrada en los adultos y subadultos. [19]
En 2011, Schott y sus colegas realizaron un análisis más exhaustivo de la ontogenia de la cúpula craneal en S. validum . El estudio encontró que la plataforma parietoescamosa conservaba la disposición de la ornamentación a lo largo del crecimiento, y que la vascularidad de las cúpulas frontoparietales disminuía con el tamaño. También encontró que la forma y el tamaño de la cúpula estaban fuertemente correlacionados con el crecimiento, y que el crecimiento era alométrico (en contraste con el isométrico ) de plano a abovedado, lo que apoya a Ornatotholus como un Stegoceras juvenil . También plantearon la hipótesis de que este modelo de crecimiento de la cúpula, con cambios dramáticos de juvenil a adulto, era la trayectoria de desarrollo común de los paquicefalosaurios. Estos investigadores notaron que aunque la observación de Williamson y Carr de que las fenestras supratemporales se cerraban con la edad era generalmente correcta, todavía había un alto grado de variación individual en el tamaño de estas fenestras, independientemente del tamaño de la frontoparietal, y esta característica puede, por lo tanto, haber sido independiente de la ontogenia. [35]
Un estudio de 2012 realizado por Schott y Evans descubrió que la cantidad y la forma de los nodos individuales en la plataforma escamosa de los cráneos de S. validum examinados variaban considerablemente, y que esta variabilidad no parecía correlacionarse con cambios ontogénicos, sino que se debía a la variación individual. Estos investigadores no encontraron correlación entre el ancho de las fenestras supratemporales y el tamaño de la plataforma escamosa. [46]
La función de las cúpulas de los paquicefalosaurios ha sido debatida, y Stegoceras ha sido utilizado como modelo para la experimentación en varios estudios. La cúpula ha sido interpretada principalmente como un arma utilizada en el combate intraespecífico , una estructura de exhibición sexual o un medio para el reconocimiento de especies . [47] [48]

La hipótesis de que los cráneos abovedados de Stegoceras y otros paquicefalosaurios se utilizaban para golpearse la cabeza fue sugerida por primera vez por el paleontólogo estadounidense Edwin Colbert en 1955. En 1970 y 1971, Galton profundizó en esta idea y argumentó que si la cúpula era simplemente ornamental, habría sido menos densa y que la estructura era ideal para resistir la fuerza. Galton sugirió que cuando Stegoceras sostenía su cráneo verticalmente, perpendicular al cuello, la fuerza se transmitiría desde el cráneo, con pocas posibilidades de que se dislocara, y por lo tanto la cúpula podría haber sido utilizada como un ariete. Creía que era poco probable que se hubiera utilizado principalmente como defensa contra los depredadores, porque la cúpula en sí carecía de púas, y las de la plataforma parietoescamosa estaban en una posición "ineficaz", pero la encontró compatible con la competencia intraespecífica. Galton imaginó que las cúpulas se golpeaban entre sí, mientras que la columna vertebral se mantenía en posición horizontal. Esto podría hacerse mientras se enfrentaban entre sí mientras se golpeaban, o mientras se cargaban con la cabeza baja (de manera análoga a las ovejas y cabras modernas). También señaló que la rigidez de la espalda habría sido útil al usar la cabeza para este propósito. En 1978, Sues estuvo de acuerdo con Galton en que la anatomía de los paquicefalosaurios era consistente con la transmisión de la tensión del impacto de cúpula a cúpula, basándose en pruebas con modelos de plexiglás . El impacto sería absorbido a través del cuello y el cuerpo, y los ligamentos y músculos del cuello evitarían lesiones por golpes de refilón (como en los borregos cimarrones modernos ). Sues también sugirió que los animales podrían haberse golpeado los flancos entre sí. [36] [49] [50]
En 1997, el paleontólogo estadounidense Kenneth Carpenter señaló que las vértebras dorsales de la espalda del paquicefalosaurio Homalocephale muestran que la espalda se curvaba hacia abajo justo antes del cuello (que no se conservó), y a menos que el cuello se curvara hacia arriba, la cabeza apuntaría al suelo. Por lo tanto, dedujo que los cuellos de Stegoceras y otros paquicefalosaurios se mantenían en una postura curvada (como es la norma en los dinosaurios), y que, por lo tanto, no habrían sido capaces de alinear la cabeza, el cuello y el cuerpo horizontalmente, lo que sería necesario para transmitir la tensión. Sus cuellos tendrían que mantenerse por debajo del nivel de la espalda, lo que habría corrido el riesgo de dañar la médula espinal en caso de impacto. Los borregos cimarrones y los bisontes modernos superan este problema al tener fuertes ligamentos desde el cuello hasta las altas espinas neurales sobre los hombros (que absorben la fuerza del impacto), pero tales características no se conocen en los paquicefalosaurios. Estos animales también absorben la fuerza del impacto a través de cámaras sinusales en la base de sus cuernos, y sus frentes y cuernos forman una amplia superficie de contacto, a diferencia de la estrecha superficie de las cúpulas de los paquicefalosaurios. Debido a que la cúpula de Stegoceras era redondeada, habría proporcionado un área muy pequeña para el impacto potencial, y las cúpulas habrían chocado entre sí (a menos que el impacto estuviera perfectamente centrado). Los paquicefalosaurios combatientes habrían tenido dificultades para verse entre sí mientras tenían la cabeza agachada, debido a las crestas óseas sobre los ojos. [36]
Debido a los problemas que encontró con la hipótesis de los cabezazos, Carpenter sugirió en cambio que las cúpulas eran adaptaciones para los cabezazos en los flancos (como se ve en algunos grandes mamíferos africanos); imaginó que dos animales estarían de pie en paralelo, uno frente al otro o en la misma dirección, y dirigirían los golpes al costado del oponente. En consecuencia, la anchura corporal relativamente grande de los paquicefalosaurios puede haber servido para proteger los órganos vitales de daños durante los cabezazos en los flancos. Es posible que Stegoceras y otros paquicefalosaurios similares hubieran asestado los golpes con un movimiento del cuello desde un lado y una rotación de la cabeza. Los lados superiores de la cúpula tienen la mayor superficie y pueden haber sido el punto de impacto. El grosor de la cúpula habría aumentado la potencia de un golpe en los costados, y esto garantizaría que el oponente sintiera la fuerza del impacto, sin resultar gravemente herido. El borde óseo sobre la órbita puede haber protegido el ojo del agresor al dar un golpe. Carpenter sugirió que los paquicefalosaurios primero habrían realizado una exhibición de amenaza moviendo y presentando sus cabezas para mostrar el tamaño de sus cúpulas (intimidación), y luego se habrían golpeado entre sí, hasta que un oponente señaló su sumisión. [36]
En 2008, Eric Snively y Adam Cox probaron el rendimiento de los cráneos de paquicefalosaurio 2D y 3D a través del análisis de elementos finitos , y descubrieron que podían soportar un impacto considerable; una mayor bóveda de las cúpulas permitía mayores fuerzas de impacto. También consideraron probable que las cúpulas de los paquicefalosaurios estuvieran cubiertas de queratina , un material fuerte que puede soportar mucha energía sin sufrir daños permanentes (como los osteodermos de los cocodrilos ), y por lo tanto incorporaron queratina en su fórmula de prueba. [51] En 2011, Snively y Jessica M. Theodor realizaron un análisis de elementos finitos simulando impactos en la cabeza con cráneos escaneados por TC de S. validum (UALVP 2), Prenocephale prenes y varios artiodáctilos existentes que dan cabezazos . Descubrieron que las correlaciones entre los golpes en la cabeza y las morfologías del cráneo encontradas en los animales vivos también existían en los paquicefalosaurios estudiados. Tanto Stegoceras como Prenocephale tenían formas craneales similares a las del borrego cimarrón, con hueso esponjoso protegiendo el cerebro. También compartían similitudes en la distribución de las regiones compactas y esponjosas con el borrego cimarrón, el duiker de vientre blanco y la jirafa . Se descubrió que el duiker de vientre blanco era el análogo morfológico más cercano a Stegoceras ; esta especie que da cabezazos tiene una cúpula que es más pequeña pero igualmente redondeada. Stegoceras era más capaz de disipar la fuerza que los artiodáctilos que se dan cabezazos con fuerzas altas, pero las cúpulas menos vascularizadas de los paquicefalosaurios más antiguos, y posiblemente la capacidad disminuida para curarse de las heridas, argumentaban en contra de tal combate en individuos más viejos. El estudio también probó los efectos de una cubierta queratinosa de la cúpula y descubrió que ayudaba en el rendimiento. Aunque Stegoceras carecía de los senos neumáticos que se encuentran debajo del punto de impacto en los cráneos de los artiodáctilos que golpean la cabeza, tenía en cambio puntales vasculares que podrían haber actuado de manera similar como soportes, así como conductos para alimentar el desarrollo de una cubierta de queratina. [52]
En 2012, Caleb M. Brown y Anthony P. Russell sugirieron que las colas rígidas probablemente no se usaban como defensa contra los golpes en los flancos, sino que podían haber permitido a los animales adoptar una postura de trípode durante el combate intraespecífico, con la cola como apoyo. Brown y Russell descubrieron que la cola podía ayudar a resistir la carga de compresión, tracción y torsión cuando el animal daba o recibía golpes con la cúpula. [9] Un estudio de 2013 de Joseph E. Peterson y colegas identificó lesiones en los cráneos de Stegoceras y otros paquicefalosaurios, que se interpretaron como infecciones causadas por traumatismos. Se encontraron lesiones en el 22% de los cráneos de paquicefalosaurios muestreados (una frecuencia consistente en todos los géneros), pero no estaban presentes en los especímenes de cabeza plana (que se han interpretado como juveniles o hembras), lo que es consistente con el uso en el combate intraespecífico (por territorio o parejas). La distribución de las lesiones en estos animales tendía a concentrarse en la parte superior de la cúpula, lo que respalda el comportamiento de dar cabezazos. Los cabezazos probablemente darían lugar a menos lesiones, que se concentrarían en los lados de la cúpula. Estas observaciones se realizaron al comparar las lesiones con las de los cráneos y los flancos de los esqueletos de ovejas modernas. Los investigadores observaron que los animales modernos que dan cabezazos utilizan sus armas tanto para el combate como para exhibirse, y que, por lo tanto, los paquicefalosaurios también podrían haber utilizado sus cúpulas para ambas cosas. Mostrar un arma y estar dispuesto a usarla puede ser suficiente para resolver disputas en algunos animales. [47]
Bryan RS Moore y sus colegas examinaron y reconstruyeron la musculatura de las extremidades de Stegoceras en 3D en 2022, utilizando el espécimen muy completo UALVP 2 como base. Descubrieron que la musculatura de las extremidades anteriores era conservadora, particularmente en comparación con las de los primeros dinosaurios saurisquios bípedos , pero la musculatura pélvica y de las extremidades traseras era en cambio más derivada (o "avanzada"), debido a las peculiaridades del esqueleto. Estas áreas tenían músculos grandes, y combinados con la pelvis ancha y las extremidades traseras robustas (y posiblemente ligamentos agrandados), esto dio como resultado una estructura pélvica fuerte y estable que habría ayudado durante los cabezazos entre individuos. Dado que las cúpulas craneales de los paquicefalosaurios crecieron con alometría positiva , y pueden haber sido utilizadas en combate, estos investigadores sugirieron que también podría haber sido el caso de los músculos de las extremidades traseras, si se usaban para impulsar el cuerpo hacia adelante durante los cabezazos. Advirtieron que si bien UALVP 2 es muy completo para un paquicefalosaurio, su estudio estaba limitado porque faltaban grandes porciones de su columna vertebral y elementos de las extremidades externas. [53]

En 1987, J. Keith Rigby y sus colegas sugirieron que las cúpulas de los paquicefalosaurios eran órganos de intercambio de calor utilizados para la termorregulación , basándose en sus "estructuras radiantes" internas ( trabéculas ). Esta idea fue apoyada por algunos otros escritores a mediados de la década de 1990. [48] En 1998, Goodwin y sus colegas consideraron que la falta de senos en el cráneo de Stegoceras y la red de hueso vascular en forma de " panal " en la cúpula no eran adecuadas para los cabezazos, y señalaron que los huesos adyacentes a la cúpula corrían el riesgo de fracturarse durante dicho contacto. Basándose en la idea de que los tendones osificados que endurecían las colas de Stegoceras y otros paquicefalosaurios les permitían adoptar una postura trípode (sugerida por primera vez por Maryańska y Osmólska en 1974), Goodwin et al. sugirieron que estas estructuras podrían haber protegido la cola contra los cabezazos en los flancos, o que la cola misma podría haber sido utilizada como arma. [17] En 2004, Goodwin y sus colegas estudiaron la histología craneal de los paquicefalosaurios y descubrieron que la vascularidad (incluidas las trabéculas) de las cúpulas disminuía con la edad, lo que encontraron inconsistente con una función en los cabezazos o el intercambio de calor. También sugirieron que una capa densa de fibras de Sharpey cerca de la superficie de la cúpula indicaba que tenía una cubierta externa en vida, lo que hace imposible conocer la forma de la cúpula en un animal vivo. Estos investigadores, en cambio, concluyeron que las cúpulas eran principalmente para el reconocimiento y la comunicación de especies (como en algunos bóvidos africanos ) y que el uso en la exhibición sexual era solo secundario. Además, especularon que la cubierta externa de las cúpulas era de colores brillantes en vida, o puede haber cambiado de color estacionalmente. [48]
En 2011, los paleontólogos estadounidenses Kevin Padian y John R. Horner propusieron que las "estructuras extrañas" en los dinosaurios en general (incluyendo domos, volantes, cuernos y crestas) se utilizaban principalmente para el reconocimiento de especies, y descartaron otras explicaciones por no estar respaldadas por evidencia. Entre otros estudios, estos autores citaron el artículo de 2004 de Goodwin et al. sobre los domos de los paquicefalosaurios como apoyo a esta idea, y señalaron que dichas estructuras no parecían ser sexualmente dimórficas. [54] En una respuesta a Padian y Horner el mismo año, Rob J. Knell y Scott D. Sampson argumentaron que el reconocimiento de especies no era improbable como una función secundaria de las "estructuras extrañas" en los dinosaurios, pero que la selección sexual (utilizada en la exhibición o el combate para competir por parejas) era una explicación más probable, debido al alto costo de desarrollarlas, y porque dichas estructuras parecen ser muy variables dentro de las especies. [55] En 2013, los paleontólogos británicos David E. Hone y Darren Naish criticaron la "hipótesis de reconocimiento de especies" y argumentaron que ningún animal actual utiliza tales estructuras principalmente para el reconocimiento de especies, y que Padian y Horner habían ignorado la posibilidad de selección sexual mutua (donde ambos sexos están ornamentados). [56]
En 2012, Schott y Evans sugirieron que la regularidad en la ornamentación escamosa a lo largo de la ontogenia de Stegoceras era consistente con el reconocimiento de especies, pero el cambio de frontoparietales planos a abovedados en la edad avanzada sugiere que la función de esta característica cambió a través de la ontogenia, y tal vez fue seleccionada sexualmente, posiblemente para el combate intraespecífico. [46] Dyer y colegas encontraron en 2023 que los especímenes de Stegoceras diferían en el grosor del saliente frontonasal, y que los cráneos con más patologías óseas eran aquellos con los salientes más altos, lo que consideraron una indicación de que la variación en el grosor del saliente representa una variación intersexual. [22] En 2023, Horner y colegas afirmaron que dado que el domo y la ornamentación asociada de Stegoceras y la ornamentación de Pachycephalosaurus se desarrollaron temprano en la vida, esto indica que se usaban para la comunicación visual, de modo que los juveniles podían reconocer a otros juveniles y a los adultos a otros adultos. No descartaron que estas características pudieran haber sido utilizadas para otros propósitos, incluido el de dar cabezazos, pero no consideraron que el trauma observado en los especímenes fuera una prueba de ello. También sugirieron que las características de algunos cráneos de paquicefalosáuridos indican que la cúpula habría soportado una estructura queratinosa más grande que un simple casquete. [57]

S. validum se conoce del Grupo Belly River del Cretácico Superior (el equivalente canadiense del Grupo Judith River en los EE. UU.), y se han recuperado especímenes de la Formación Dinosaur Park (Campaniano tardío, 76,5 a 75 millones de años) en el Parque Provincial Dinosaurio (incluido el espécimen lectotipo), y la Formación Oldman (Campaniano medio, 77,5 a 76,5 millones de años) de Alberta, Canadá. Los paquicefalosaurios Hanssuesia (si no es un sinónimo de Stegoceras ) y Foraminacephale también se conocen de ambas formaciones. [4] [22] S. novomexicanum se conoce de Fruitland (Campaniano tardío, alrededor de 75 millones de años) y la Formación Kirtland inferior ( Campaniano tardío , alrededor de 74 millones de años) de Nuevo México, y si esta especie pertenece correctamente a Stegoceras , el género habría tenido una amplia distribución geográfica. [25] La presencia de paquicefalosaurios similares tanto en el oeste como en el norte de América del Norte durante el último Cretácico muestra que eran una parte importante de las faunas de dinosaurios allí. [27]
Tradicionalmente se ha sugerido que los paquicefalosaurios habitaban en entornos montañosos; el desgaste de sus cráneos supuestamente era resultado de haber sido arrastrados por el agua desde las zonas altas, y las comparaciones con los borregos cimarrones reforzaron la teoría. En 2014, Jordan C. Mallon y Evans cuestionaron esta idea, ya que el desgaste y las ubicaciones originales de los cráneos no son consistentes con el hecho de haber sido transportados de esa manera, y en su lugar propusieron que los paquicefalosaurios norteamericanos habitaban en entornos aluviales (asociados con el agua) y en llanuras costeras . [58]
La Formación Dinosaur Park se interpreta como un entorno de bajo relieve de ríos y llanuras aluviales que se volvieron más pantanosos e influenciados por las condiciones marinas con el tiempo a medida que el Mar Interior Occidental transgredía hacia el oeste. [59] El clima era más cálido que el actual Alberta, sin heladas , pero con estaciones más húmedas y secas. Las coníferas eran aparentemente las plantas dominantes del dosel , con un sotobosque de helechos , helechos arborescentes y angiospermas . [60] Dinosaur Park es conocido por su diversa comunidad de herbívoros. Además de Stegoceras , la formación también ha producido fósiles de los ceratopsianos Centrosaurus , Styracosaurus y Chasmosaurus , los hadrosáuridos Prosaurolophus , Lambeosaurus , Gryposaurus , Corythosaurus y Parasaurolophus , y los anquilosaurios Edmontonia y Euoplocephalus . Los terópodos presentes incluyen a los tiranosáuridos Gorgosaurus y Daspletosaurus . [61] Otros dinosaurios conocidos de la Formación Oldman incluyen al hadrosaurio Brachylophosaurus , los ceratopsianos Coronosaurus y Albertaceratops , ornitomímidos , terizinosaurios y posiblemente anquilosaurios. Los terópodos incluyeron troodóntidos , oviraptorosaurios , el dromeosáurido Saurornitholestes y posiblemente un tiranosáurido albertosaurino. [62]
{kind=link}