Unión de Holliday
En biología, las uniones de Holliday funcionan como intermediarios en la recombinación genética; estas uniones o cruces (junctions) tienen generalmente una secuencia simétrica, por lo que son móviles.
La estructura recibe su nombre por Robin Holliday, que propuso su existencia en 1964.
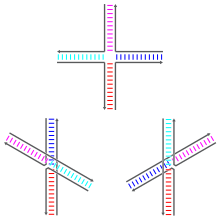
[2] La forma no amontonada es una conformación extendida casi plana y cuadrada.
Por otra parte, los confórmeros amontonados tienen dos dominios en doble hélice continuos separados por un ángulo de unos 60° hacia la derecha.
[2] En uniones con secuencias simétricas, el punto de ramificación es móvil y puede migrar en un proceso aleatorio.
[2] Adicionalmente, pueden originarse estructuras cruciformes que implican a las uniones de Holliday para aliviar la tensión en la hélice de secuencias simétricas en ADN superenrollado.
La separación del conjunto original de fibras origina dos moléculas que pueden presentar conversión génica pero no sobrecruzamiento cromosómico, mientras que la separación del otro conjunto de dos fibras causa que las moléculas recombinantes resultantes presenten sobrecruzamiento.
A medida que la burbuja se acerca al ADN roto, la fibra antisentido 5' más larga invade de nuevo la fibra sentido de esta porción del ADN, transcribiendo una segunda copia.
Cuando termina la replicación, ambas colas son reconectadas para formar dos uniones de Holliday, que son después unidas por proteínas de variadas maneras.
Como los complejos de uniones de Holliday aislados son demasiado flexibles para ensamblarse en matrices (arrays) ordenadas grandes, los motivos estructurales con muchas uniones de Holliday se utilizan para crear "baldosas" rígidas que pueden después ensamblarse en "matrices" más grandes.
El complejo puede teóricamente ser diseñado para forzar las uniones a tener una orientación paralela o antiparalela, pero en la práctica las variedades antiparalelas se comportan mejor, y las versiones paralelas raramente si usan.
[14][15] El motivo estructural DX es también la pieza de construcción fundamental del método ADN origami, que se utiliza para hacer estructuras bi o tridimensionles más grandes de forma arbitraria.
En lugar de usar baldosas DX individuales, se pliega una hebra de entramado largo sencillo en la forma deseada usando varias fibras grapa cortas.
[14][15] Robin Holliday propuso esta estructura como parte de su modelo de recombinación homóloga en 1964, basado en su investigación en los organismos Ustilago maydis y Saccharomyces cerevisiae.
Como cada hebra sobrecruzada se realinea (reanneals) con su fibra compañera original, desplaza la fibra complementaria original hacia delante, haciendo que la unión de Holliday migre, creando los segmentos heterodúplex.
[4] El modelo original de Holliday asumía que el ADN heterodúplex estaría presente en ambos cromosomas pero los datos experimentales obtenidos en levaduras refutaron esta idea, lo que llevó a que el modelo fuera puesto al día en 1975 por Matt Meselson y Charley Radding, que introdujeron la idea de la migración de la rama.
[2] Pero los análisis químicos hechos en la década de 1980 mostraron que, en realidad, la unión prefería las conformaciones antiparalelas, lo que fue un descubrimiento considerado controvertido, y el propio Robin Holliday dudaba inicialmente de él.