Canal de calcio tipo L
También se ha visto evidencia que desempeñan un papel en la expresión génica, la estabilidad del ARNm, la supervivencia neuronal, la lesión axonal inducida por isquemia, la eficacia sináptica y la activación y desactivación de otros canales iónicos.
[4] La investigación sobre las subunidades CaV1 continúa revelando más sobre su estructura, función y aplicaciones farmacéuticas.
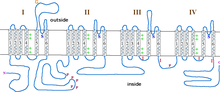
[10] La subunidad α1 del canal es un heterotetrámero con cuatro regiones transmembrana,[12] conocidas como Dominios I-IV, que atraviesan el la membrana plasmática seis veces como hélices α, denominándose S0-S6 (S0 y S1 juntas cruzan la membrana una vez).
Cada subunidad está formada por 6 dominios alfa-helicoidales transmembrana que atraviesan la membrana (numerados S1-S6).
Son los canales de calcio más comunes en el músculo cardíaco.
La apertura del poro en los canales de calcio tipo L tiene lugar en la subunidad α1.
Los paquetes hidrofóbicos en el complejo Ca2+/Cam también se unirá a tres secciones del dominio IQ conocidas como "anclas aromáticas".
El verapamilo o diltiazem se utilizan en clínica como antiarrítmicos ya que actúan principalmente sobre las fibras musculares miocárdicas.
[4] Una forma bien observada de modulación se debe al empalme alternativo.
Tiene una hélice α cargada positivamente en el terminal C llamada DCRD y una hélice cargada negativamente justo después del motivo IQ (sitio de interacción CaM) llamado PCRD.
Las dos hélices pueden formar una estructura que se une competitivamente con CaM para reducir la probabilidad de estado abierto y disminuir la inhibición dependiente del calcio (CDI).
Esto se debe al hecho de que las subunidades β aumentan la probabilidad de apertura del canal, la actividad en la membrana plasmática y antagonizan la ubiquitinación del canal.
[8] Los canales de calcio tipo L también son modulados por receptores acoplados a proteínas G y el sistema nervioso adrenérgico.