Los ungulados ( / ˈʌŋɡjʊl eɪts , -ɡjə- , -lɪts , -ləts / UNG - gyuu -layts, -gyə-, -lits, -ləts ) son miembros del clado Euungulata ( " ungulados verdaderos " ), que consiste principalmente en grandes mamíferos con pezuñas. Una vez parte del clado "Ungulata" junto con el clado Paenungulata, "Ungulata" ha sido determinado desde entonces como un clado polifilético y por lo tanto inválido basado en datos moleculares. Como resultado, los ungulados verdaderos han sido reclasificados desde entonces al clado más nuevo Euungulata en 2001 dentro del clado Laurasiatheria mientras que Paenungulata ha sido reclasificado a un clado distante Afrotheria . [4] Los ungulados vivos se dividen en dos órdenes: Perissodactyla que incluye equinos , rinocerontes y tapires ; y Artiodactyla que incluye ganado , antílopes , cerdos , jirafas , camellos , ovejas , ciervos e hipopótamos , entre otros. Los cetáceos como las ballenas , los delfines y las marsopas también se clasifican como artiodáctilos, aunque no tienen pezuñas. La mayoría de los ungulados terrestres usan las puntas unguladas de sus dedos para soportar su peso corporal mientras están de pie o en movimiento. Otros dos órdenes de ungulados, Notoungulata y Litopterna , ambos nativos de América del Sur, se extinguieron a fines del Pleistoceno, hace alrededor de 12 000 años.
El término significa, aproximadamente, "ser ungulado" o "animal ungulado". Como término descriptivo, "ungulado" normalmente excluye a los cetáceos ya que no poseen la mayoría de las características morfológicas típicas de otros ungulados, pero descubrimientos recientes indican que también descendían de los primeros artiodáctilos . [5] Los ungulados son típicamente herbívoros y muchos emplean bacterias intestinales especializadas para poder digerir la celulosa, aunque algunos miembros pueden desviarse de esto: varias especies de cerdos y los entelodontos extintos son omnívoros, mientras que los cetáceos y los mesoniquios extintos son carnívoros.
Ungulado proviene del adjetivo del latín tardío ungulatus, que significa " con pezuña " . Ungulatus es una forma diminutiva del latín unguis , que significa " uña " (uña de la mano; uña del pie). [6]
Euungulata es un clado (o en algunas taxonomías, un gran orden) de mamíferos. Los dos órdenes actuales de ungulados son Perissodactyla (ungulados de dedos impares) y Artiodactyla (ungulados de dedos pares) . Hyracoidea (damánes) , Sirenia (vacas marinas) (dugongos y manatíes) y Proboscidea (elefantes) se agruparon en el pasado dentro del clado "Ungulata", que luego se descubrió que era un clado polifilético y ahora inválido. Los tres órdenes de Paenungulata ahora se consideran un clado y se agrupan en el clado Afrotheria, mientras que Euungulata ahora se agrupa bajo el clado Laurasiatheria . [1]
En 2009, trabajos morfológicos [7] [8] [9] [10] y moleculares [11] [12] descubrieron que los cerdos hormigueros, los damanes, las vacas marinas y los elefantes estaban más estrechamente relacionados entre sí y con los sengis , los tenrecs y los topos dorados que con los perisodáctilos y los artiodáctilos, y forman el clado Afrotheria . Los elefantes, las vacas marinas y los damanes se agruparon en el clado Paenungulata , mientras que el cerdo hormiguero se ha considerado un pariente cercano de ellos o un pariente cercano de los sengis en el clado Afroinsectiphilia . [13] Este es un ejemplo sorprendente de evolución convergente . [14]
Actualmente existe cierta controversia sobre si este Euungulata más pequeño es un grupo cladístico (basado en la evolución), o simplemente un grupo fenético ( taxón de forma ) o taxón popular (similar, pero no necesariamente relacionado). Algunos estudios han encontrado de hecho que los ungulados mesaxonianos y los ungulados paraxonianos forman un linaje monofilético, [15] [16] [17] estrechamente relacionado con los Ferae (los carnívoros y los pangolines ) [18] [19] en el clado Fereuungulata o con los murciélagos . [20] Otros estudios encontraron que los dos órdenes no estaban tan estrechamente relacionados, ya que algunos ubican a los perisodáctilos como parientes cercanos de los murciélagos y a los Ferae en Pegasoferae [21] y otros ubican a los artiodáctilos como parientes cercanos de los murciélagos. [22]






A continuación se presenta una taxonomía simplificada (suponiendo que los ungulados forman efectivamente una agrupación natural) con las familias actuales, ordenadas según sus relaciones de parentesco. Tenga en cuenta que todavía había algunas áreas grises de conflicto, como el caso de la relación entre las familias de los pecoranos y las familias de las ballenas barbadas . Consulte cada familia para conocer las relaciones de las especies, así como las controversias en sus respectivos artículos.
A continuación se presenta el consenso general de la filogenia de las familias de ungulados. [22] [23]


Perissodactyla y Artiodactyla incluyen la mayoría de los grandes mamíferos terrestres. Estos dos grupos aparecieron por primera vez durante el Paleoceno tardío , extendiéndose rápidamente a una amplia variedad de especies en numerosos continentes, y se han desarrollado en paralelo desde entonces. Algunos científicos creían que los ungulados modernos descendían de un grado evolutivo de mamíferos conocidos como condilartos . [24] El primer miembro conocido de este grupo puede haber sido el diminuto Protungulatum , un mamífero que coexistió con el último de los dinosaurios no aviares hace 66 millones de años. [25] Sin embargo, muchas autoridades no lo consideran un verdadero placentario, y mucho menos un ungulado. [26] Los enigmáticos dinoceratans estuvieron entre los primeros grandes mamíferos herbívoros, aunque su relación exacta con otros mamíferos aún se debate y una de las teorías es que podrían ser simplemente parientes lejanos de los ungulados actuales; El estudio más reciente los recupera como parte del verdadero conjunto de ungulados, más cercano a Carodnia . [27]
En Australia, el marsupial recientemente extinto Chaeropus ("bandicut de patas de cerdo") también desarrolló pezuñas similares a las de los artiodáctilos, [28] un ejemplo de evolución convergente .

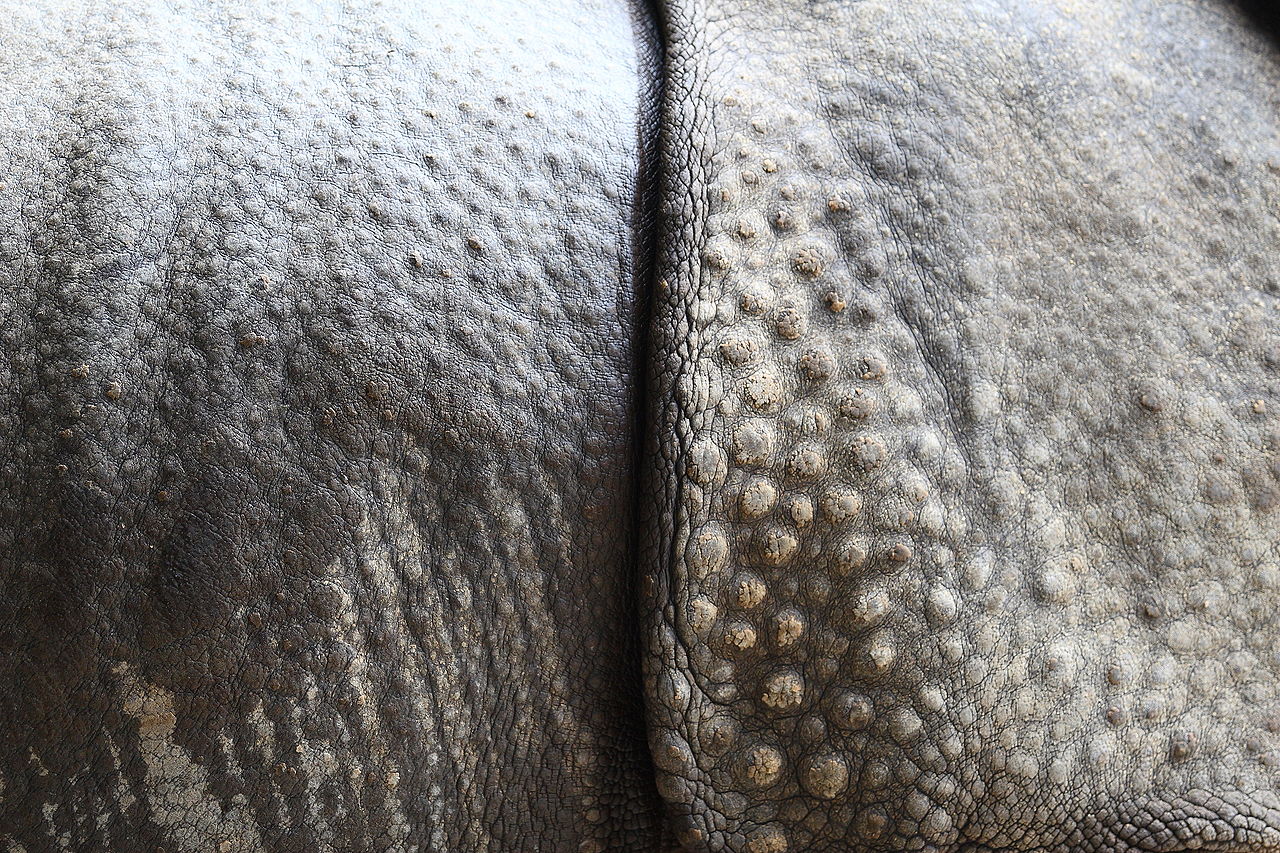
Se pensaba que los perisodáctilos habían evolucionado a partir de los fenacodóntidos , animales pequeños del tamaño de ovejas que ya mostraban signos de características anatómicas que sus descendientes heredarían (la reducción de los dedos I y V, por ejemplo). [30] A principios del Eoceno , hace 55 millones de años (Ma), se habían diversificado y extendido para ocupar varios continentes. Los caballos y los tapires evolucionaron en América del Norte; [31] los rinocerontes parecen haberse desarrollado en Asia a partir de animales similares a los tapires y luego colonizaron las Américas durante el Eoceno medio (alrededor de 45 Ma). De las aproximadamente 15 familias, solo tres sobreviven (McKenna y Bell, 1997; Hooker, 2005). Estas familias eran muy diversas en forma y tamaño; incluían a los enormes brontoterios y a los extraños calicoterios . El perisodáctilo más grande, un rinoceronte asiático llamado Paraceratherium , alcanzó las 15 toneladas (17 toneladas), más del doble del peso de un elefante . [32]
Se ha descubierto en un estudio cladístico que los antracobúnidos y los desmostilios (dos linajes que anteriormente se habían clasificado como afroterios (más específicamente más cercanos a los elefantes)) han sido clasificados como un clado estrechamente relacionado con los perisodáctilos. [2] Los desmostilios eran grandes cuadrúpedos anfibios con extremidades enormes y una cola corta. [33] [ cita larga faltante ] Crecieron hasta 1,8 metros (6 pies) de longitud y se cree que pesaron más de 200 kilogramos (440 lb). Sus fósiles se conocían desde la cuenca del Pacífico norte , [34] desde el sur de Japón a través de Rusia , las Islas Aleutianas y la costa del Pacífico de América del Norte hasta el extremo sur de Baja California . Su forma dental y esquelética sugiere que los desmostilios eran herbívoros acuáticos dependientes de los hábitats litorales . Su nombre hace referencia a sus característicos molares, en los que cada cúspide estaba modificada en columnas huecas, de modo que un molar típico se habría parecido a un conjunto de tubos o, en el caso de los molares desgastados, a volcanes. Fueron los únicos mamíferos marinos que se extinguieron.
Los meridiungulados sudamericanos contienen los piroteres y astrapotheres , que son algo parecidos a los tapires , los litopternos mesaxónicos y los diversos notoungulados . En conjunto, se decía que los meridiungulados habían evolucionado a partir de animales como Hyopsodus . [30] Durante un tiempo, sus relaciones con otros ungulados fueron un misterio. Algunos paleontólogos incluso han desafiado la monofilia de Meridiungulata al sugerir que los piroteres pueden estar más estrechamente relacionados con otros mamíferos, como Embrithopoda (un orden africano que estaba relacionado con los elefantes ) que con otros ungulados sudamericanos. [35] Un estudio reciente basado en el colágeno óseo ha descubierto que al menos los litopternos y los notoungulados estaban estrechamente relacionados con los perisodáctilos. [36]
Los fósiles más antiguos conocidos asignados a Equidae datan del Eoceno temprano , hace 54 millones de años. Habían sido asignados al género Hyracotherium , pero la especie tipo de ese género ahora se considera que no es miembro de esta familia, pero las otras especies se han dividido en diferentes géneros. Estos primeros équidos eran animales del tamaño de un zorro con tres dedos en las patas traseras y cuatro en las delanteras. Eran herbívoros que se alimentaban de plantas relativamente blandas y ya estaban adaptados para correr. La complejidad de sus cerebros sugiere que ya eran animales alertas e inteligentes. [37] Las especies posteriores redujeron el número de dedos y desarrollaron dientes más adecuados para triturar hierba y otros alimentos vegetales duros.
Los rinocerotóideos se separaron de otros perisodáctilos a principios del Eoceno. Los fósiles de Hyrachyus eximus encontrados en América del Norte datan de este período. Este pequeño antepasado sin cuernos se parecía más a un tapir o a un caballo pequeño que a un rinoceronte. Tres familias, a veces agrupadas como la superfamilia Rhinocerotoidea, evolucionaron a finales del Eoceno: Hyracodontidae , Amynodontidae y Rhinocerotidae , creando así una explosión de diversidad sin igual durante un tiempo hasta que los cambios ambientales eliminaron drásticamente varias especies.
Los primeros tapires, como Heptodon , aparecieron a principios del Eoceno. [38] Parecían muy similares a las formas modernas, pero tenían aproximadamente la mitad del tamaño y carecían de probóscide. Los primeros tapires verdaderos aparecieron en el Oligoceno . Para el Mioceno , géneros como Miotapirus eran casi indistinguibles de las especies actuales. Se cree que los tapires asiáticos y americanos divergieron hace unos 20 a 30 millones de años; y los tapires migraron de América del Norte a América del Sur hace unos 3 millones de años, como parte del Gran Intercambio Americano . [39]
Los perisodáctilos fueron el grupo dominante de grandes animales ramoneadores terrestres durante todo el Oligoceno. Sin embargo, el auge de las gramíneas en el Mioceno (hace unos 20 millones de años) supuso un cambio importante: las especies de artiodáctilos, con sus estómagos más complejos, pudieron adaptarse mejor a una dieta basta y poco nutritiva, y pronto cobraron importancia. No obstante, muchas especies de perisodáctilos sobrevivieron y prosperaron hasta finales del Pleistoceno (hace unos 10.000 años), cuando se enfrentaron a la presión de la caza humana y el cambio de hábitat.

Se pensaba que los artiodáctilos habían evolucionado a partir de un pequeño grupo de condilartos, Arctocyonidae , que eran omnívoros no especializados, superficialmente parecidos a los mapaches u osos del Paleoceno temprano (hace unos 65 a 60 millones de años). Tenían extremidades relativamente cortas que carecían de especializaciones asociadas con sus parientes (por ejemplo, dígitos laterales reducidos, huesos fusionados y pezuñas), [30] y colas largas y pesadas. Su anatomía primitiva hace poco probable que pudieran perseguir a sus presas, pero con sus poderosas proporciones, garras y caninos largos, pueden haber sido capaces de dominar a animales más pequeños en ataques sorpresa. [30] Evidentemente, estos mamíferos pronto evolucionaron en dos linajes separados: los mesoniquios y los artiodáctilos.

Los primeros artiodáctilos se parecían a los chevrotains o cerdos actuales : criaturas pequeñas de patas cortas que comían hojas y partes blandas de las plantas . A finales del Eoceno (hace 46 millones de años), ya se habían desarrollado los tres subórdenes modernos: Suina (el grupo de los cerdos ); Tylopoda (el grupo de los camellos ); y Ruminantia (el grupo de las cabras y el ganado vacuno ). Sin embargo, los artiodáctilos estaban lejos de ser dominantes en ese momento: los perisodáctilos tuvieron mucho más éxito y fueron mucho más numerosos. Los artiodáctilos sobrevivieron en roles de nicho, generalmente ocupando hábitats marginales , y es presumiblemente en ese momento que desarrollaron sus complejos sistemas digestivos , lo que les permitió sobrevivir con alimentos de menor calidad. Mientras que la mayoría de los artiodáctilos estaban tomando posesión de los nichos dejados por varios perisodáctilos extintos, un linaje de artiodáctilos comenzó a aventurarse en los mares.

La teoría tradicional de la evolución de los cetáceos era que los cetáceos estaban relacionados con los mesoniquios . Estos animales tenían dientes triangulares inusuales muy similares a los de los cetáceos primitivos. Es por eso que los científicos creyeron durante mucho tiempo que los cetáceos evolucionaron a partir de una forma de mesoniquio. Hoy, muchos científicos creen que los cetáceos evolucionaron a partir de la misma cepa que dio origen a los hipopótamos. Este hipotético grupo ancestral probablemente se dividió en dos ramas hace unos 54 millones de años . [5] Una rama evolucionaría en cetáceos , posiblemente comenzando hace unos 52 millones de años con la protoballena Pakicetus y otros ancestros cetáceos tempranos conocidos colectivamente como Archaeoceti , que finalmente experimentaron una adaptación acuática en los cetáceos completamente acuáticos . [40] La otra rama se convirtió en los antracoterios , una gran familia de bestias de cuatro patas, la primera de las cuales, a finales del Eoceno, se habría parecido a hipopótamos flacos con cabezas comparativamente pequeñas y estrechas. Todas las ramas de los antracoterios, excepto la que evolucionó a Hippopotamidae , se extinguieron durante el Plioceno sin dejar descendientes. [41]
Se dice que la familia Raoellidae es la familia de artiodáctilos más cercana a los cetáceos. [42] [43] En consecuencia, las nuevas teorías sobre la evolución de los cetáceos plantean la hipótesis de que las ballenas y sus ancestros escaparon de la depredación, no de la competencia, al adaptarse lentamente al océano. [44] [45] [46]

Los mesoniquios fueron representados como "lobos con pezuñas" y fueron los primeros depredadores mamíferos importantes, apareciendo en el Paleoceno. [47] Los primeros mesoniquios tenían cinco dedos en sus pies, que probablemente descansaban planos sobre el suelo durante la marcha ( locomoción plantígrada ), pero los mesoniquios posteriores tenían cuatro dedos que terminaban en pequeñas pezuñas en todos sus dedos y estaban cada vez mejor adaptados a correr. Al igual que los miembros corredores de los ungulados de dedos pares, los mesoniquios ( Pachyaena , por ejemplo) caminaban sobre sus dedos ( locomoción digitígrada ). [47] A los mesoniquios les fue muy mal al final de la época del Eoceno, con solo un género, Mongolestes , [48] sobreviviendo hasta la época del Oligoceno temprano , a medida que el clima cambiaba y surgía una feroz competencia de los creodontos mejor adaptados .

Los ungulados presentaban una gran diversidad en respuesta a la selección sexual y a los acontecimientos ecológicos ; la mayoría de los ungulados carecen de clavícula . [49] Los ungulados terrestres eran en su mayoría herbívoros, y algunos de ellos pastaban . Sin embargo, había excepciones a esto, ya que se sabía que los cerdos, los pecaríes, los hipopótamos y los duikers tenían una dieta omnívora. Algunos cetáceos eran los únicos ungulados modernos que eran carnívoros; las ballenas barbadas consumen animales significativamente más pequeños en relación con su tamaño corporal, como pequeñas especies de peces y krill ; las ballenas dentadas, dependiendo de la especie, pueden consumir una amplia gama de especies: calamares , peces, tiburones y otras especies de mamíferos como focas y otras ballenas. En términos de ecosistema, los ungulados han colonizado todos los rincones del planeta, desde las montañas hasta las profundidades del océano ; desde las praderas hasta los desiertos y algunos han sido domesticados por los humanos .
Los ungulados han desarrollado adaptaciones especializadas, especialmente en las áreas de apéndices craneales, dentición y morfología de las patas, incluida la modificación del astrágalo (uno de los huesos del tobillo al final de la parte inferior de la pierna) con una cabeza corta y robusta.
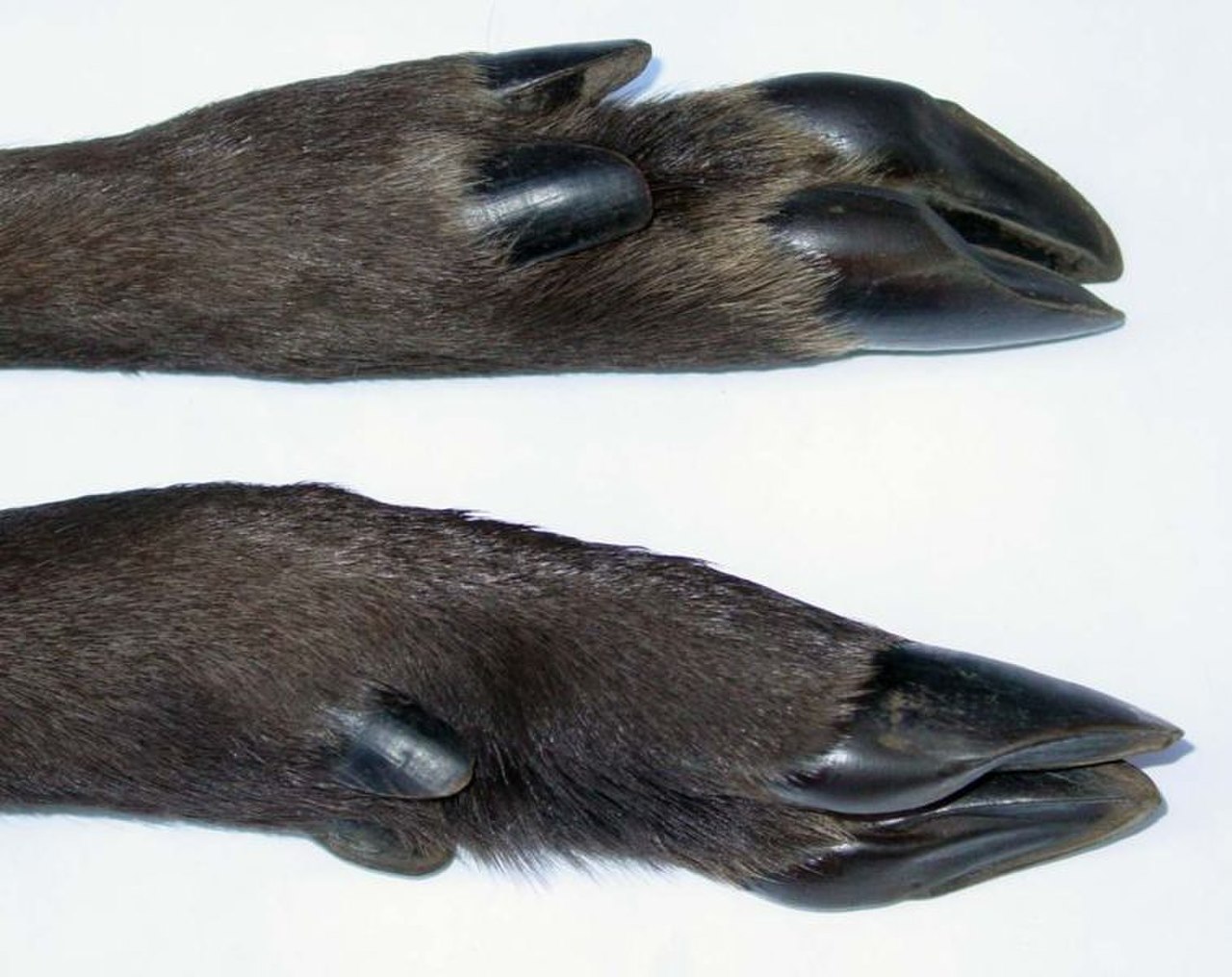
La pezuña es la punta del dedo del pie de un mamífero ungulado , reforzada por una gruesa cubierta córnea ( queratina ). La pezuña consta de una suela dura o gomosa y una pared dura formada por una uña gruesa enrollada alrededor de la punta del dedo. Tanto la suela como el borde de la pared del casco normalmente soportan el peso del animal. Las pezuñas crecen continuamente y se desgastan constantemente por el uso. En la mayoría de los ungulados modernos, el radio y el cúbito están fusionados a lo largo de la extremidad anterior; los ungulados primitivos, como los arctociónidos , no compartían esta estructura esquelética única. [50] La fusión del radio y el cúbito evita que un ungulado gire su extremidad anterior. Dado que esta estructura esquelética no tiene una función específica en los ungulados, se considera una característica homóloga que los ungulados comparten con otros mamíferos. Este rasgo se habría transmitido de un ancestro común. Aunque los nombres coloquiales de los dos órdenes de ungulados se basaban en el número de dedos de sus miembros ("dedos impares" para los perisodáctilos y "dedos pares" para los artiodáctilos terrestres), no es una razón precisa para agruparlos. Los tapires tienen cuatro dedos en la parte delantera, pero eran miembros del orden de los "dedos impares"; los pecaríes y los cetáceos modernos eran miembros del orden de los "dedos pares", pero los pecaríes tienen tres dedos en la parte delantera y las ballenas eran un ejemplo extremo, ya que tienen aletas en lugar de pezuñas. Los científicos los habían clasificado según la distribución de su peso en los dedos.
Los perisodáctilos tienen un pie mesaxónico, lo que significa que el peso se distribuye en el tercer dedo de todas las patas gracias a la simetría plana de sus pies. Se ha producido una reducción de los dedos a partir del ancestro común, siendo el ejemplo clásico los caballos con sus pezuñas únicas. En consecuencia, hubo un nombre alternativo para los perisodáctilos, el casi obsoleto Mesaxonia. Los perisodáctilos no fueron el único linaje de mamíferos que desarrolló este rasgo; los meridiungulados han desarrollado pies mesaxónicos en numerosas ocasiones.
Los artiodáctilos terrestres tienen un pie paraxónico, lo que significa que el peso se distribuye en el tercer y cuarto dedo de todas las patas. La mayoría de estos mamíferos tienen pezuñas hendidas, con dos pezuñas más pequeñas conocidas como espolones que se ubicaban más arriba en la pata. Los primeros cetáceos (los arqueocetos ) también tenían esta característica, además de tener un astrágalo y un hueso cuboides en el tobillo, que eran otros rasgos diagnósticos de los artiodáctilos. [51]

En los cetáceos modernos, las extremidades anteriores se habían convertido en aletas pectorales y las partes posteriores eran internas y reducidas. Ocasionalmente, los genes que codifican extremidades más largas hacen que un cetáceo moderno desarrolle patas en miniatura (conocido como atavismo ). El principal método de movimiento es un movimiento de arriba a abajo con la aleta caudal, llamada aleta caudal, que se utiliza para la propulsión , mientras que las aletas pectorales junto con toda la sección de la cola proporcionan control direccional. Todos los cetáceos modernos aún conservan sus dedos a pesar de que la apariencia externa sugiera lo contrario.
La mayoría de los ungulados han desarrollado dientes caninos reducidos y molares especializados , incluyendo dientes bunodontes (cúspides bajas y redondeadas) e hipsodontes (corona alta). El desarrollo de la hipsodoncia ha sido de particular interés ya que esta adaptación estuvo fuertemente asociada con la expansión de los pastizales durante el Mioceno hace unos 25 millones de años. A medida que los biomas forestales declinaron, los pastizales se expandieron, abriendo nuevos nichos para los mamíferos. Muchos ungulados cambiaron de dietas de ramoneo a dietas de pastoreo, y posiblemente impulsados por la sílice abrasiva en la hierba, la hipsodoncia se volvió común. Sin embargo, evidencia reciente vincula la evolución de la hipsodoncia a hábitats abiertos y arenosos y no a la hierba en sí. Esto se denomina la hipótesis de la arenilla, no de la hierba . [52]
Algunos ungulados carecen por completo de incisivos superiores y en su lugar tienen una almohadilla dental para facilitar la alimentación. [53] [54] Se puede encontrar en camellos, rumiantes y algunas ballenas dentadas; las ballenas barbadas modernas eran notables porque tenían barbas en su lugar para filtrar el krill del agua. En el otro espectro, los dientes se han desarrollado como armas o exhibición sexual, como se ve en cerdos y pecaríes, algunas especies de ciervos, ciervos almizcleros, hipopótamos, ballenas picudas y el narval, con su largo diente canino. [55]

Los ungulados han desarrollado una variedad de apéndices craneales que se pueden encontrar en los cérvidos (con la excepción del ciervo almizclero). En los bueyes y los antílopes, el tamaño y la forma de los cuernos varían mucho, pero la estructura básica es siempre un par de protuberancias óseas simples sin ramificaciones, que a menudo tienen una forma espiral, retorcida o estriada, cada una cubierta por una vaina permanente de queratina . La estructura única del cuerno es la única característica morfológica inequívoca de los bóvidos que los distingue de otros pecoranos . [56] [57] El desarrollo de los cuernos masculinos se ha relacionado con la selección sexual, [58] [59] mientras que la presencia de cuernos en las hembras probablemente se deba a la selección natural. [58] [60] Los cuernos de las hembras suelen ser más pequeños que los de los machos y, a veces, tienen una forma diferente. Se cree que los cuernos de los bóvidos hembras evolucionaron para defenderse de los depredadores o para expresar territorialidad, ya que las hembras no territoriales, que pueden usar la cripsis para defenderse de los depredadores, a menudo carecen de cuernos. [60]
Los cuernos del rinoceronte, a diferencia de los de otros mamíferos con cuernos, están compuestos únicamente de queratina. Estos cuernos descansan sobre la cresta nasal del cráneo del animal.
Las astas son exclusivas de los cérvidos y se encuentran principalmente en los machos: las únicas hembras de cérvidos con astas son las del caribú y el reno , cuyas astas son normalmente más pequeñas que las de los machos. Sin embargo, las hembras fértiles de otras especies de ciervos tienen la capacidad de producir astas en ocasiones, generalmente debido al aumento de los niveles de testosterona. [61] Cada asta crece desde un punto de unión en el cráneo llamado pedículo. Mientras una asta está creciendo, está cubierta de una piel altamente vascularizada llamada terciopelo, que suministra oxígeno y nutrientes al hueso en crecimiento. [62] Las astas se consideran uno de los casos más exagerados de rasgos sexuales secundarios masculinos en el reino animal, [63] y crecen más rápido que cualquier otro hueso de mamífero. [64] El crecimiento ocurre en la punta, inicialmente como cartílago que luego se mineraliza para convertirse en hueso. Una vez que la asta ha alcanzado su tamaño completo, el terciopelo se pierde y el hueso de la asta muere. Esta estructura ósea muerta es la asta madura. En la mayoría de los casos, el hueso de la base es destruido por los osteoclastos y las astas finalmente se caen. [62] Como resultado de su rápida tasa de crecimiento, las astas suponen una demanda nutricional sustancial para los ciervos; por lo tanto, pueden constituir una señal honesta de eficiencia metabólica y capacidad de recolección de alimentos. [65]
Los osiconos son protuberancias similares a cuernos (o astas) que se encuentran en las cabezas de las jirafas y los okapis machos . Son similares a los cuernos de los antílopes y el ganado vacuno , salvo que derivan de cartílago osificado [66] y que los osiconos permanecen cubiertos de piel y pelo en lugar de cuerno.
Los apéndices craneales del berrendo son únicos. Cada "cuerno" del berrendo está compuesto por una lámina ósea delgada y aplanada lateralmente que crece desde los huesos frontales del cráneo y forma un núcleo permanente. Al igual que en los Jiráfidos, la piel cubre los núcleos óseos, pero en el berrendo se desarrolla en una vaina queratinosa que se desprende y vuelve a crecer anualmente. A diferencia de los cuernos de la familia Bovidae, las vainas córneas del berrendo están ramificadas y cada vaina posee una púa que apunta hacia adelante (de ahí el nombre de berrendo). Los cuernos de los machos están bien desarrollados.
{{cite book}}: CS1 maint: varios nombres: lista de autores ( enlace ){{cite journal}}: CS1 maint: varios nombres: lista de autores ( enlace )