Micro-ARN
[6] Una vez descubiertos los siARN y la existencia en las células de proteínas que catalizan la degradación del mARN, los investigadores se preguntaron si los siARN también estaban codificados en el genoma, y empezaron a purificar pequeños ARN (19-25 nt) a partir de diferentes especies animales.
Muchos micro-ARN están bien conservados entre especies,[8] y muchos componentes de la maquinaria de los micro-ARN se han encontrado incluso en arqueas y bacterias, lo que revela que su origen es muy antiguo.
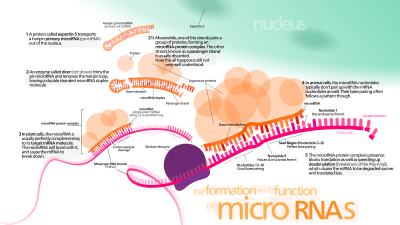
Posteriormente, una enzima nuclear llamada DROSHA corta las bases de la horquilla, formando lo que se denomina pre-micro-ARN.
Los micro-ARN de animales suelen mostrar complementariedad imperfecta con la región 3' UTR, y generalmente inhiben la traducción del mARN, mientras que los de plantas suelen mostrar complementariedad perfecta con regiones codificantes e inducen el corte y la posterior degradación del mARN diana (como ocurre con los siARNs en animales).
[12] Según esto, cualquier desregulación de los micro-ARN podría conllevar grandes problemas regulatorios en la célula, induciendo quizá fenotipos cancerosos.
[14] Los micro-ARN se transcriben a partir de diferentes localizaciones genómicas como largos tránscritos primarios (pri-micro-ARN) por la ARN-polimerasa II.
Los genes que codifican micro-ARN son mucho más largos que los micro-ARN procesados maduros; los micro-ARN se transcriben inicialmente como tránscritos primarios o pri-micro-ARN con una caperuza en 5' y una cola de poli-adeninas (poly-A) en 3' y se procesan en el núcleo celular en estructuras cortas de 70-nucleótidos en forma de tallo-asa (stem-loop) conocidas como pre-micro-ARN.
[20][21] Estos pre-micro-ARN son luego procesados a micro-ARN maduros en el citoplasma mediante la interacción con la ribonucleasa DICER, que también inicia la formación del complejo RISC (ARN-induced silencing complex).
[25] DROSHA[26] (ARNSEN en humanos)[19] es una proteína nuclear de un tamaño entre 130 y 160 kDa (kiloDalton).
Contiene los dominios siguientes: DROSHA funciona en un complejo (Microprocesador), conjuntamente con una proteína de unión a ARN (denominada PASHA en Drosophila o DGCR8 en mamíferos).
Un análisis bioinformático de pri-micro-ARN en humanos y moscas identificó regiones estructurales similares, denominadas 'segmentos basales (basal segments)', 'tallos inferiores (lower stems)', 'tallos superiores (upper stems)' y 'asas terminales (terminal loops)'; basándose en estas estructuras conservadas, se han determinado perfiles termodinámicos de los pri-micro-ARN.
En la mayoría de las moléculas analizadas esta región contiene nucleótidos no apareados y la energía libre del dúplex es relativamente alta en comparación con las regiones tallo superior e inferior [cita requerida].
De hecho, se han detectado claras similitudes entre pri-micro-ARN codificados bien en hebras 5' o 3'.
Las proteínas Argonauta, los componentes catalíticos de RISC, están localizadas en regiones específicas del citoplasma denominadas P-bodies (cuerpos-P, o cuerpos citoplásmicos o cuerpos GW, porque contienen la proteína GW182), los cuales son regiones con altas tasas de degradación de mARN;[36] también se ha detectado actividad micro-ARN en los P-bodies.
[38] Sin embargo, estudios posteriores han demostrado que los P-bodies no son imprescindibles para el proceso de iARN, ya que células sin P-bodies pueden producir silenciamiento tanto con siARN como con micro-ARN.
[42] Por último, en estudios recientes se ha detectado que en determinadas condiciones, los micro-ARN pueden también activar la síntesis proteica.
Por otro lado, se ha observado además que los micro-ARN contribuyen a la progresión maligna del cáncer, específicamente mediando la invasión tumoral y la formación de metástasis.
[57] Por ejemplo, en C. elegans, los micro-ARN permiten un paso rápido a través de las diferentes fases del desarrollo:[58] los micro-ARN lin-4[59] y let-7[60] controlan el momento en los que se define el destino de las células neuronales e hipodérmicas durante el desarrollo larvario.
[82] Dos estudios independientes en 2007 en ratones[83][84] indican que una cantidad significativa de micro-ARN maternos se heredan por los zigotos, y que dichos micro-ARN maternos podrían tener un papel importante en las etapas tempranas del desarrollo embrionario (efecto materno).
Estudios de asociación de genoma completo (GWAS) se han empleado para identificar ciertos micro-ARNs que modulando la traducción del mARN funcionan como reguladores cruciales del colesterol, triglicéridos y homeostasis energética en el organismo.
Debido a que se necesita ABCA1 para su síntesis, y este ha disminuido con la introducción de los micro-ARNs.
Todo esto se revierte al introducir anti-micro-ARNs, por tanto, estos podrían ser aplicados en el tratamiento de enfermedades relacionadas con la homeostasis, sensibilidad glucídica, señalización de la insulina, triglicéridos y colesterol en sangre.