Los Plesiosauria [a] [4] o plesiosaurios son un orden o clado de reptiles marinos mesozoicos extintos , pertenecientes a los Sauropterigia .
Los plesiosaurios aparecieron por primera vez en el último período Triásico , posiblemente en la etapa Rético , hace unos 203 millones de años. [5] Se volvieron especialmente comunes durante el período Jurásico , prosperando hasta su desaparición debido al evento de extinción masiva del Cretácico-Paleógeno al final del período Cretácico , hace unos 66 millones de años. Tenían una distribución oceánica mundial y algunas especies habitaban al menos parcialmente ambientes de agua dulce. [6]
Los plesiosaurios fueron de los primeros reptiles fósiles descubiertos. A principios del siglo XIX, los científicos se dieron cuenta de lo peculiar que era su constitución y se les dio un nombre como orden independiente en 1835. El primer género de plesiosaurio, el epónimo Plesiosaurus , recibió su nombre en 1821. Desde entonces, se han descrito más de cien especies válidas. A principios del siglo XXI, el número de descubrimientos ha aumentado, lo que ha llevado a una mejor comprensión de su anatomía, sus relaciones y su forma de vida.
Los plesiosaurios tenían un cuerpo ancho y plano y una cola corta. Sus extremidades habían evolucionado hasta convertirse en cuatro aletas largas, que eran impulsadas por músculos fuertes unidos a placas óseas anchas formadas por la cintura escapular y la pelvis. Las aletas realizaban un movimiento de vuelo a través del agua. Los plesiosaurios respiraban aire y tenían crías vivas; hay indicios de que eran de sangre caliente.
Los plesiosaurios mostraron dos tipos morfológicos principales . Algunas especies, con la constitución "plesiosauromorfa", tenían cuellos (a veces extremadamente) largos y cabezas pequeñas; eran relativamente lentos y capturaban pequeños animales marinos. Otras especies, algunas de las cuales alcanzaban una longitud de hasta diecisiete metros, tenían la constitución "pliosauromorfa" con un cuello corto y una cabeza grande; eran depredadores de ápice , cazadores rápidos de presas grandes. Los dos tipos están relacionados con la división estricta tradicional de los Plesiosauria en dos subórdenes, los Plesiosauroidea de cuello largo y los Pliosauroidea de cuello corto . Sin embargo, la investigación moderna indica que varios grupos "de cuello largo" podrían haber tenido algunos miembros de cuello corto o viceversa. Por lo tanto, se han introducido los términos puramente descriptivos "plesiosauromorfo" y "pliosauromorfo", que no implican una relación directa. "Plesiosauroidea" y "Pliosauroidea" hoy tienen un significado más limitado. El término "plesiosaurio" se utiliza propiamente para referirse a Plesiosauria en su conjunto, pero informalmente a veces se pretende indicar únicamente las formas de cuello largo, los antiguos Plesiosauroidea.

Los elementos esqueléticos de los plesiosaurios se encuentran entre los primeros fósiles de reptiles extintos reconocidos como tales. [7] En 1605, Richard Verstegen de Amberes ilustró en su A Restitution of Decayed Intelligence vértebras de plesiosaurio que él atribuía a peces y que consideró una prueba de que Gran Bretaña alguna vez estuvo conectada al continente europeo. [8] El galés Edward Lhuyd en su Lithophylacii Brittannici Ichnographia de 1699 también incluyó representaciones de vértebras de plesiosaurio que nuevamente se consideraron vértebras de peces o Ichthyospondyli . [9] Otros naturalistas durante el siglo XVII agregaron restos de plesiosaurio a sus colecciones, como John Woodward ; solo mucho más tarde se entendió que eran de naturaleza plesiosauriana y hoy se conservan parcialmente en el Museo Sedgwick . [7]
En 1719, William Stukeley describió un esqueleto parcial de un plesiosaurio, que había sido traído a su atención por el bisabuelo de Charles Darwin , Robert Darwin de Elston . La placa de piedra provenía de una cantera en Fulbeck en Lincolnshire y había sido utilizada, con el fósil en su parte inferior, para reforzar la pendiente de un abrevadero en Elston en Nottinghamshire . Después de que se descubrieran los extraños huesos que contenía, se exhibió en la vicaría local como los restos de un pecador ahogado en el Gran Diluvio . Stukeley afirmó su naturaleza " diluvial ", pero entendió que representaba alguna criatura marina, tal vez un cocodrilo o un delfín. [10] El espécimen está hoy en exhibición en el Museo de Historia Natural , y su número de inventario es NHMUK PV R.1330 (anteriormente BMNH R.1330). Es el esqueleto fósil de reptil más o menos completo descubierto más antiguo en una colección de museo. Tal vez pueda referirse a Plesiosaurus dolichodeirus . [7]

Durante el siglo XVIII, el número de descubrimientos ingleses de plesiosaurios aumentó rápidamente, aunque todos ellos fueron de naturaleza más o menos fragmentaria. Los coleccionistas importantes fueron los reverendos William Mounsey y Baptist Noel Turner, activos en el valle de Belvoir , cuyas colecciones fueron descritas en 1795 por John Nicholls en la primera parte de su Historia y antigüedades del condado de Leicestershire . [11] Uno de los esqueletos parciales de plesiosaurio de Turner todavía se conserva como espécimen NHMUK PV R.45 (anteriormente BMNH R.45) en el Museo Británico de Historia Natural; hoy se lo conoce como Thalassiodracon . [7]

A principios del siglo XIX, los plesiosaurios eran todavía poco conocidos y su constitución especial no se entendía. No se hizo una distinción sistemática con los ictiosaurios , por lo que los fósiles de un grupo a veces se combinaban con los del otro para obtener un espécimen más completo. En 1821, un esqueleto parcial descubierto en la colección del coronel Thomas James Birch, [12] fue descrito por William Conybeare y Henry Thomas De la Beche , y reconocido como representante de un grupo distintivo. Se nombró un nuevo género, Plesiosaurus . El nombre genérico se derivó del griego πλήσιος, plèsios , "más cercano a" y del latinizado saurus , en el sentido de "saurio", para expresar que Plesiosaurus estaba en la Cadena del Ser más cerca de los Sauria , particularmente el cocodrilo, que Ichthyosaurus , que tenía la forma de un pez más humilde. [13] Por lo tanto, el nombre debería leerse más como "que se acerca al Sauria" o "casi reptil" que como "casi lagarto". [14] Partes del espécimen todavía están presentes en el Museo de Historia Natural de la Universidad de Oxford . [7]
Poco después, la morfología se hizo mucho más conocida. En 1823, Thomas Clark informó de un cráneo casi completo, probablemente perteneciente a Thalassiodracon , que ahora está preservado por el British Geological Survey como espécimen BGS GSM 26035. [7] El mismo año, la coleccionista comercial de fósiles Mary Anning y su familia descubrieron un esqueleto casi completo en Lyme Regis en Dorset , Inglaterra, en lo que hoy se llama la Costa Jurásica . Fue adquirido por el duque de Buckingham , quien lo puso a disposición del geólogo William Buckland . Él a su vez dejó que Conybeare lo describiera el 24 de febrero de 1824 en una conferencia en la Sociedad Geológica de Londres , [15] durante la misma reunión en la que por primera vez se nombró a un dinosaurio, Megalosaurus . Los dos hallazgos revelaron la constitución única y extraña de los animales, en 1832 por el profesor Buckland comparado con "una serpiente marina atravesando una tortuga". En 1824, Conybeare también proporcionó un nombre específico a Plesiosaurus : dolichodeirus , que significa "cuello largo". En 1848, el esqueleto fue comprado por el Museo Británico de Historia Natural y catalogado como espécimen NHMUK OR 22656 (anteriormente BMNH 22656). [7] Cuando se publicó la conferencia, Conybeare también nombró una segunda especie: Plesiosaurus giganteus . Esta era una forma de cuello corto que luego se asignó a Pliosauroidea . [16]

Los plesiosaurios se hicieron más conocidos para el público en general a través de dos publicaciones profusamente ilustradas del coleccionista Thomas Hawkins : Memorias de Ichthyosauri y Plesiosauri de 1834 [17] y El libro de los grandes dragones marinos de 1840. Hawkins tenía una visión muy idiosincrásica de los animales, [18] viéndolos como creaciones monstruosas del diablo, durante una fase pre-adamítica de la historia. [19] Hawkins finalmente vendió sus valiosos y atractivamente restaurados especímenes al Museo Británico de Historia Natural. [20]
Durante la primera mitad del siglo XIX, el número de hallazgos de plesiosaurios aumentó de forma constante, especialmente a través de descubrimientos en los acantilados marinos de Lyme Regis. Sir Richard Owen solo nombró casi cien especies nuevas. La mayoría de sus descripciones, sin embargo, se basaban en huesos aislados, sin un diagnóstico suficiente para poder distinguirlas de las otras especies que se habían descrito previamente. Muchas de las nuevas especies descritas en esta época han sido posteriormente invalidadas . El género Plesiosaurus es particularmente problemático, ya que la mayoría de las nuevas especies se colocaron en él de modo que se convirtió en un taxón de basura . Poco a poco, se nombraron otros géneros. Hawkins ya había creado nuevos géneros, aunque estos ya no se consideran válidos. En 1841, Owen nombró a Pliosaurus brachydeirus . Su etimología hace referencia al anterior Plesiosaurus dolichodeirus , ya que deriva de πλεῖος, pleios , "más completamente", lo que refleja que según Owen estaba más cerca de Sauria que Plesiosaurus . Su nombre específico significa "con cuello corto". [21] Más tarde, se reconoció que los Pliosauridae tenían una morfología fundamentalmente diferente de los plesiosauridos. La familia Plesiosauridae ya había sido acuñada por John Edward Gray en 1825. [22] En 1835, Henri Marie Ducrotay de Blainville nombró al orden Plesiosauria. [23]
En la segunda mitad del siglo XIX se hicieron importantes descubrimientos fuera de Inglaterra. Si bien se trataba de algunos descubrimientos alemanes, se trataba principalmente de plesiosaurios encontrados en los sedimentos de la vía marítima interior occidental del Cretácico americano , la tiza de Niobrara . Un fósil en particular marcó el inicio de las Guerras de los Huesos entre los paleontólogos rivales Edward Drinker Cope y Othniel Charles Marsh .

En 1867, el médico Theophilus Turner descubrió cerca de Fort Wallace en Kansas un esqueleto de plesiosaurio, que donó a Cope. [24] Cope intentó reconstruir el animal suponiendo que la extremidad más larga de la columna vertebral era la cola, la más corta, el cuello. Pronto se dio cuenta de que el esqueleto que tomaba forma bajo sus manos tenía algunas cualidades muy especiales: las vértebras del cuello tenían cheurones y en las vértebras de la cola las superficies articulares estaban orientadas de atrás hacia adelante. [25] Emocionado, Cope concluyó que había descubierto un grupo completamente nuevo de reptiles: los Streptosauria o "saurios girados", que se distinguirían por las vértebras invertidas y la falta de extremidades traseras, siendo la cola la principal fuente de propulsión. [26] Después de haber publicado una descripción de este animal, [27] seguida de una ilustración en un libro de texto sobre reptiles y anfibios, [28] Cope invitó a Marsh y Joseph Leidy a admirar su nuevo Elasmosaurus platyurus . Tras escuchar la interpretación de Cope durante un rato, Marsh sugirió que una explicación más sencilla de la extraña constitución sería que Cope había invertido la columna vertebral en relación con el cuerpo en su conjunto. Cuando Cope reaccionó indignado a esta sugerencia, Leidy silenciosamente tomó el cráneo y lo colocó contra la presunta última vértebra de la cola, a la que encajaba perfectamente: era de hecho la primera vértebra del cuello, con todavía un trozo del cráneo trasero unido a ella. [29] Mortificado, Cope intentó destruir toda la edición del libro de texto y, cuando esto fracasó, publicó inmediatamente una edición mejorada con una ilustración correcta pero una fecha de publicación idéntica. [30] Disculpó su error afirmando que había sido engañado por el propio Leidy, quien, al describir un espécimen de Cimoliasaurus , también había invertido la columna vertebral. [31] Marsh afirmó más tarde que el asunto fue la causa de su rivalidad con Cope: "desde entonces ha sido mi acérrimo enemigo". Tanto Cope como Marsh en su rivalidad nombraron muchos géneros y especies de plesiosaurios, la mayoría de los cuales hoy se consideran inválidos. [32]
A principios de siglo, la mayor parte de la investigación sobre plesiosaurios la realizó un ex alumno de Marsh, el profesor Samuel Wendell Williston . En 1914, Williston publicó su libro Water reptiles of the past and present . [33] A pesar de tratar los reptiles marinos en general, durante muchos años seguiría siendo el texto general más extenso sobre plesiosaurios. [34] En 2013, Olivier Rieppel estaba preparando un primer libro de texto moderno. A mediados del siglo XX, Estados Unidos siguió siendo un importante centro de investigación, principalmente a través de los descubrimientos de Samuel Paul Welles .
Mientras que durante el siglo XIX y la mayor parte del XX se describían nuevos plesiosaurios a un ritmo de tres o cuatro géneros nuevos cada década, el ritmo se aceleró de repente en la década de 1990, con el descubrimiento de diecisiete plesiosaurios en este período. El ritmo de los descubrimientos se aceleró a principios del siglo XXI, con unos tres o cuatro plesiosaurios nombrados cada año. [35] Esto implica que aproximadamente la mitad de los plesiosaurios conocidos son relativamente nuevos para la ciencia, resultado de una investigación de campo mucho más intensa. Parte de esto se está llevando a cabo fuera de las áreas tradicionales, por ejemplo, en nuevos sitios desarrollados en Nueva Zelanda , Argentina , Chile , [36] Noruega , Japón , China y Marruecos , pero las ubicaciones de los descubrimientos más originales han demostrado ser todavía productivas, con nuevos hallazgos importantes en Inglaterra y Alemania. Algunos de los nuevos géneros son un cambio de nombre de especies ya conocidas, que se consideraron lo suficientemente diferentes como para justificar un nombre de género separado.
En 2002, se anunció a la prensa el « Monstruo de Aramberri ». Descubierto en 1982 en el pueblo de Aramberri , en el estado de Nuevo León , al norte de México , fue clasificado originalmente como un dinosaurio . El espécimen es en realidad un plesiosaurio muy grande, que posiblemente alcanzó los 15 m (49 pies) de longitud. Los medios publicaron informes exagerados que afirmaban que medía 25 metros (82 pies) de largo y pesaba hasta 150.000 kilogramos (330.000 libras), lo que lo habría convertido en uno de los depredadores más grandes de todos los tiempos. [37] [38]
En 2004, un pescador local descubrió en la Reserva Natural Nacional de la Bahía de Bridgwater , en Somerset (Reino Unido), lo que parecía ser un plesiosaurio juvenil completamente intacto. El fósil, que data de hace 180 millones de años, como indican los amonites asociados a él, medía 1,5 metros de largo y podría estar relacionado con el Rhomaleosaurus . Es probablemente el ejemplar mejor conservado de un plesiosaurio descubierto hasta ahora. [39] [40] [41]
En 2005, el Dr. Tamaki Sato, un paleontólogo de vertebrados japonés, determinó que los restos de tres plesiosaurios ( Dolichorhynchops herschelensis ) descubiertos en la década de 1990 cerca de Herschel, Saskatchewan, eran una nueva especie. [42]
En 2006, un equipo combinado de investigadores estadounidenses y argentinos (estos últimos del Instituto Antártico Argentino y el Museo de La Plata ) encontraron el esqueleto de un plesiosaurio juvenil que medía 1,5 metros (4 pies 11 pulgadas) de largo en la isla Vega en la Antártida. [43] El fósil se encuentra actualmente en exhibición en el museo geológico de la Escuela de Minas y Tecnología de Dakota del Sur . [44]
En 2008, se desenterraron en Svalbard restos fósiles de un plesiosaurio no descrito que fue llamado Predator X , ahora conocido como Pliosaurus funkei . [45] Tenía una longitud de 12 m (39 pies), y su fuerza de mordida de 149 kilonewtons (33.000 lb f ) es una de las más poderosas conocidas. [46]
En diciembre de 2017, se encontró un gran esqueleto de un plesiosaurio en el continente antártico, la criatura más antigua del continente y el primero de su especie en la Antártida. [47]
No sólo ha aumentado el número de descubrimientos de campo, sino que, además, desde la década de 1950, los plesiosaurios han sido objeto de un trabajo teórico más extenso. La metodología de la cladística ha permitido, por primera vez, el cálculo exacto de sus relaciones evolutivas. Se han publicado varias hipótesis sobre la forma en que cazaban y nadaban, incorporando conocimientos modernos generales sobre biomecánica y ecología . Los numerosos descubrimientos recientes han puesto a prueba estas hipótesis y han dado lugar a otras nuevas. [ ¿ Investigación original? ]

Los Plesiosauria tienen su origen en los Sauropterigia , un grupo de reptiles quizás arquelosaurios que regresaron al mar. Un subgrupo sauropterigio avanzado, los Eusauropterigia carnívoros con cabezas pequeñas y cuellos largos, se dividió en dos ramas durante el Triásico Superior . Una de ellas, los Nothosauroidea , mantuvo articulaciones funcionales en los codos y las rodillas; pero la otra, los Pistosauria , se adaptó más plenamente a un estilo de vida marino. Su columna vertebral se volvió más rígida y la propulsión principal mientras nadaban ya no provenía de la cola sino de las extremidades, que se transformaron en aletas. [48] Los Pistosauria se volvieron de sangre caliente y vivíparos , dando a luz a crías vivas. [49] Los primeros miembros basales del grupo, tradicionalmente llamados " pistoáuridos ", todavía eran en gran parte animales costeros. Sus cinturas escapulares permanecieron débiles, sus pelvis no podían soportar la potencia de una fuerte brazada de natación y sus aletas eran romas. Más tarde, un grupo de pistosaurios más avanzado se separó: los Plesiosauria. Estos tenían cinturas escapulares reforzadas, pelvis más planas y aletas más puntiagudas. Otras adaptaciones que les permitieron colonizar los mares abiertos incluyeron articulaciones rígidas en las extremidades; un aumento en el número de falanges de la mano y el pie; una conexión lateral más estrecha de la serie de falanges de los dedos de las manos y los pies, y una cola más corta. [50] [51]

Desde el Jurásico más temprano , la etapa Hettangiense , se conoce una rica radiación de plesiosaurios, lo que implica que el grupo ya debe haberse diversificado en el Triásico Tardío ; de esta diversificación, sin embargo, solo se han descubierto unas pocas formas (muy) basales, la más derivada Rhaeticosaurus . La evolución posterior de los plesiosaurios es muy controvertida. Los diversos análisis cladísticos no han dado como resultado un consenso sobre las relaciones entre los principales subgrupos de plesiosaurios. Tradicionalmente, los plesiosaurios se han dividido en Plesiosauroidea de cuello largo y Pliosauroidea de cuello corto . Sin embargo, la investigación moderna sugiere que algunos grupos generalmente de cuello largo podrían haber tenido miembros de cuello corto. Para evitar confusiones entre la filogenia , las relaciones evolutivas, y la morfología , la manera en que está construido el animal, las formas de cuello largo se denominan "plesiosauromorfas" y las de cuello corto "pliosauromorfas", sin que las especies "plesiosauromorfas" estén necesariamente más estrechamente relacionadas entre sí que con las formas "pliosauromorfas". [52]

El último ancestro común de los plesiosaurios fue probablemente una forma bastante pequeña de cuello corto. Durante el Jurásico temprano, el subgrupo con más especies fue el de los Rhomaleosauridae , una escisión posiblemente muy basal de especies que también eran de cuello corto. Los plesiosaurios en este período medían como máximo cinco metros (dieciséis pies) de largo. Para el Toarciense , hace unos 180 millones de años, otros grupos, entre ellos los Plesiosauridae , se volvieron más numerosos y algunas especies desarrollaron cuellos más largos, lo que resultó en longitudes corporales totales de hasta diez metros (33 pies). [53]
A mediados del Jurásico, aparecieron pliosauridos de gran tamaño , caracterizados por una cabeza grande y un cuello corto, como Liopleurodon y Simolestes . Estas formas tenían cráneos de hasta tres metros de largo y alcanzaban una longitud de hasta diecisiete metros y un peso de diez toneladas. Los pliosauridos tenían dientes grandes y cónicos y fueron los carnívoros marinos dominantes de su época. Durante la misma época, hace aproximadamente 160 millones de años, estaban presentes los Cryptoclididae , especies de menor tamaño, con un cuello largo y una cabeza pequeña. [54]
Los leptocléidos aparecieron durante el Cretácico Inferior . Eran formas bastante pequeñas que, a pesar de sus cuellos cortos, podrían haber estado más estrechamente relacionadas con los plesiosaurios que con los pliosaurios. Más tarde, en el Cretácico Inferior, aparecieron los elasmosáuridos ; estos estaban entre los plesiosaurios más largos, alcanzando hasta quince metros (cincuenta pies) de longitud debido a cuellos muy largos que contenían hasta 76 vértebras, más que cualquier otro vertebrado conocido. Los pliosaurios todavía estaban presentes, como lo demuestran los grandes depredadores, como Kronosaurus . [54]
A principios del Cretácico Superior , los ictiosaurios se extinguieron; tal vez un grupo de plesiosaurios evolucionó para llenar sus nichos: los policotílicos , que tenían cuellos cortos y cabezas peculiarmente alargadas con hocicos estrechos. Durante el Cretácico Superior, los elasmosáuridos todavía tenían muchas especies. [54]
Todos los plesiosaurios se extinguieron como resultado del evento KT al final del período Cretácico, hace aproximadamente 66 millones de años. [55]
En la filogenia moderna , los clados son grupos definidos que contienen todas las especies que pertenecen a una determinada rama del árbol evolutivo . Una forma de definir un clado es dejar que consista en el último ancestro común de dos de esas especies y todos sus descendientes. Un clado de este tipo se llama " clado de nodo ". En 2008, Patrick Druckenmiller y Anthony Russell definieron de esta manera a Plesiosauria como el grupo que consiste en el último ancestro común de Plesiosaurus dolichocheirus y Peloneustes philarchus y todos sus descendientes. [56] Plesiosaurus y Peloneustes representaban los subgrupos principales de Plesiosauroidea y Pliosauroidea y fueron elegidos por razones históricas; cualquier otra especie de estos grupos habría sido suficiente.
Otra forma de definir un clado es dejar que consista en todas las especies más estrechamente relacionadas con una especie determinada que uno en cualquier caso desea incluir en el clado que con otra especie que uno por el contrario desea excluir. Un clado de este tipo se llama un " clado de tallo ". Tal definición tiene la ventaja de que es más fácil incluir todas las especies con una cierta morfología . Plesiosauria fue definido en 2010 por Hillary Ketchum y Roger Benson como un taxón basado en tallos : "todos los taxones más estrechamente relacionados con Plesiosaurus dolichodeirus y Pliosaurus brachydeirus que con Augustasaurus hagdorni ". Ketchum y Benson (2010) también acuñaron un nuevo clado Neoplesiosauria, un taxón basado en nodos que fue definido por como " Plesiosauria dolichodeirus , Pliosaurus brachydeirus , su ancestro común más reciente y todos sus descendientes". [54] El clado Neoplesiosauria muy probablemente sea materialmente idéntico a Plesiosauria sensu Druckenmiller & Russell, por lo que designaría exactamente la misma especie, y el término pretendía reemplazar este concepto.
Benson et al. (2012) encontraron que los Pliosauroidea tradicionales eran parafiléticos en relación con los Plesiosauroidea. Se encontró que los Rhomaleosauridae estaban fuera de Neoplesiosauria, pero aún dentro de Plesiosauria. Se encontró que el pistosaurio del Carniense temprano Bobosaurus estaba un paso más avanzado que Augustasaurus en relación con Plesiosauria y, por lo tanto, representaba por definición el plesiosaurio más basal conocido. Este análisis se centró en los plesiosaurios basales y, por lo tanto, solo se incluyeron un pliosaurido derivado y un criptoclidio , mientras que los elasmosáuridos no se incluyeron en absoluto. Un análisis más detallado publicado tanto por Benson como por Druckenmiller en 2014 no pudo resolver las relaciones entre los linajes en la base de Plesiosauria. [57]

El siguiente cladograma sigue un análisis de Benson y Druckenmiller (2014). [57]


En general, los plesiosaurios variaban en longitud adulta de entre 1,5 metros (4,9 pies) a aproximadamente 15 metros (49 pies). El grupo contenía así algunos de los mayores depredadores marinos de ápice en el registro fósil , aproximadamente igualando en tamaño a los ictiosaurios , mosasáuridos , tiburones y ballenas dentadas más largos . Algunos restos de plesiosaurios, como un conjunto de mandíbulas inferiores altamente reconstruidas y fragmentarias de 2,875 metros (9,43 pies) de largo conservadas en el Museo de la Universidad de Oxford y atribuibles a Pliosaurus rossicus (anteriormente referido a Stretosaurus [58] y Liopleurodon ), indicaban una longitud de 17 metros (56 pies). Sin embargo, recientemente se argumentó que su tamaño no se puede determinar actualmente debido a que están mal reconstruidos y es más probable una longitud de 12,7 metros (42 pies) o menos. [59] Se estimó que MCZ 1285, un espécimen actualmente atribuible a Kronosaurus queenslandicus , del Cretácico Inferior de Australia , tenía una longitud de cráneo de 2,21–2,85 m (7,3–9,4 pies). [59] [60] Una serie de vértebras del cuello de la Formación Kimmeridge Clay indican un pliosaurio, probablemente Pliosaurus , que pudo haber medido hasta 14,4 metros (47 pies) de largo. [61]
El plesiosaurio típico tenía un cuerpo ancho y plano y una cola corta . Los plesiosaurios conservaron sus dos pares de extremidades ancestrales, que habían evolucionado hasta convertirse en grandes aletas . [62] Los plesiosaurios estaban relacionados con los anteriores Nothosauridae , [63] que tenían un cuerpo más parecido al de un cocodrilo. La disposición de las aletas es inusual para los animales acuáticos, ya que probablemente las cuatro extremidades se usaban para impulsar al animal a través del agua con movimientos hacia arriba y hacia abajo. La cola probablemente solo se usaba para ayudar en el control direccional. Esto contrasta con los ictiosaurios y los mosasaurios posteriores , en los que la cola proporcionaba la propulsión principal. [64]
Para impulsar las aletas, la cintura escapular y la pelvis se habían modificado en gran medida, desarrollándose en placas óseas anchas en la parte inferior del cuerpo, que servían como superficie de unión para grandes grupos musculares, capaces de tirar de las extremidades hacia abajo. En el hombro, el coracoides se había convertido en el elemento más grande que cubre la mayor parte del pecho. La escápula era mucho más pequeña, formando el borde frontal exterior del tronco. Hacia la mitad, continuaba en una clavícula y finalmente en un pequeño hueso interclavicular. Como en la mayoría de los tetrápodos , la articulación del hombro estaba formada por la escápula y el coracoides. En la pelvis, la placa ósea estaba formada por el isquion en la parte posterior y el hueso púbico más grande en frente de él. El íleon , que en los vertebrados terrestres soporta el peso de la extremidad trasera, se había convertido en un elemento pequeño en la parte posterior, ya no unido ni al hueso púbico ni al fémur. La articulación de la cadera estaba formada por el isquion y el hueso púbico. Las placas pectoral y pélvica estaban conectadas por un plastrón , una caja ósea formada por las costillas abdominales pareadas , cada una con una sección media y otra externa. Esta disposición inmovilizaba todo el tronco. [64]
Para convertirse en aletas, las extremidades habían cambiado considerablemente. Las extremidades eran muy grandes, cada una tan larga como el tronco. Las extremidades anteriores y posteriores se parecían mucho entre sí. El húmero en el brazo superior y el fémur en la pierna superior se habían convertido en grandes huesos planos, ensanchados en sus extremos exteriores. Las articulaciones del codo y de la rodilla ya no eran funcionales: el antebrazo y la pierna inferior no podían flexionarse en relación con los elementos de la extremidad superior, sino que formaban una continuación plana de ellos. Todos los huesos externos se habían convertido en elementos de soporte planos de las aletas, estrechamente conectados entre sí y apenas capaces de girar, flexionarse, extenderse o extenderse. Esto era cierto para el cúbito , el radio , los metacarpianos y los dedos, así como para la tibia , el peroné , los metatarsianos y los dedos de los pies. Además, para alargar las aletas, el número de falanges había aumentado, hasta dieciocho seguidas, un fenómeno llamado hiperfalangia . Las aletas no eran perfectamente planas, sino que tenían un perfil superior ligeramente curvado de forma convexa, como un perfil aerodinámico , para poder "volar" a través del agua. [64]

Aunque los plesiosaurios variaban poco en la estructura del tronco, y pueden ser llamados "conservadores" en este sentido, hubo grandes diferencias entre los subgrupos en lo que respecta a la forma del cuello y el cráneo. Los plesiosaurios pueden dividirse en dos tipos morfológicos principales que difieren en el tamaño de la cabeza y el cuello . Los "plesiosauromorfos", como Cryptoclididae , Elasmosauridae y Plesiosauridae , tenían cuellos largos y cabezas pequeñas. Los "pliosauromorfos", como Pliosauridae y Rhomaleosauridae , tenían cuellos más cortos con una cabeza grande y alargada. Las variaciones en la longitud del cuello no fueron causadas por un alargamiento de las vértebras cervicales individuales, sino por un aumento en su número. Elasmosaurus tiene setenta y dos vértebras cervicales; el récord conocido lo tiene el elasmosáurido Albertonectes , con setenta y seis cervicales. [65] La gran cantidad de articulaciones sugirió a los primeros investigadores que el cuello debe haber sido muy flexible; De hecho, se suponía que era posible una curvatura del cuello similar a la de un cisne (en islandés , los plesiosaurios incluso se llaman Svaneðlur , "lagartos cisne"). Sin embargo, la investigación moderna ha confirmado una conjetura anterior de Williston de que las largas espinas en forma de placa en la parte superior de las vértebras, el processus spinosi , limitaban fuertemente el movimiento vertical del cuello. Aunque la curvatura horizontal estaba menos restringida, en general, el cuello debe haber sido bastante rígido y ciertamente era incapaz de doblarse en espirales serpentinas. Esto es aún más cierto en el caso de los "pliosauromófos" de cuello corto, que tenían tan solo once vértebras cervicales. En las formas tempranas, las vértebras del cuello anficélicas o anfiplanas tenían costillas cervicales de dos cabezas; las formas posteriores tenían costillas de una sola cabeza. En el resto de la columna vertebral , el número de vértebras dorsales variaba entre aproximadamente diecinueve y treinta y dos; de las vértebras sacras, entre dos y seis, y de las vértebras de la cola, entre veintiuna y treinta y dos aproximadamente. Estas vértebras aún poseían los procesos originales heredados de los ancestros terrestres de los sauropterigios y no habían sido reducidas a simples discos parecidos a los de los peces, como sucedió con las vértebras de los ictiosaurios. Las vértebras de la cola poseían huesos en forma de chevron. Las vértebras dorsales de los plesiosaurios son fácilmente reconocibles por dos grandes forámenes subcentralia , aberturas vasculares pareadas en la parte inferior. [64]
El cráneo de los plesiosaurios mostraba la condición de " euryapsid ", careciendo de las fenestras temporales inferiores , las aberturas en los lados inferiores posteriores. Las fenestras temporales superiores formaban grandes aberturas a los lados del techo posterior del cráneo, la inserción de los músculos que cierran las mandíbulas inferiores. En general, los huesos parietales eran muy grandes, con una cresta en la línea media, mientras que los huesos escamosos típicamente formaban un arco, excluyendo los parietales del occipucio . Las cuencas oculares eran grandes, en general apuntando oblicuamente hacia arriba; los pliosauridos tenían ojos dirigidos más lateralmente. Los ojos estaban sostenidos por anillos esclerales , cuya forma muestra que eran relativamente planos, una adaptación al buceo. Las fosas nasales internas ubicadas anteriormente, las coanas , tienen surcos palatinos para canalizar el agua, cuyo flujo se mantendría por la presión hidrodinámica sobre las fosas nasales externas ubicadas posteriormente, frente a las cuencas oculares, durante la natación. Según una hipótesis, durante su paso por los conductos nasales, el agua habría sido 'olida' por los epitelios olfativos. [66] [67] Sin embargo, más atrás, hay un segundo par de aberturas en el paladar; una hipótesis posterior sostiene que estas son las coanas reales y el par delantero en realidad representaba glándulas salinas pareadas . [68] La distancia entre las cuencas de los ojos y las fosas nasales era tan limitada porque los huesos nasales estaban muy reducidos, incluso ausentes en muchas especies. Los premaxilares tocaban directamente los huesos frontales ; en los elasmosáuridos, incluso llegaban hasta los huesos parietales . A menudo, también faltaban los huesos lagrimales . [51]

La forma y el número de los dientes eran muy variables. Algunas formas tenían cientos de dientes con forma de aguja. La mayoría de las especies tenían dientes cónicos más grandes con una sección transversal redonda u ovalada. Dichos dientes eran de cuatro a seis en el premaxilar y de catorce a veinticinco en el maxilar ; el número en las mandíbulas inferiores era aproximadamente igual al del cráneo. Los dientes estaban colocados en alvéolos, tenían esmalte arrugado verticalmente y carena . En algunas especies, los dientes frontales eran notablemente más largos, para atrapar presas. [69]
Los restos de tejidos blandos de plesiosaurios son raros, pero a veces, especialmente en depósitos de esquisto , se han conservado parcialmente, por ejemplo, mostrando los contornos del cuerpo. Un descubrimiento temprano a este respecto fue el holotipo de Plesiosaurus conybeari (actualmente Attenborosaurus ). De tales hallazgos se sabe que la piel era lisa, sin escamas aparentes pero con pequeñas arrugas, que el borde posterior de las aletas se extendía considerablemente detrás de los huesos de las extremidades; [70] y que la cola tenía una aleta vertical, como informó Wilhelm Dames en su descripción de Plesiosaurus guilelmiimperatoris (actualmente Seeleyosaurus ). [71] La posibilidad de una aleta caudal ha sido confirmada por estudios recientes sobre la forma de la espina neural caudal de Pantosaurus , Cryptoclidus y Rhomaleosaurus zetlandicus . [72] [73] [74] Un estudio de 2020 afirma que la aleta caudal tenía una configuración horizontal. [75]


La probable fuente de alimento de los plesiosaurios variaba dependiendo de si pertenecían a las formas de cuello largo "plesiosauromorfas" o a las especies de cuello corto "pliosauromorfas".
Los cuellos extremadamente largos de los "plesiosauromorfos" han provocado especulaciones sobre su función desde el mismo momento en que se hizo evidente su constitución especial. Conybeare había ofrecido tres posibles explicaciones. El cuello podría haber servido para interceptar a los peces que se movían rápidamente en una persecución. Alternativamente, los plesiosaurios podrían haber descansado en el fondo del mar, mientras que la cabeza era enviada a buscar presas, lo que parecía confirmarse por el hecho de que los ojos estaban dirigidos relativamente hacia arriba. Finalmente, Conybeare sugirió la posibilidad de que los plesiosaurios nadaran en la superficie, dejando que sus cuellos se hundieran hacia abajo para buscar alimento en niveles inferiores. Todas estas interpretaciones suponían que el cuello era muy flexible. La idea moderna de que el cuello era, de hecho, bastante rígido, con un movimiento vertical limitado, ha hecho necesarias nuevas explicaciones. Una hipótesis es que la longitud del cuello permitió sorprender a los bancos de peces, llegando la cabeza antes de que la vista o la onda de presión de la trompa pudiera alertarlos. Los "plesiosauromorfos" cazaban visualmente, como lo demuestran sus grandes ojos, y tal vez empleaban un sentido direccional del olfato. Los cefalópodos de cuerpo duro y blando probablemente formaban parte de su dieta. Sus mandíbulas probablemente eran lo suficientemente fuertes como para morder a través de los caparazones duros de este tipo de presa. Se han encontrado especímenes fósiles con caparazones de cefalópodos todavía en su estómago. [76] Los peces óseos ( Osteichthyes ), que se diversificaron aún más durante el Jurásico, probablemente también fueron presas. Una hipótesis muy diferente afirma que los "plesiosauromorfos" se alimentaban en el fondo. Los cuellos rígidos se habrían utilizado para arar el fondo del mar, comiendo el bentos . Esto se habría demostrado por los largos surcos presentes en los antiguos fondos marinos. [77] [78] En 2017 se sugirió un estilo de vida de este tipo para Morturneria . [79] Los "plesiosauromorfos" no estaban bien adaptados para atrapar presas grandes y de rápido movimiento, ya que sus largos cuellos, aunque aparentemente aerodinámicos, causaban una enorme fricción en la piel . Sankar Chatterjee sugirió en 1989 que algunos Cryptocleididae se alimentaban por suspensión y filtraban el plancton . Aristonectes , por ejemplo, tenía cientos de dientes, lo que le permitía filtrar pequeños crustáceos del agua. [80]
Los "pliosauromorfos" de cuello corto eran carnívoros superiores, o depredadores de ápice , en sus respectivas redes alimentarias . [81] Eran depredadores de persecución [82] o de emboscada de presas de varios tamaños y alimentadores oportunistas; sus dientes podían usarse para perforar presas de cuerpo blando, especialmente peces. [83] Sus cabezas y dientes eran muy grandes, adecuados para agarrar y destrozar animales grandes. Su morfología les permitía nadar a gran velocidad. También cazaban visualmente.
Los plesiosaurios eran presas de otros carnívoros, como lo demuestran las marcas de mordedura dejadas por un tiburón que se han descubierto en una aleta fosilizada de plesiosaurio [84] y los restos fosilizados del contenido del estómago de un mosasaurio que se cree que son los restos de un plesiosaurio. [85]
También se han descubierto esqueletos con gastrolitos , piedras, en sus estómagos, aunque no se ha establecido si para ayudar a descomponer los alimentos, especialmente los cefalópodos, en una molleja muscular, o para variar la flotabilidad , o ambos. [86] [87] Sin embargo, el peso total de los gastrolitos encontrados en varios especímenes parece ser insuficiente para modificar la flotabilidad de estos grandes reptiles. [88] Los primeros gastrolitos de plesiosaurio, encontrados con Mauisaurus gardneri (un nomen nudum [89] ), fueron reportados por Harry Govier Seeley en 1877. [90] El número de estas piedras por individuo es a menudo muy grande. En 1949, un fósil de Alzadasaurus (espécimen SDSM 451, posteriormente renombrado a Styxosaurus ) mostró 253 de ellos. [91] El tamaño de las piedras individuales es a menudo considerable. En 1991 se investigó un espécimen de elasmosáurido, KUVP 129744, que contenía un gastrolito con un diámetro de diecisiete centímetros y un peso de 1300 gramos; y una piedra algo más corta de 1490 gramos. En total, estaban presentes cuarenta y siete gastrolitos, con un peso combinado de 13 kilogramos. El tamaño de las piedras se ha visto como una indicación de que no fueron tragadas por accidente, sino deliberadamente, el animal quizás cubriendo grandes distancias en busca de un tipo de roca adecuado. [92] El espécimen tipo de Scalamagnus (MNA V10046) está asociado con 289 gastrolitos , lo cual es inusual en comparación con la mayoría de los esqueletos de policotílidos que generalmente carecen de gastrolitos. Con un rango de menos de 0,1 gramos a 18,5 gramos, la masa total de los gastrolitos fue de aproximadamente 518 gramos. Aproximadamente tres cuartas partes de las piedras pesaban menos de 2 gramos, y la masa media y la masa mediana de las piedras se estimaron en 1,9 gramos y 0,8 gramos respectivamente. Los gastrolitos tenían un valor medio alto y variabilidad en la esfericidad, lo que sugiere que este individuo obtenía sus piedras de ríos ubicados a lo largo del lado occidental de la vía marítima interior occidental. [93]
La característica forma de cuatro aletas del cuerpo ha provocado una considerable especulación sobre el tipo de movimiento que utilizaban los plesiosaurios. El único grupo moderno con cuatro aletas son las tortugas marinas, que solo utilizan el par delantero para propulsarse. Conybeare y Buckland ya habían comparado las aletas con las alas de las aves. Sin embargo, tal comparación no fue muy informativa, ya que la mecánica del vuelo de las aves en este período era poco conocida. A mediados del siglo XIX, se suponía generalmente que los plesiosaurios empleaban un movimiento de remo. Las aletas habrían sido movidas hacia adelante en una posición horizontal, para minimizar la fricción, y luego rotadas axialmente a una posición vertical para ser tiradas hacia atrás, causando la mayor fuerza reactiva posible . De hecho, tal método sería muy ineficiente: el movimiento de recuperación en este caso no genera empuje y el movimiento hacia atrás genera una enorme turbulencia. A principios del siglo XX, los principios recién descubiertos del vuelo de las aves sugirieron a varios investigadores que los plesiosaurios, como las tortugas y los pingüinos, realizaban un movimiento de vuelo mientras nadaban. Esto fue propuesto por ejemplo por Eberhard Fraas en 1905, [94] y en 1908 por Othenio Abel . [95] Al volar, el movimiento de la aleta es más vertical, su punta describe un óvalo u "8". Idealmente, la aleta se mueve primero oblicuamente hacia adelante y hacia abajo y luego, después de una ligera retracción y rotación, cruza este camino desde abajo para ser tirada hacia adelante y hacia arriba. Durante ambas brazadas, hacia abajo y hacia arriba, según el principio de Bernoulli , el empuje hacia adelante y hacia arriba es generado por el perfil superior convexo y curvado de la aleta, el borde delantero ligeramente inclinado con respecto al flujo de agua, mientras que la turbulencia es mínima. Sin embargo, a pesar de las evidentes ventajas de tal método de natación, en 1924 el primer estudio sistemático sobre la musculatura de los plesiosaurios por David Meredith Seares Watson concluyó que, no obstante, realizaban un movimiento de remo. [96]
A mediados del siglo XX, el "modelo de remo" de Watson siguió siendo la hipótesis dominante en relación con la brazada de natación del plesiosaurio. En 1957, Lambert Beverly Halstead , que en ese momento utilizaba el apellido Tarlo, propuso una variante: las extremidades traseras habrían remado en el plano horizontal, pero las extremidades delanteras habrían remado, moviéndose hacia abajo y hacia atrás. [97] [98] En 1975, el modelo tradicional fue desafiado por Jane Ann Robinson , quien revivió la hipótesis del "vuelo". Argumentó que los principales grupos musculares estaban ubicados de manera óptima para un movimiento de aleta vertical, no para tirar de las extremidades horizontalmente, y que la forma de las articulaciones de los hombros y la cadera habría impedido la rotación vertical necesaria para remar. [99] En un artículo posterior, Robinson propuso que la energía cinética generada por las fuerzas ejercidas sobre el tronco por las brazadas, se habría almacenado y liberado como energía elástica en la caja torácica, lo que permitiría un sistema de propulsión especialmente eficiente y dinámico. [100]
En el modelo de Robinson, tanto la palada descendente como la ascendente habrían sido potentes. En 1982, fue criticada por Samuel Tarsitano, Eberhard Frey y Jürgen Riess, quienes afirmaron que, si bien los músculos de la parte inferior de las placas del hombro y la pelvis eran claramente lo suficientemente potentes como para tirar de las extremidades hacia abajo, simplemente faltaban grupos musculares comparables en la parte superior de estas placas para elevar las extremidades y, de haber estado presentes, no se podrían haber empleado con fuerza, ya que su abultamiento conlleva el peligro de dañar los órganos internos. Propusieron un modelo de vuelo más limitado en el que se combinaba una potente palada descendente con una recuperación en gran medida sin potencia, en la que la aleta volvía a su posición original por el impulso del cuerpo que se movía hacia adelante y se hundía temporalmente. [101] [102] Este modelo de vuelo modificado se convirtió en una interpretación popular. En 1984, Stephen Godfrey prestó menos atención a una hipótesis alternativa que proponía que tanto las extremidades anteriores como las posteriores realizaban un movimiento de remo profundo hacia atrás combinado con una brazada de recuperación potente hacia adelante, similar al movimiento realizado por las extremidades anteriores de los leones marinos. [103]
En 2010, Frank Sanders y Kenneth Carpenter publicaron un estudio en el que concluían que el modelo de Robinson había sido correcto. Frey y Riess se habrían equivocado al afirmar que las placas del hombro y la pelvis no tenían músculos adheridos a sus lados superiores. Si bien estos grupos musculares probablemente no eran muy potentes, esto podría haberse compensado fácilmente con los grandes músculos de la espalda, especialmente el dorsal ancho , que habría estado bien desarrollado en vista de las altas espinas de la columna vertebral. Además, la estructura plana de las articulaciones del hombro y la cadera indicaba claramente que el movimiento principal era vertical, no horizontal. [104]

Como todos los tetrápodos con extremidades, los plesiosaurios deben haber tenido un cierto modo de andar , un patrón de movimiento coordinado de las, en este caso, aletas. De todas las posibilidades, en la práctica la atención se ha dirigido en gran medida a la cuestión de si el par delantero y el par trasero se movían simultáneamente, de modo que las cuatro aletas se activaban al mismo tiempo, o en un patrón alterno, empleándose cada par por turno. Frey y Riess en 1991 propusieron un modelo alternativo, que habría tenido la ventaja de una propulsión más continua. [105] En 2000, Theagarten Lingham-Soliar evadió la cuestión al concluir que, al igual que las tortugas marinas, los plesiosaurios solo usaban el par delantero para una brazada de potencia. El par trasero se habría utilizado simplemente para dirigir. Lingham-Soliar dedujo esto de la forma de la articulación de la cadera, que habría permitido solo un movimiento vertical limitado. Además, una separación de la función de propulsión y de dirección habría facilitado la coordinación general del cuerpo y evitado un cabeceo demasiado extremo . Rechazó la hipótesis de Robinson de que la energía elástica se almacenaba en la caja torácica, considerando que las costillas eran demasiado rígidas para ello. [106]
La interpretación de Frey y Riess se convirtió en la dominante, pero fue cuestionada en 2004 por Sanders, quien demostró experimentalmente que, mientras que un movimiento alterno podría haber causado un cabeceo excesivo, un movimiento simultáneo habría causado solo un ligero cabeceo, que podría haber sido fácilmente controlado por las aletas traseras. De los otros movimientos axiales, el balanceo podría haberse controlado activando alternativamente las aletas del lado derecho o izquierdo, y la guiñada por el cuello largo o una aleta caudal vertical. Sanders no creía que el par trasero no se usara para la propulsión, concluyendo que las limitaciones impuestas por la articulación de la cadera eran muy relativas. [107] En 2010, Sanders y Carpenter concluyeron que, con una marcha alterna, la turbulencia causada por el par delantero habría obstaculizado una acción efectiva del par trasero. Además, una larga fase de planeo después de un enganche simultáneo habría sido muy eficiente energéticamente. [104] También es posible que la marcha fuera opcional y se adaptara a las circunstancias. Durante una persecución rápida y constante, un movimiento alternativo habría sido útil; en una emboscada, un golpe simultáneo habría permitido alcanzar una velocidad máxima. Al buscar presas a una distancia mayor, una combinación de un movimiento simultáneo con planeo habría consumido menos energía. [108] En 2017, un estudio de Luke Muscutt, utilizando un modelo de robot, concluyó que las aletas traseras se empleaban activamente, lo que permitía un aumento del 60% de la fuerza de propulsión y un aumento del 40% de la eficiencia. No habría habido una única fase óptima para todas las condiciones, ya que es probable que la marcha se hubiera modificado según lo exigiera la situación. [109]

En general, es difícil determinar la velocidad máxima de las criaturas marinas extintas. En el caso de los plesiosaurios, esto se hace más difícil por la falta de consenso sobre el movimiento de sus aletas y su forma de andar. No existen cálculos exactos de su número de Reynolds . Las impresiones fósiles muestran que la piel era relativamente lisa, sin escamas, y esto puede haber reducido la resistencia al avance de la forma . [104] Hay pequeñas arrugas presentes en la piel que pueden haber impedido la separación del flujo laminar en la capa límite y, por lo tanto, reducido la fricción de la piel .
La velocidad sostenida puede estimarse calculando la resistencia de un modelo simplificado del cuerpo, que puede aproximarse mediante un esferoide alargado , y el nivel sostenible de producción de energía por los músculos . Un primer estudio de este problema fue publicado por Judy Massare en 1988. [110] Incluso al asumir una baja eficiencia hidrodinámica de 0,65, el modelo de Massare parecía indicar que los plesiosaurios, si fueran de sangre caliente, habrían navegado a una velocidad de cuatro metros por segundo, o aproximadamente catorce kilómetros por hora, excediendo considerablemente las velocidades conocidas de los delfines y ballenas actuales. [111] Sin embargo, en 2002 Ryosuke Motani demostró que las fórmulas que Massare había utilizado eran defectuosas. Un nuevo cálculo, utilizando fórmulas corregidas, dio como resultado una velocidad de medio metro por segundo (1,8 km/h) para un plesiosaurio de sangre fría y un metro y medio por segundo (5,4 km/h) para un plesiosaurio endotérmico . Incluso la estimación más alta es aproximadamente un tercio menor que la velocidad de los cetáceos actuales . [112]
Massare también intentó comparar las velocidades de los plesiosaurios con las de los otros dos grupos principales de reptiles marinos, los Ichthyosauria y los Mosasauridae . Concluyó que los plesiosaurios eran aproximadamente un veinte por ciento más lentos que los ictiosaurios avanzados, que empleaban un movimiento tuniforme muy efectivo, oscilando solo la cola, pero un cinco por ciento más rápido que los mosasáuridos, que se suponía que nadaban con un movimiento anguiliforme ineficiente, similar al de la anguila, del cuerpo. [111]
Las numerosas especies de plesiosaurios pueden haber diferido considerablemente en sus velocidades de natación, lo que refleja las diversas formas corporales presentes en el grupo. Mientras que los "pliosauromorfos" de cuello corto (por ejemplo, Liopleurodon ) pueden haber sido nadadores rápidos, los "plesiosauromorfos" de cuello largo estaban construidos más para la maniobrabilidad que para la velocidad, ralentizados por una fuerte fricción de la piel, pero capaces de un rápido movimiento de balanceo. Algunas formas de cuello largo, como los Elasmosauridae , también tienen aletas rechonchas relativamente cortas con una relación de aspecto baja , lo que reduce aún más la velocidad pero mejora el balanceo. [113]
Hay pocos datos disponibles que muestren exactamente a qué profundidad se sumergían los plesiosaurios. Que buceaban a una profundidad considerable está probado por los rastros de enfermedad por descompresión . Las cabezas de los húmeros y los fémures con muchos fósiles muestran necrosis del tejido óseo, causada por un ascenso demasiado rápido después de una inmersión profunda. Sin embargo, esto no permite deducir una profundidad exacta ya que el daño podría haber sido causado por unas pocas inmersiones muy profundas, o alternativamente por un gran número de descensos relativamente superficiales. Las vértebras no muestran tal daño: probablemente estaban protegidas por un suministro de sangre superior, hecho posible por las arterias que ingresaban al hueso a través de los dos agujeros subcentrales , grandes aberturas en sus caras inferiores. [114]
El descenso se habría visto facilitado por una fuerza de Arquímedes negativa , es decir, por ser más denso que el agua. Por supuesto, esto habría tenido la desventaja de dificultar la subida de nuevo. Los plesiosaurios jóvenes muestran paquiosis , una densidad extrema del tejido óseo, que podría haber aumentado el peso relativo. Los individuos adultos tienen más hueso esponjoso. Se han sugerido los gastrolitos como un método para aumentar el peso [115] o incluso como un medio para alcanzar la flotabilidad neutra , tragándolos o escupiéndolos de nuevo según fuera necesario. [116] También podrían haber sido utilizados para aumentar la estabilidad. [117]
Los ojos relativamente grandes de Cryptocleididae se han considerado como una adaptación al buceo profundo. [118]
Un estudio de 2020 ha postulado que los sauropterigios dependían de movimientos de cola verticales de forma muy similar a los cetáceos . En los plesiosaurios, la trompa era rígida, por lo que esta acción era más limitada y se realizaba en conjunción con las aletas. [75]
Tradicionalmente, se suponía que los grupos de reptiles extintos eran de sangre fría como los reptiles modernos. Nuevas investigaciones durante las últimas décadas han llevado a la conclusión de que algunos grupos, como los dinosaurios terópodos y los pterosaurios , muy probablemente eran de sangre caliente. Si quizás los plesiosaurios también eran de sangre caliente es difícil de determinar. Una de las indicaciones de un metabolismo alto es la presencia de hueso fibrolamelar de rápido crecimiento. Sin embargo, la paquiosis con individuos juveniles hace que sea difícil establecer si los plesiosaurios poseían dicho hueso. Sin embargo, ha sido posible verificar su ocurrencia con miembros más basales del grupo más inclusivo al que pertenecían los plesiosaurios, los Sauropterigia . Un estudio en 2010 concluyó que el hueso fibrolamelar estaba originalmente presente en los sauropterigios. [119] Una publicación posterior en 2013 encontró que los Nothosauridae carecían de este tipo de matriz ósea pero que los Pistosauria basales lo poseían, un signo de un metabolismo más elevado. [120] Por lo tanto, es más parsimonioso suponer que los pistosaurios más derivados, los plesiosaurios, también tenían un metabolismo más rápido. Un artículo publicado en 2018 afirmó que los plesiosaurios tenían tasas metabólicas en reposo (RMR) en el rango de las aves según el modelo osteohistológico cuantitativo. [121] Sin embargo, estos resultados son problemáticos en vista de los principios generales de la fisiología de los vertebrados (ver la ley de Kleiber ); la evidencia de los estudios isotópicos del esmalte dental de los plesiosaurios de hecho sugiere endotermia a RMR más bajas, con temperaturas corporales inferidas de ca. 26 °C (79 °F). [122]

Como los reptiles en general son ovíparos , hasta finales del siglo XX se había visto como posible que plesiosaurios más pequeños pudieran haber subido a una playa para poner huevos, como las tortugas modernas . Sus fuertes extremidades y una parte inferior plana parecían haber hecho esto factible. Este método fue defendido, por ejemplo, por Halstead. Sin embargo, como esas extremidades ya no tenían articulaciones funcionales en los codos o las rodillas y la parte inferior por su propia planitud habría generado mucha fricción, ya en el siglo XIX se planteó la hipótesis de que los plesiosaurios habían sido vivíparos . Además, era difícil concebir cómo las especies más grandes, tan grandes como las ballenas, podrían haber sobrevivido a un varamiento. Los hallazgos fósiles de embriones de ictiosaurios mostraron que al menos un grupo de reptiles marinos había parido crías vivas. El primero en afirmar que se habían encontrado embriones similares en plesiosaurios fue Harry Govier Seeley , quien informó en 1887 de haber adquirido un nódulo con cuatro a ocho esqueletos diminutos. [123] En 1896, describió este descubrimiento con más detalle. [124] De ser auténticos, los embriones de plesiosaurios habrían sido muy pequeños, como los de los ictiosaurios. Sin embargo, en 1982 Richard Anthony Thulborn demostró que Seeley había sido engañado por un fósil "manipulado" de un nido de cangrejos de río. [125]
Un espécimen real de plesiosaurio encontrado en 1987 finalmente demostró que los plesiosaurios daban a luz a crías vivas: [126] Este fósil de una Polycotylus latippinus embarazada muestra que estos animales dieron a luz a un solo juvenil grande y probablemente invirtieron cuidado parental en su descendencia, similar a las ballenas modernas. La cría medía 1,5 metros (cinco pies) de largo y, por lo tanto, era grande en comparación con su madre de cinco metros (dieciséis pies) de longitud, lo que indica una estrategia K en la reproducción. [127] Poco se sabe sobre las tasas de crecimiento o un posible dimorfismo sexual .
Del cuidado parental indicado por el gran tamaño de las crías, se puede deducir que el comportamiento social en general era relativamente complejo. [126] No se sabe si los plesiosaurios cazaban en manadas. Su tamaño cerebral relativo parece ser típico de los reptiles. De los sentidos, la vista y el olfato eran importantes, el oído menos; los elasmosáuridos han perdido completamente el estribo . Se ha sugerido que en algunos grupos el cráneo albergaba órganos electrosensibles. [128] [129]
Algunos fósiles de plesiosaurio presentan patologías , fruto de enfermedades o de la vejez. En 2012 se describió una mandíbula de Pliosaurus con una articulación mandibular claramente afectada por artritis , un signo típico de senescencia . [130]
Se han encontrado fósiles de plesiosaurio en todos los continentes, incluida la Antártida . [131]
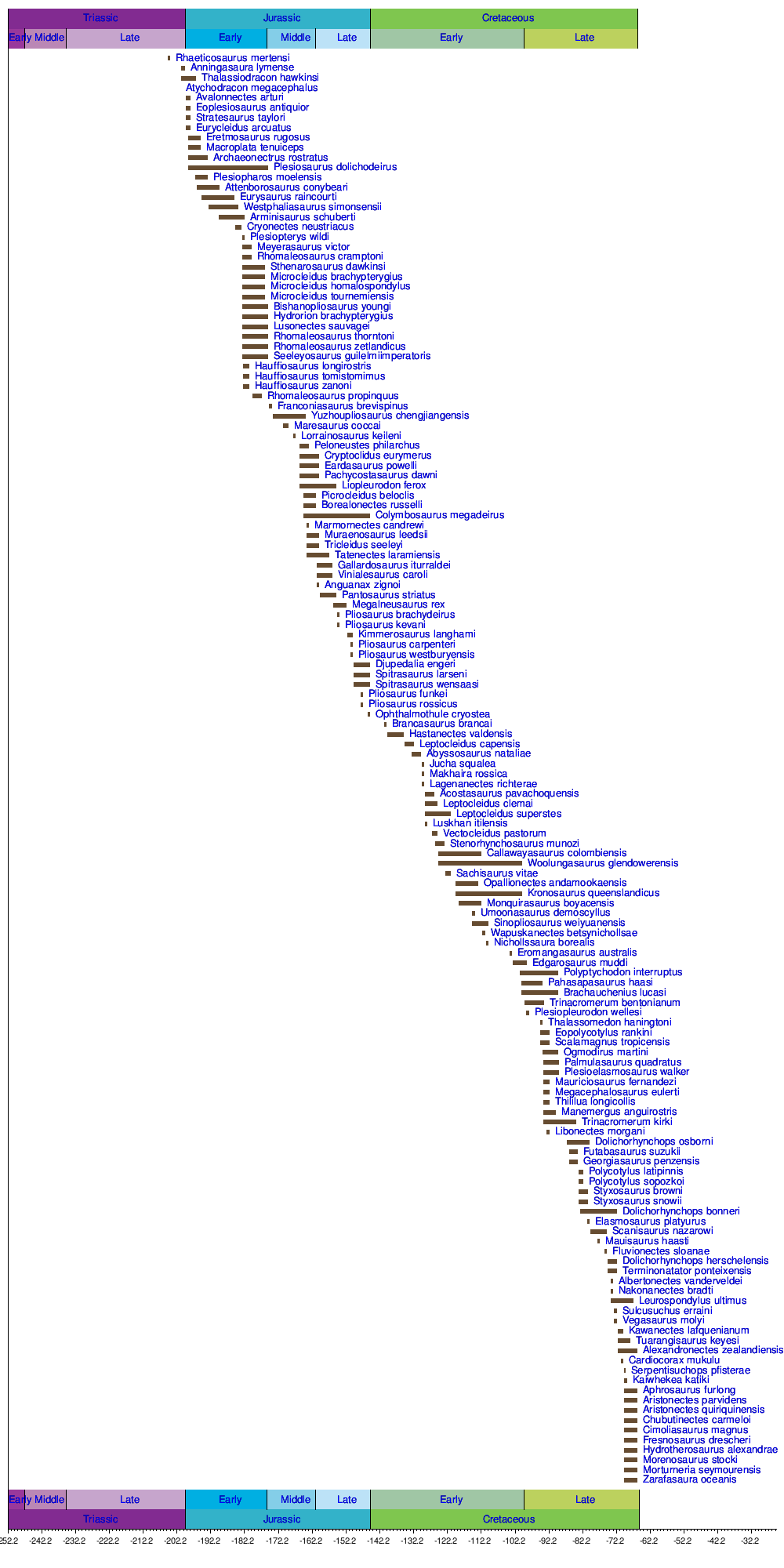
La siguiente es una lista de formaciones geológicas que han producido fósiles de plesiosaurio.
_trailer_-_Plesiosaurus_1.png/1280px-When_Dinosaurs_Ruled_the_Earth_(1970)_trailer_-_Plesiosaurus_1.png)
La creencia de que los plesiosaurios son dinosaurios es un error común , y los plesiosaurios a menudo son representados erróneamente como dinosaurios en la cultura popular. [138] [139]
Se ha sugerido que las leyendas de serpientes marinas y los avistamientos modernos de supuestos monstruos en lagos o en el mar podrían explicarse por la supervivencia de los plesiosaurios hasta los tiempos modernos. Esta propuesta criptozoológica ha sido rechazada por la comunidad científica en general, que la considera basada en la fantasía y la pseudociencia . Se ha demostrado que los supuestos cadáveres de plesiosaurios son en realidad cadáveres parcialmente descompuestos de tiburones peregrinos . [140] [141] [142]
Aunque a menudo se dice que el monstruo del Lago Ness se parece a un plesiosaurio, también se suele decir que tiene un aspecto completamente diferente. Se han presentado varias razones para considerar improbable que sea un plesiosaurio. Entre ellas se incluyen la suposición de que el agua del lago es demasiado fría para que un supuesto reptil de sangre fría pueda sobrevivir fácilmente, el hecho de que la osteología del cuello del plesiosaurio hace que sea absolutamente seguro decir que el plesiosaurio no podría levantar la cabeza como un cisne fuera del agua como lo hace el monstruo del Lago Ness, la suposición de que los animales que respiran aire serían fáciles de ver siempre que aparecieran en la superficie para respirar, [143] el hecho de que el lago es demasiado pequeño y no contiene suficiente comida para poder sustentar una colonia reproductora de animales grandes y, por último, el hecho de que el lago se formó hace solo 10.000 años al final de la última edad de hielo , y la última aparición fósil de plesiosaurios data de hace más de 66 millones de años. [144] Las explicaciones frecuentes para los avistamientos incluyen olas , objetos inanimados flotantes, trucos de luz, animales conocidos que nadan y bromas pesadas. [145] Sin embargo, en la imaginación popular, los plesiosaurios han llegado a ser identificados con el Monstruo del Lago Ness. Esto ha hecho que los plesiosaurios sean más conocidos por el público en general. [146]