
El núcleo celular (del latín núcleo o nuculeus 'grano, semilla'; pl.: núcleos ) es un orgánulo rodeado de membrana que se encuentra en las células eucariotas . Las células eucariotas suelen tener un solo núcleo, pero unos pocos tipos de células, como los glóbulos rojos de los mamíferos , no tienen núcleo, y unos pocos otros, incluidos los osteoclastos , tienen muchos . Las principales estructuras que componen el núcleo son la envoltura nuclear , una membrana doble que encierra todo el orgánulo y aísla su contenido del citoplasma celular ; y la matriz nuclear , una red dentro del núcleo que agrega soporte mecánico.
El núcleo celular contiene casi todo el genoma de la célula . El ADN nuclear suele estar organizado en múltiples cromosomas : largas cadenas de ADN salpicadas de diversas proteínas , como las histonas , que protegen y organizan el ADN. Los genes dentro de estos cromosomas están estructurados de tal manera que promueven la función celular. El núcleo mantiene la integridad de los genes y controla las actividades de la célula regulando la expresión genética .
Debido a que la envoltura nuclear es impermeable a las moléculas grandes , se requieren poros nucleares para regular el transporte nuclear de moléculas a través de la envoltura. Los poros cruzan ambas membranas nucleares, proporcionando un canal a través del cual las moléculas más grandes deben ser transportadas activamente por proteínas transportadoras mientras se permite el libre movimiento de moléculas pequeñas e iones . El movimiento de moléculas grandes como proteínas y ARN a través de los poros es necesario tanto para la expresión genética como para el mantenimiento de los cromosomas. Aunque el interior del núcleo no contiene ningún subcompartimento limitado por la membrana, existen varios cuerpos nucleares , compuestos de proteínas únicas, moléculas de ARN y partes particulares de los cromosomas. El más conocido de ellos es el nucléolo , involucrado en el ensamblaje de los ribosomas .

El núcleo celular contiene la mayor parte del material genético de la célula en forma de múltiples moléculas lineales de ADN organizadas en estructuras llamadas cromosomas . Cada célula humana contiene aproximadamente dos metros de ADN. [1] : 405 Durante la mayor parte del ciclo celular, estos están organizados en un complejo de ADN-proteína conocido como cromatina , y durante la división celular se puede ver que la cromatina forma los cromosomas bien definidos que se conocen a partir de un cariotipo . Una pequeña fracción de los genes de la célula se encuentran en cambio en las mitocondrias . [1] : 438
Existen dos tipos de cromatina. La eucromatina es la forma menos compacta del ADN y contiene genes que la célula expresa con frecuencia. [2] El otro tipo, la heterocromatina , es la forma más compacta y contiene ADN que se transcribe con poca frecuencia. Esta estructura se clasifica además en heterocromatina facultativa , que consiste en genes que se organizan como heterocromatina solo en ciertos tipos de células o en ciertas etapas del desarrollo, y heterocromatina constitutiva que consiste en componentes estructurales de los cromosomas, como los telómeros y los centrómeros . [3] Durante la interfase, la cromatina se organiza en parches individuales discretos, [4] llamados territorios cromosómicos . [5] Los genes activos, que generalmente se encuentran en la región eucromática del cromosoma, tienden a ubicarse hacia el límite del territorio del cromosoma. [6]
Los anticuerpos contra ciertos tipos de organización de la cromatina, en particular los nucleosomas , se han asociado con una serie de enfermedades autoinmunes , como el lupus eritematoso sistémico . [7] Estos se conocen como anticuerpos antinucleares (ANA) y también se han observado en conjunto con la esclerosis múltiple como parte de una disfunción general del sistema inmunológico. [8]

El núcleo contiene casi todo el ADN de la célula , rodeado por una red de filamentos intermedios fibrosos llamada matriz nuclear , y está envuelto en una membrana doble llamada envoltura nuclear . La envoltura nuclear separa el líquido dentro del núcleo, llamado nucleoplasma , del resto de la célula. El tamaño del núcleo está correlacionado con el tamaño de la célula, y esta relación se informa en una variedad de tipos y especies de células. [9] En eucariotas, el núcleo en muchas células ocupa típicamente el 10% del volumen celular. [10] : 178 El núcleo es el orgánulo más grande en las células animales. [11] : 12 En las células humanas, el diámetro del núcleo es de aproximadamente seis micrómetros (μm). [10] : 179
La envoltura nuclear consta de dos membranas , una membrana nuclear interna y otra externa , perforadas por poros nucleares . [10] : 649 Juntas, estas membranas sirven para separar el material genético de la célula del resto del contenido celular y permiten que el núcleo mantenga un entorno distinto del resto de la célula. A pesar de su estrecha aposición alrededor de gran parte del núcleo, las dos membranas difieren sustancialmente en forma y contenido. La membrana interna rodea el contenido nuclear, proporcionando su borde definitorio. [11] : 14 Incrustadas dentro de la membrana interna, varias proteínas se unen a los filamentos intermedios que dan al núcleo su estructura. [10] : 649 La membrana externa encierra la membrana interna y es continua con la membrana del retículo endoplasmático adyacente . [10] : 649 Como parte de la membrana del retículo endoplasmático, la membrana nuclear externa está tachonada de ribosomas que traducen activamente proteínas a través de la membrana. [10] : 649 El espacio entre las dos membranas se llama espacio perinuclear y es continuo con el lumen del retículo endoplásmico . [10] : 649
En una envoltura nuclear de mamífero hay entre 3000 y 4000 complejos de poros nucleares (NPC) que perforan la envoltura. [10] : 650 Cada NPC contiene una estructura en forma de anillo óctuplemente simétrica en una posición donde se fusionan las membranas interna y externa. [12] El número de NPC puede variar considerablemente entre tipos de células; las células gliales pequeñas solo tienen unos pocos cientos, y las células de Purkinje grandes tienen alrededor de 20.000. [10] : 650 El NPC proporciona transporte selectivo de moléculas entre el nucleoplasma y el citosol . [13] El complejo de poros nucleares está compuesto de aproximadamente treinta proteínas diferentes conocidas como nucleoporinas . [10] : 649 Los poros tienen un peso molecular de alrededor de 60 a 80 millones de daltons y constan de alrededor de 50 (en levaduras ) a varios cientos de proteínas (en vertebrados ). [11] : 622–4 Los poros tienen un diámetro total de 100 nm; sin embargo, el espacio a través del cual las moléculas se difunden libremente tiene solo unos 9 nm de ancho, debido a la presencia de sistemas reguladores dentro del centro del poro. Este tamaño permite selectivamente el paso de pequeñas moléculas solubles en agua mientras evita que moléculas más grandes, como ácidos nucleicos y proteínas más grandes, entren o salgan del núcleo de manera inapropiada. Estas moléculas grandes deben transportarse activamente al núcleo. Adherida al anillo hay una estructura llamada canasta nuclear que se extiende hacia el nucleoplasma y una serie de extensiones filamentosas que llegan hasta el citoplasma. Ambas estructuras sirven para mediar la unión a las proteínas de transporte nuclear. [1] : 509–10
La mayoría de las proteínas, subunidades ribosomales y algunos ARN se transportan a través de los complejos de poros en un proceso mediado por una familia de factores de transporte conocidos como carioferinas . Las carioferinas que median el movimiento hacia el núcleo también se denominan importinas, mientras que las que median el movimiento fuera del núcleo se denominan exportinas. La mayoría de las carioferinas interactúan directamente con su carga, aunque algunas utilizan proteínas adaptadoras . [14] Las hormonas esteroides como el cortisol y la aldosterona , así como otras pequeñas moléculas liposolubles implicadas en la señalización intercelular , pueden difundirse a través de la membrana celular y hacia el citoplasma, donde se unen a las proteínas receptoras nucleares que se trafican hacia el núcleo. Allí sirven como factores de transcripción cuando se unen a su ligando ; en ausencia de un ligando, muchos de estos receptores funcionan como histonas desacetilasas que reprimen la expresión génica. [1] : 488
En las células animales, dos redes de filamentos intermedios proporcionan soporte mecánico al núcleo: la lámina nuclear forma una malla organizada en la cara interna de la envoltura, mientras que en la cara citosólica de la envoltura se proporciona un soporte menos organizado. Ambos sistemas proporcionan soporte estructural a la envoltura nuclear y sitios de anclaje para los cromosomas y los poros nucleares. [15]
La lámina nuclear está compuesta principalmente por proteínas laminosas . Como todas las proteínas, las laminas se sintetizan en el citoplasma y luego se transportan al interior del núcleo, donde se ensamblan antes de incorporarse a la red existente de láminas nucleares. [16] [17] Las laminas que se encuentran en la cara citosólica de la membrana, como la emerina y la nesprina , se unen al citoesqueleto para proporcionar soporte estructural. Las laminas también se encuentran dentro del nucleoplasma donde forman otra estructura regular, conocida como velo nucleoplásmico , [18] [19] que es visible mediante microscopía de fluorescencia . La función real del velo no está clara, aunque está excluido del nucléolo y está presente durante la interfase . [20] Las estructuras laminosas que componen el velo, como LEM3 , se unen a la cromatina y la alteración de su estructura inhibe la transcripción de genes codificadores de proteínas. [21]
Al igual que los componentes de otros filamentos intermedios, el monómero de lámina contiene un dominio alfa-helicoidal utilizado por dos monómeros para enrollarse uno alrededor del otro, formando una estructura de dímero llamada bobina enrollada . Luego, dos de estas estructuras de dímero se unen una al lado de la otra, en una disposición antiparalela , para formar un tetrámero llamado protofilamento . Ocho de estos protofilamentos forman una disposición lateral que se retuerce para formar un filamento similar a una cuerda . Estos filamentos se pueden ensamblar o desensamblar de manera dinámica, lo que significa que los cambios en la longitud del filamento dependen de las velocidades competitivas de adición y eliminación de filamentos. [15]
Las mutaciones en los genes de las láminas que provocan defectos en el ensamblaje de los filamentos causan un grupo de trastornos genéticos raros conocidos como laminopatías . La laminopatía más notable es la familia de enfermedades conocida como progeria , que provoca la aparición de envejecimiento prematuro en quienes la padecen. No se entiende bien el mecanismo exacto por el cual los cambios bioquímicos asociados dan lugar al fenotipo envejecido. [22]
El nucléolo es la estructura más grande de las estructuras discretas, densamente teñidas y sin membrana, conocidas como cuerpos nucleares, que se encuentran en el núcleo. Se forma alrededor de repeticiones en tándem de ADNr , ADN que codifica para el ARN ribosómico (ARNr). Estas regiones se denominan regiones organizadoras del nucleolo (NOR). Las principales funciones del nucléolo son sintetizar ARNr y ensamblar ribosomas . La cohesión estructural del nucléolo depende de su actividad, ya que el ensamblaje ribosómico en el nucléolo da como resultado la asociación transitoria de los componentes nucleolares, lo que facilita un mayor ensamblaje ribosómico y, por lo tanto, una mayor asociación. Este modelo está respaldado por observaciones de que la inactivación del ADNr da como resultado la entremezcla de estructuras nucleolares. [23]
En el primer paso del ensamblaje de los ribosomas, una proteína llamada ARN polimerasa I transcribe el ADNr, que forma un gran precursor de pre-ARNr. Este se divide en dos grandes subunidades de ARNr : 5.8S y 28S , y una pequeña subunidad de ARNr 18S . [10] : 328 [24] La transcripción, el procesamiento postranscripcional y el ensamblaje del ARNr se producen en el nucléolo, con la ayuda de pequeñas moléculas de ARN nucleolar (ARNsno), algunas de las cuales se derivan de intrones empalmados de ARN mensajeros que codifican genes relacionados con la función ribosómica. Las subunidades ribosómicas ensambladas son las estructuras más grandes que pasan a través de los poros nucleares . [1] : 526
Cuando se observa bajo el microscopio electrónico , se puede ver que el nucléolo consta de tres regiones distinguibles: los centros fibrilares (FC) más internos, rodeados por el componente fibrilar denso (DFC) (que contiene fibrilarina y nucleolina ), que a su vez está bordeado por el componente granular (GC) (que contiene la proteína nucleofosmina ). La transcripción del ADNr ocurre en el FC o en el límite FC-DFC y, por lo tanto, cuando aumenta la transcripción del ADNr en la célula, se detectan más FC. La mayor parte de la escisión y modificación de los ARNr ocurre en el DFC, mientras que los últimos pasos que involucran el ensamblaje de proteínas en las subunidades ribosómicas ocurren en el GC. [24]
Las motas son estructuras subnucleares enriquecidas con factores de empalme de ARN mensajero previo y se encuentran en las regiones intercromatinales del nucleoplasma de las células de los mamíferos. [25] A nivel de microscopio de fluorescencia aparecen como estructuras irregulares y puntiformes, que varían en tamaño y forma, y cuando se examinan con microscopio electrónico se ven como grupos de gránulos intercromatinales . Las motas son estructuras dinámicas, y tanto sus componentes proteicos como de ARN-proteína pueden ciclar continuamente entre las motas y otras ubicaciones nucleares, incluidos los sitios de transcripción activos. Las motas pueden funcionar con p53 como potenciadores de la actividad genética para mejorar directamente la actividad de ciertos genes. Además, los objetivos genéticos p53 asociados y no asociados a las motas son funcionalmente distintos. [26]
Los estudios sobre la composición, la estructura y el comportamiento de las motas han proporcionado un modelo para comprender la compartimentación funcional del núcleo y la organización de la maquinaria de expresión génica [27] empalmando snRNP [28] [29] y otras proteínas de empalme necesarias para el procesamiento del pre-ARNm. [27] Debido a los requisitos cambiantes de una célula, la composición y la ubicación de estos cuerpos cambia de acuerdo con la transcripción y regulación del ARNm a través de la fosforilación de proteínas específicas. [30] Las motas de empalme también se conocen como motas nucleares (motas nucleares), compartimentos de factores de empalme (compartimentos SF), cúmulos de gránulos intercromatínicos (IGC) y snurposomas B. [ 31] Los snurposomas B se encuentran en los núcleos de los ovocitos de anfibios y en los embriones de Drosophila melanogaster . Los snurposomas B aparecen solos o unidos a los cuerpos de Cajal en las micrografías electrónicas de los núcleos de anfibios. [32] Si bien originalmente se pensaba que las motas nucleares eran sitios de almacenamiento para los factores de empalme [33] , un estudio más reciente demostró que organizar genes y sustratos de pre-ARNm cerca de las motas aumenta la eficiencia cinética del empalme de pre-ARNm, lo que en última instancia aumenta los niveles de proteína mediante la modulación del empalme [34] .

Un núcleo contiene típicamente entre una y diez estructuras compactas llamadas cuerpos de Cajal o cuerpos enrollados (CB), cuyo diámetro mide entre 0,2 μm y 2,0 μm dependiendo del tipo celular y la especie. [35] Cuando se observan bajo un microscopio electrónico, se parecen a bolas de hilo enredado [36] y son focos densos de distribución para la proteína coilina . [37] Los CB están involucrados en una serie de funciones diferentes relacionadas con el procesamiento del ARN, específicamente la maduración del ARN nucleolar pequeño (snoRNA) y el ARN nuclear pequeño (snRNA), y la modificación del ARNm de histonas. [35]
Similares a los cuerpos de Cajal están los cuerpos de Cajal de Géminis, o gemas, cuyo nombre se deriva de la constelación de Géminis en referencia a su estrecha relación "gemela" con los CB. Las gemas son similares en tamaño y forma a los CB, y de hecho son virtualmente indistinguibles bajo el microscopio. [37] A diferencia de los CB, las gemas no contienen ribonucleoproteínas nucleares pequeñas (snRNP), pero sí contienen una proteína llamada supervivencia de la neurona motora (SMN) cuya función se relaciona con la biogénesis de snRNP. Se cree que las gemas ayudan a los CB en la biogénesis de snRNP, [38] aunque también se ha sugerido a partir de evidencia microscópica que los CB y las gemas son diferentes manifestaciones de la misma estructura. [37] Estudios ultraestructurales posteriores han demostrado que las gemas son gemelas de los cuerpos de Cajal con la diferencia en el componente coilina; los cuerpos de Cajal son SMN positivos y coilina positivos, y las gemas son SMN positivos y coilina negativos. [39]
Además de los cuerpos nucleares descritos por primera vez por Santiago Ramón y Cajal (por ejemplo, nucléolo, motas nucleares, cuerpos de Cajal), el núcleo contiene otros cuerpos nucleares, como la asociación cariosomal en interfase polimórfica (PIKA), los cuerpos de leucemia promielocítica (PML) y las paraspeckles . Aunque se sabe poco sobre varios de estos dominios, son importantes porque muestran que el nucleoplasma no es una mezcla uniforme, sino que contiene subdominios funcionales organizados. [41]
Otras estructuras subnucleares aparecen como parte de procesos patológicos anormales. Por ejemplo, se ha informado de la presencia de pequeñas varillas intranucleares en algunos casos de miopatía nemalínica . Esta afección suele ser consecuencia de mutaciones en la actina , y las propias varillas están formadas por actina mutante, así como por otras proteínas del citoesqueleto. [43]
Los dominios PIKA, o asociaciones cariosomales interfásicas polimórficas, se describieron por primera vez en estudios de microscopía en 1991. Su función sigue sin estar clara, aunque no se pensaba que estuvieran asociados con la replicación activa del ADN, la transcripción o el procesamiento del ARN. [44] Se ha descubierto que a menudo se asocian con dominios discretos definidos por la localización densa del factor de transcripción PTF, que promueve la transcripción del ARN nuclear pequeño (snRNA). [45]
Las proteínas leucémicas promielocíticas (cuerpos nucleares PML) son cuerpos esféricos que se encuentran dispersos por todo el nucleoplasma y miden alrededor de 0,1 a 1,0 μm. Se conocen con otros nombres, como dominio nuclear 10 (ND10), cuerpos de Kremer y dominios oncogénicos PML. [46] Los cuerpos nucleares PML reciben su nombre de uno de sus componentes principales, la proteína leucémica promielocítica (PML). A menudo se observan en el núcleo en asociación con los cuerpos de Cajal y los cuerpos de escisión. [41] Los ratones Pml-/-, que no pueden crear cuerpos nucleares PML, se desarrollan normalmente sin efectos nocivos obvios, lo que demuestra que los cuerpos nucleares PML no son necesarios para la mayoría de los procesos biológicos esenciales. [47]
Descubiertos por Fox et al. en 2002, los paraspeckles son compartimentos de forma irregular en el espacio intercromatínico del núcleo. [48] Documentados por primera vez en células HeLa, donde generalmente hay entre 10 y 30 por núcleo, [49] ahora se sabe que los paraspeckles también existen en todas las células primarias humanas, líneas celulares transformadas y secciones de tejido. [50] Su nombre se deriva de su distribución en el núcleo; "para" es la abreviatura de paralelo y "speckles" se refiere a las motas de empalme a las que siempre están en estrecha proximidad. [49]
Los paraspeckles secuestran proteínas nucleares y ARN y, por lo tanto, parecen funcionar como una esponja molecular [51] que está involucrada en la regulación de la expresión génica. [52] Además, los paraspeckles son estructuras dinámicas que se alteran en respuesta a cambios en la actividad metabólica celular. Dependen de la transcripción [48] y en ausencia de transcripción de la ARN Pol II, el paraspeckle desaparece y todos sus componentes proteicos asociados (PSP1, p54nrb, PSP2, CFI(m)68 y PSF) forman una tapa perinucleolar en forma de medialuna en el nucléolo. Este fenómeno se demuestra durante el ciclo celular. En el ciclo celular , los paraspeckles están presentes durante la interfase y durante toda la mitosis excepto la telofase . Durante la telofase, cuando se forman los dos núcleos hijos, no hay transcripción de la ARN Pol II , por lo que los componentes proteicos forman una tapa perinucleolar. [50]
Las fibrillas de pericromatina son visibles únicamente con microscopio electrónico. Están ubicadas junto a la cromatina transcripcionalmente activa y se cree que son los sitios de procesamiento activo del pre-ARNm . [33]
Los clastosomas son cuerpos nucleares pequeños (0,2-0,5 μm) descritos como que tienen una forma de anillo grueso debido a la cápsula periférica alrededor de estos cuerpos. [40] Este nombre se deriva del griego klastos , roto y soma , cuerpo. [40] Los clastosomas no suelen estar presentes en células normales, lo que los hace difíciles de detectar. Se forman en condiciones proteolíticas altas dentro del núcleo y se degradan una vez que hay una disminución en la actividad o si las células se tratan con inhibidores del proteasoma . [40] [53] La escasez de clastosomas en las células indica que no son necesarios para la función del proteasoma . [54] También se ha demostrado que el estrés osmótico causa la formación de clastosomas. [55] Estos cuerpos nucleares contienen subunidades catalíticas y reguladoras del proteasoma y sus sustratos, lo que indica que los clastosomas son sitios para degradar proteínas. [54]
El núcleo proporciona un sitio para la transcripción genética que está separado del lugar de la traducción en el citoplasma, lo que permite niveles de regulación genética que no están disponibles para los procariotas . La función principal del núcleo celular es controlar la expresión genética y mediar la replicación del ADN durante el ciclo celular. [1] : 171
La envoltura nuclear permite controlar el contenido nuclear y lo separa del resto del citoplasma cuando es necesario. Esto es importante para controlar los procesos a ambos lados de la membrana nuclear: en la mayoría de los casos en los que es necesario restringir un proceso citoplasmático, se retira un participante clave al núcleo, donde interactúa con factores de transcripción para regular negativamente la producción de ciertas enzimas en la vía. Este mecanismo regulador ocurre en el caso de la glucólisis , una vía celular para descomponer la glucosa para producir energía. La hexoquinasa es una enzima responsable del primer paso de la glucólisis, formando glucosa-6-fosfato a partir de glucosa. En altas concentraciones de fructosa-6-fosfato , una molécula hecha más tarde a partir de glucosa-6-fosfato, una proteína reguladora retira la hexoquinasa al núcleo, [56] donde forma un complejo represor transcripcional con proteínas nucleares para reducir la expresión de genes involucrados en la glucólisis. [57]
Para controlar qué genes se están transcribiendo, la célula separa algunas proteínas de factores de transcripción responsables de regular la expresión génica del acceso físico al ADN hasta que son activadas por otras vías de señalización. Esto evita incluso niveles bajos de expresión génica inapropiada. Por ejemplo, en el caso de los genes controlados por NF-κB , que están involucrados en la mayoría de las respuestas inflamatorias , la transcripción se induce en respuesta a una vía de señalización como la iniciada por la molécula de señalización TNF-α , que se une a un receptor de membrana celular, lo que resulta en el reclutamiento de proteínas de señalización y, finalmente, activa el factor de transcripción NF-κB. Una señal de localización nuclear en la proteína NF-κB permite que se transporte a través del poro nuclear y hacia el núcleo, donde estimula la transcripción de los genes diana. [15]
La compartimentación permite que la célula impida la traducción del ARNm no empalmado. [58] El ARNm eucariota contiene intrones que deben eliminarse antes de traducirse para producir proteínas funcionales. El empalme se realiza dentro del núcleo antes de que los ribosomas puedan acceder al ARNm para su traducción. Sin el núcleo, los ribosomas traducirían el ARNm recién transcrito (sin procesar), lo que daría lugar a proteínas malformadas y no funcionales. [1] : 108–15
La función principal del núcleo celular es controlar la expresión génica y mediar la replicación del ADN durante el ciclo celular. [1] : 171 Se ha descubierto que la replicación ocurre de forma localizada en el núcleo celular. En la fase S de la interfase del ciclo celular, tiene lugar la replicación. Contrariamente a la visión tradicional de mover las horquillas de replicación a lo largo del ADN estancado, surgió un concepto de fábricas de replicación , lo que significa que las horquillas de replicación se concentran hacia algunas regiones "fábricas" inmovilizadas a través de las cuales pasan las hebras de ADN molde como cintas transportadoras. [59]
La expresión génica implica primero la transcripción, en la que el ADN se utiliza como plantilla para producir ARN. En el caso de los genes que codifican proteínas, el ARN producido a partir de este proceso es el ARN mensajero (ARNm), que luego debe ser traducido por los ribosomas para formar una proteína. Como los ribosomas se encuentran fuera del núcleo, el ARNm producido debe ser exportado. [60]
Dado que el núcleo es el sitio de la transcripción, también contiene una variedad de proteínas que median directamente la transcripción o están involucradas en la regulación del proceso. Estas proteínas incluyen helicasas , que desenrollan la molécula de ADN de doble cadena para facilitar el acceso a ella, ARN polimerasas , que se unen al promotor de ADN para sintetizar la molécula de ARN en crecimiento, topoisomerasas , que cambian la cantidad de superenrollamiento en el ADN, ayudándolo a enrollarse y desenrollarse, así como una gran variedad de factores de transcripción que regulan la expresión. [61]
Las moléculas de ARNm recién sintetizadas se conocen como transcripciones primarias o pre-ARNm. Deben sufrir una modificación postranscripcional en el núcleo antes de ser exportadas al citoplasma; el ARNm que aparece en el citoplasma sin estas modificaciones se degrada en lugar de usarse para la traducción de proteínas. Las tres modificaciones principales son la protección de 5' , la poliadenilación de 3' y el empalme de ARN . Mientras está en el núcleo, el pre-ARNm se asocia con una variedad de proteínas en complejos conocidos como partículas de ribonucleoproteína heterogénea (hnRNP). La adición de la protección de 5' ocurre de manera cotranscripcional y es el primer paso en la modificación postranscripcional. La cola de poliadenina de 3' solo se agrega después de que se completa la transcripción. [1] : 509–18
El empalme de ARN, llevado a cabo por un complejo llamado espliceosoma , es el proceso por el cual los intrones, o regiones de ADN que no codifican proteínas, se eliminan del pre-ARNm y los exones restantes se conectan para volver a formar una única molécula continua. Este proceso normalmente ocurre después del capping 5' y la poliadenilación 3' pero puede comenzar antes de que se complete la síntesis en transcripciones con muchos exones. [1] : 494 Muchos pre-ARNm se pueden empalmar de múltiples maneras para producir diferentes ARNm maduros que codifican diferentes secuencias de proteínas . Este proceso se conoce como empalme alternativo y permite la producción de una gran variedad de proteínas a partir de una cantidad limitada de ADN. [62]
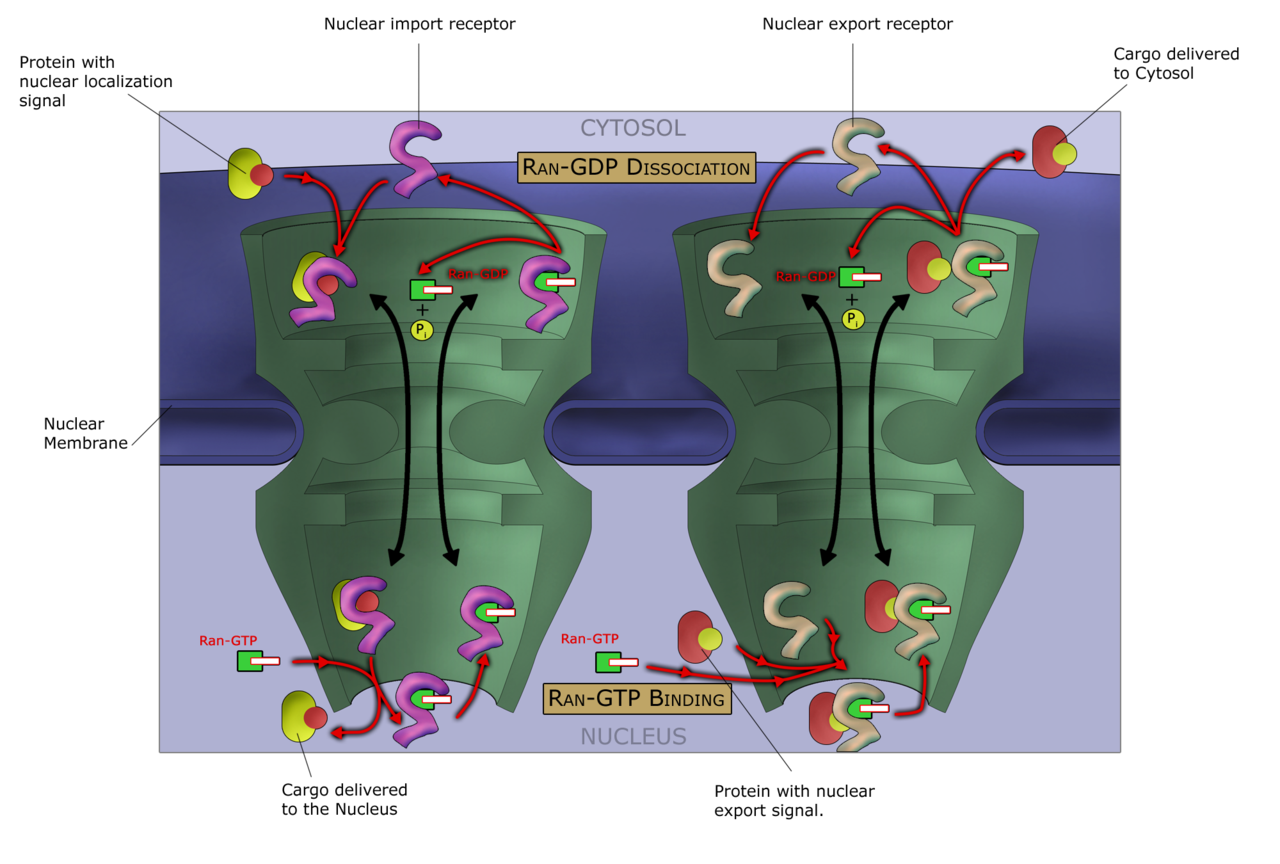
La entrada y salida de moléculas grandes del núcleo está estrechamente controlada por los complejos de poros nucleares. Aunque las moléculas pequeñas pueden entrar en el núcleo sin regulación, [63] las macromoléculas como el ARN y las proteínas requieren carioferinas de asociación llamadas importinas para entrar en el núcleo y exportinas para salir. Las proteínas de "carga" que deben ser translocadas del citoplasma al núcleo contienen secuencias cortas de aminoácidos conocidas como señales de localización nuclear , que están unidas por las importinas, mientras que las transportadas desde el núcleo al citoplasma llevan señales de exportación nuclear unidas por las exportinas. La capacidad de las importinas y exportinas para transportar su carga está regulada por GTPasas , enzimas que hidrolizan la molécula guanosina trifosfato (GTP) para liberar energía. La GTPasa clave en el transporte nuclear es Ran , que está unida a GTP o GDP (guanosina difosfato), dependiendo de si está ubicada en el núcleo o en el citoplasma. Mientras que las importinas dependen de RanGTP para disociarse de su carga, las exportinas requieren RanGTP para unirse a su carga. [14]
La importación nuclear depende de que la importina se una a su carga en el citoplasma y la transporte a través del poro nuclear hasta el núcleo. Dentro del núcleo, RanGTP actúa para separar la carga de la importina, lo que permite que la importina salga del núcleo y se reutilice. La exportación nuclear es similar, ya que la exportina se une a la carga dentro del núcleo en un proceso facilitado por RanGTP, sale a través del poro nuclear y se separa de su carga en el citoplasma. [64]
Existen proteínas de exportación especializadas para la translocación de ARNm y ARNt maduros al citoplasma una vez que se ha completado la modificación postranscripcional. Este mecanismo de control de calidad es importante debido al papel central de estas moléculas en la traducción de proteínas. La expresión incorrecta de una proteína debido a la escisión incompleta de exones o la incorporación incorrecta de aminoácidos podría tener consecuencias negativas para la célula; por lo tanto, el ARN modificado de forma incompleta que llega al citoplasma se degrada en lugar de usarse en la traducción. [1]

Durante su vida, un núcleo puede descomponerse o destruirse, ya sea en el proceso de división celular o como consecuencia de la apoptosis (el proceso de muerte celular programada ). Durante estos eventos, los componentes estructurales del núcleo (la envoltura y la lámina) pueden degradarse sistemáticamente. En la mayoría de las células, el desmontaje de la envoltura nuclear marca el final de la profase de la mitosis. Sin embargo, este desmontaje del núcleo no es una característica universal de la mitosis y no ocurre en todas las células. Algunos eucariotas unicelulares (por ejemplo, las levaduras) experimentan la llamada mitosis cerrada , en la que la envoltura nuclear permanece intacta. En la mitosis cerrada, los cromosomas hijos migran a polos opuestos del núcleo, que luego se divide en dos. Las células de eucariotas superiores, sin embargo, generalmente experimentan mitosis abierta , que se caracteriza por la ruptura de la envoltura nuclear. Los cromosomas hijos luego migran a polos opuestos del huso mitótico y los nuevos núcleos se reensamblan alrededor de ellos. [1] : 854
En un momento determinado del ciclo celular en la mitosis abierta, la célula se divide para formar dos células. Para que este proceso sea posible, cada una de las nuevas células hijas debe tener un conjunto completo de genes, un proceso que requiere la replicación de los cromosomas, así como la segregación de los conjuntos separados. Esto ocurre cuando los cromosomas replicados, las cromátidas hermanas , se unen a los microtúbulos , que a su vez están unidos a diferentes centrosomas . Las cromátidas hermanas pueden entonces ser arrastradas a ubicaciones separadas en la célula. En muchas células, el centrosoma se encuentra en el citoplasma, fuera del núcleo; los microtúbulos serían incapaces de unirse a las cromátidas en presencia de la envoltura nuclear. [65] Por lo tanto, en las primeras etapas del ciclo celular, comenzando en la profase y hasta alrededor de la prometafase , la membrana nuclear se desmantela. [18] Asimismo, durante el mismo período, la lámina nuclear también se desmonta, un proceso regulado por la fosforilación de las láminas por las proteínas quinasas como la proteína quinasa CDC2 . [66] Hacia el final del ciclo celular, la membrana nuclear se reforma y, aproximadamente al mismo tiempo, las láminas nucleares se vuelven a ensamblar mediante la desfosforilación de las láminas. [66]
Sin embargo, en los dinoflagelados , la envoltura nuclear permanece intacta, los centrosomas se localizan en el citoplasma y los microtúbulos entran en contacto con los cromosomas, cuyas regiones centroméricas se incorporan a la envoltura nuclear (la llamada mitosis cerrada con huso extranuclear). En muchos otros protistas (p. ej., ciliados , esporozoos ) y hongos, los centrosomas son intranucleares y su envoltura nuclear tampoco se desmonta durante la división celular. [67]
La apoptosis es un proceso controlado en el que se destruyen los componentes estructurales de la célula, lo que da como resultado la muerte de la célula. Los cambios asociados con la apoptosis afectan directamente al núcleo y su contenido, por ejemplo, en la condensación de la cromatina y la desintegración de la envoltura nuclear y la lámina. La destrucción de las redes de láminas está controlada por proteasas apoptóticas especializadas llamadas caspasas , que escinden las proteínas de la lámina y, por lo tanto, degradan la integridad estructural del núcleo. La escisión de la lámina a veces se utiliza como un indicador de laboratorio de la actividad de la caspasa en ensayos de actividad apoptótica temprana. [18] Las células que expresan láminas mutantes resistentes a la caspasa son deficientes en cambios nucleares relacionados con la apoptosis, lo que sugiere que las láminas juegan un papel en la iniciación de los eventos que conducen a la degradación apoptótica del núcleo. [18] La inhibición del ensamblaje de la lámina en sí es un inductor de la apoptosis. [68]
La envoltura nuclear actúa como una barrera que impide que los virus de ADN y ARN entren en el núcleo. Algunos virus necesitan acceder a las proteínas del interior del núcleo para replicarse y/o ensamblarse. Los virus de ADN, como el herpesvirus, se replican y ensamblan en el núcleo celular y salen por gemación a través de la membrana nuclear interna. Este proceso va acompañado del desmontaje de la lámina de la cara nuclear de la membrana interna. [18]
Inicialmente se sospechó que las inmunoglobulinas en general y los autoanticuerpos en particular no ingresaban al núcleo. Actualmente, hay evidencia de que en condiciones patológicas (por ejemplo, lupus eritematoso ) la IgG puede ingresar al núcleo. [69]
La mayoría de los tipos de células eucariotas suelen tener un solo núcleo, pero algunas no tienen núcleo, mientras que otras tienen varios. Esto puede ser resultado de un desarrollo normal, como en la maduración de los glóbulos rojos de los mamíferos , o de una división celular defectuosa. [70]
Una célula anucleada no contiene núcleo y, por lo tanto, es incapaz de dividirse para producir células hijas. La célula anucleada más conocida es el glóbulo rojo de los mamíferos, o eritrocito , que también carece de otros orgánulos como las mitocondrias, y sirve principalmente como un vaso de transporte para llevar oxígeno desde los pulmones a los tejidos del cuerpo. Los eritrocitos maduran a través de la eritropoyesis en la médula ósea , donde pierden sus núcleos, orgánulos y ribosomas. El núcleo se expulsa durante el proceso de diferenciación de un eritroblasto a un reticulocito , que es el precursor inmediato del eritrocito maduro. [71] La presencia de mutágenos puede inducir la liberación de algunos eritrocitos "micronucleados" inmaduros en el torrente sanguíneo. [72] [73] Las células anucleadas también pueden surgir de una división celular defectuosa en la que una hija carece de núcleo y la otra tiene dos núcleos.
En las plantas con flores , esta condición ocurre en los elementos del tubo criboso . [74]
Las células multinucleadas contienen múltiples núcleos. La mayoría de las especies de protozoos acantáreos [75] y algunos hongos en micorrizas [76] tienen células multinucleadas de forma natural. Otros ejemplos incluyen los parásitos intestinales del género Giardia , que tienen dos núcleos por célula. [77] Los ciliados tienen dos tipos de núcleos en una sola célula, un macronúcleo somático y un micronúcleo de línea germinal . [78] En los humanos, las células del músculo esquelético , también llamadas miocitos y sincitios , se vuelven multinucleadas durante el desarrollo; la disposición resultante de los núcleos cerca de la periferia de las células permite un espacio intracelular máximo para las miofibrillas . [1] Otras células multinucleadas en el ser humano son los osteoclastos , un tipo de célula ósea . Las células multinucleadas y binucleadas también pueden ser anormales en los humanos; Por ejemplo, las células que surgen de la fusión de monocitos y macrófagos , conocidas como células gigantes multinucleadas , a veces acompañan la inflamación [79] y también están implicadas en la formación de tumores. [80]
Se sabe que varios dinoflagelados tienen dos núcleos. A diferencia de otras células multinucleadas, estos núcleos contienen dos linajes distintos de ADN: uno del dinoflagelado y el otro de una diatomea simbiótica . [81]
El origen evolutivo del núcleo, la característica principal definitoria de la célula eucariota, ha sido objeto de mucha especulación. Se han propuesto cuatro hipótesis principales para explicar la existencia del núcleo, aunque ninguna ha obtenido aún un apoyo generalizado. [82] [83] [84]
El primer modelo, conocido como "modelo sintrófico", propone que una relación simbiótica entre las arqueas y las bacterias creó la célula eucariota que contiene el núcleo. (Los organismos de los dominios Archaeal y Bacterial no tienen núcleo celular. [85] ) Se plantea la hipótesis de que la simbiosis se originó cuando las arqueas antiguas similares a las arqueas metanogénicas modernas , invadieron y vivieron dentro de bacterias similares a las mixobacterias modernas , formando finalmente el núcleo primitivo. Esta teoría es análoga a la teoría aceptada para el origen de las mitocondrias y los cloroplastos eucariotas , que se cree que se desarrollaron a partir de una relación endosimbiótica similar entre protoeucariotas y bacterias aeróbicas. [86] Una posibilidad es que la membrana nuclear surgiera como un nuevo sistema de membrana tras el origen de las mitocondrias en un huésped arqueobacteriano . [87] La membrana nuclear puede haber servido para proteger el genoma de las especies reactivas de oxígeno dañinas producidas por las protomitocondrias. [88] El origen arqueológico del núcleo está respaldado por observaciones de que las arqueas y los eucariotas tienen genes similares para ciertas proteínas, incluidas las histonas . Las observaciones de que las mixobacterias son móviles, pueden formar complejos multicelulares y poseen quinasas y proteínas G similares a las eucariotas, respaldan un origen bacteriano para la célula eucariota. [89]
Un segundo modelo propone que las células protoeucariotas evolucionaron a partir de bacterias sin una etapa endosimbiótica. Este modelo se basa en la existencia de bacterias Planctomycetota modernas que poseen una estructura nuclear con poros primitivos y otras estructuras de membrana compartimentadas. [90] Una propuesta similar afirma que una célula similar a la eucariota, el cronocito , evolucionó primero y fagocitó arqueas y bacterias para generar el núcleo y la célula eucariota. [91]
El modelo más controvertido, conocido como eucariogénesis viral , postula que el núcleo rodeado de membrana, junto con otras características eucariotas, se originó a partir de la infección de un procariota por un virus. La sugerencia se basa en similitudes entre eucariotas y virus, como cadenas de ADN lineales, protección de ARNm y unión fuerte a proteínas (analogizando las histonas con las envolturas virales ). Una versión de la propuesta sugiere que el núcleo evolucionó en concierto con la fagocitosis para formar un " depredador " celular temprano . [92] Otra variante propone que los eucariotas se originaron a partir de arqueas tempranas infectadas por poxvirus , sobre la base de la similitud observada entre las polimerasas de ADN en los poxvirus modernos y eucariotas. [93] [94] Se ha sugerido que la cuestión no resuelta de la evolución del sexo podría estar relacionada con la hipótesis de la eucariogénesis viral. [95]
Una propuesta más reciente, la hipótesis de la exomembrana , sugiere que el núcleo se originó a partir de una única célula ancestral que desarrolló una segunda membrana celular exterior; la membrana interior que encierra la célula original se convirtió entonces en la membrana nuclear y desarrolló estructuras de poros cada vez más elaboradas para el paso de componentes celulares sintetizados internamente, como las subunidades ribosómicas. [96]


El núcleo fue el primer orgánulo que se descubrió. El dibujo más antiguo que se conserva es probablemente el del primer microscopista Antonie van Leeuwenhoek (1632-1723), quien observó un "lumen", el núcleo, en los glóbulos rojos del salmón . [97] A diferencia de los glóbulos rojos de los mamíferos, los de otros vertebrados todavía contienen núcleos. [98]
El núcleo también fue descrito por Franz Bauer en 1804 [99] y con más detalle en 1831 por el botánico escocés Robert Brown en una charla en la Sociedad Linneana de Londres . Brown estaba estudiando orquídeas bajo el microscopio cuando observó un área opaca, a la que llamó "areola" o "núcleo", en las células de la capa externa de la flor. [100] No sugirió una función potencial.
En 1838, Matthias Schleiden propuso que el núcleo desempeña un papel en la generación de células, por lo que introdujo el nombre de " citoblasto " ("constructor de células"). Creía que había observado nuevas células que se reunían alrededor de "citoblastos". Franz Meyen era un fuerte oponente de esta opinión, ya que había descrito la multiplicación de las células por división y creía que muchas células no tendrían núcleo. La idea de que las células pueden generarse de novo, por el "citoblasto" o de otra manera, contradecía el trabajo de Robert Remak (1852) y Rudolf Virchow (1855), quienes propagaron decisivamente el nuevo paradigma de que las células son generadas únicamente por células (" Omnis cellula e cellula "). La función del núcleo seguía sin estar clara. [101]
Entre 1877 y 1878, Oscar Hertwig publicó varios estudios sobre la fecundación de los huevos de erizo de mar , demostrando que el núcleo del espermatozoide entra en el ovocito y se fusiona con su núcleo. Esta fue la primera vez que se sugirió que un individuo se desarrolla a partir de una célula nucleada (única). Esto estaba en contradicción con la teoría de Ernst Haeckel de que la filogenia completa de una especie se repetiría durante el desarrollo embrionario, incluida la generación de la primera célula nucleada a partir de una "monérula", una masa sin estructura de protoplasma primordial (" Urschleim "). Por lo tanto, la necesidad del núcleo del espermatozoide para la fecundación se discutió durante bastante tiempo. Sin embargo, Hertwig confirmó su observación en otros grupos animales, incluidos los anfibios y los moluscos . Eduard Strasburger produjo los mismos resultados para las plantas en 1884. Esto allanó el camino para asignar al núcleo un papel importante en la herencia. En 1873, August Weismann postuló la equivalencia de las células germinales paternas y maternas para la herencia. La función del núcleo como portador de información genética sólo se hizo evidente más tarde, tras el descubrimiento de la mitosis y el redescubrimiento de las reglas mendelianas a principios del siglo XX; de este modo se desarrolló la teoría cromosómica de la herencia . [101]
{{cite book}}: Mantenimiento de CS1: falta la ubicación del editor ( enlace )