Ceratosaurus / ˌ s ɛr ə t oʊ ˈ s ɔːr ə s / (del griego κέρας/κέρατος, keras/keratos que significa "cuerno" y σαῦρος sauros que significa "lagarto") fue un dinosaurio terópodo carnívoro que vivió en el período Jurásico Superior ( edades Kimmeridgiense a Tithonian ). El género fue descrito por primera vez en 1884 por el paleontólogo estadounidense Othniel Charles Marsh basándose en un esqueleto casi completo descubierto en Garden Park, Colorado , en rocas pertenecientes a la Formación Morrison . La especie tipo es Ceratosaurus nasicornis .
El espécimen de Garden Park sigue siendo el esqueleto más completo conocido del género y solo se han descrito un puñado de especímenes adicionales desde entonces. Dos especies adicionales, Ceratosaurus dentisulcatus y Ceratosaurus magnicornis , fueron descritas en 2000 a partir de dos esqueletos fragmentarios de la cantera Cleveland-Lloyd de Utah y de las cercanías de Fruita, Colorado . Sin embargo, la validez de estas especies adicionales ha sido cuestionada, y los tres esqueletos posiblemente representan diferentes etapas de crecimiento de la misma especie. En 1999, se informó del descubrimiento del primer espécimen juvenil. En 2000, se excavó y describió un espécimen parcial de la Formación Lourinhã de Portugal , lo que proporcionó evidencia de la presencia del género fuera de América del Norte. También se han reportado restos fragmentarios de Tanzania , Uruguay y Suiza , aunque su asignación a Ceratosaurus actualmente no es aceptada por la mayoría de los paleontólogos .
Ceratosaurus era un terópodo de tamaño mediano. Se estima que el espécimen original medía 5,3 m (17 pies) o 5,69 m (18,7 pies) de largo, mientras que el espécimen descrito como C. dentisulcatus era más grande, de alrededor de 7 m (23 pies) de largo. Ceratosaurus se caracterizaba por tener mandíbulas profundas que sostenían dientes proporcionalmente muy largos en forma de cuchilla, un cuerno prominente en forma de cresta en la línea media del hocico y un par de cuernos sobre los ojos. Las extremidades anteriores eran muy cortas, pero seguían siendo completamente funcionales. La mano tenía cuatro dedos con garras en los tres primeros. La cola era profunda de arriba a abajo. Una fila de pequeños osteodermos (huesos de la piel) estaba presente en la mitad del cuello, la espalda y la cola. Osteodermos adicionales estaban presentes en posiciones desconocidas en el cuerpo del animal.
Ceratosaurus da su nombre a Ceratosauria , un clado de dinosaurios terópodos que divergieron tempranamente del linaje evolutivo que condujo a las aves modernas . Dentro de Ceratosauria, algunos paleontólogos propusieron que estaba más estrechamente relacionado con Genyodectes de Argentina, que comparte los dientes fuertemente alargados. El género geológicamente más antiguo Proceratosaurus de Inglaterra , aunque originalmente descrito como un presunto antecedente de Ceratosaurus , más tarde se descubrió que era un tiranosáuroide temprano. Ceratosaurus compartió su hábitat con otros grandes géneros de terópodos, incluidos Torvosaurus y Allosaurus , y se ha sugerido que estos terópodos ocupaban diferentes nichos ecológicos para reducir la competencia. Ceratosaurus puede haber cazado dinosaurios herbívoros, aunque algunos paleontólogos sugirieron que cazaba presas acuáticas como peces. El cuerno nasal probablemente no se usó como arma como lo sugirió originalmente Marsh, sino que es más probable que se usara únicamente para exhibición.
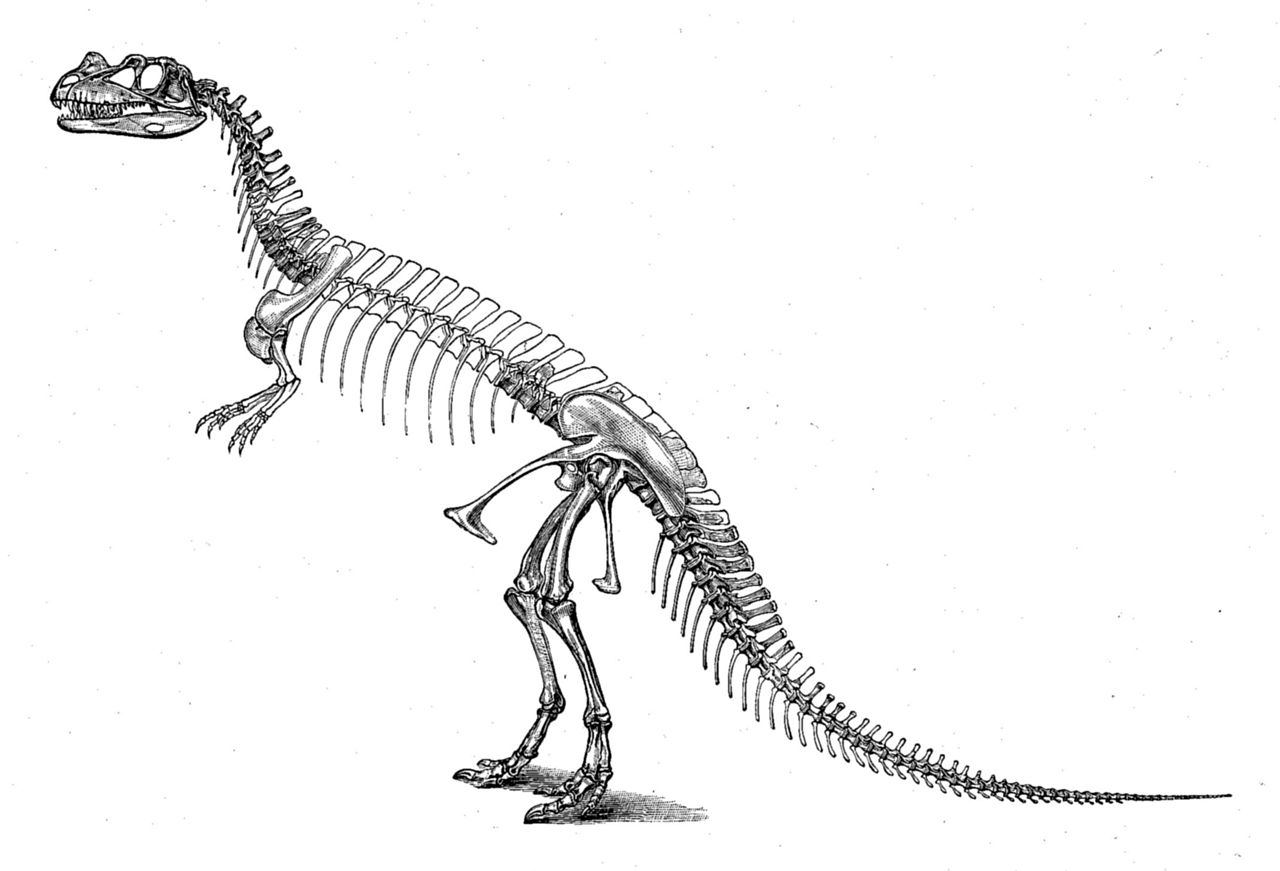
El primer espécimen, holotipo USNM 4735, fue descubierto y excavado por el granjero Marshall Parker Felch en 1883 y 1884. [1] Encontrado en articulación, con los huesos aún conectados entre sí, estaba casi completo, incluido el cráneo. Las partes faltantes significativas incluyen un número desconocido de vértebras, todas menos las últimas costillas del tronco, los húmeros (huesos de la parte superior del brazo), los huesos distales de los dedos de ambas manos, la mayor parte del brazo derecho, la mayor parte de la pierna izquierda y la mayor parte de los pies. [2] : 77 El espécimen fue encontrado encerrado en arenisca dura, lo que provocó que el cráneo y la columna vertebral se distorsionaran mucho durante la fosilización . [2] : 2, 114 El sitio del descubrimiento, ubicado en el área de Garden Park al norte de Cañon City, Colorado , y conocido como Felch Quarry 1, se considera uno de los sitios fósiles más ricos de la Formación Morrison . Se habían recuperado numerosos fósiles de dinosaurios de esta cantera incluso antes del descubrimiento de Ceratosaurus , en particular un espécimen casi completo de Allosaurus (USNM 4734) en 1883 y 1884. [2] : 7, 114
Después de la excavación, el espécimen fue enviado al Museo Peabody de Historia Natural en New Haven , donde fue estudiado por Marsh, quien lo describió como el nuevo género y especie Ceratosaurus nasicornis en 1884. [3] [2] : 114 El nombre Ceratosaurus puede traducirse como "lagarto cornudo" (de las palabras griegas κερας/κερατος , keras/keratos —"cuerno" y σαυρος / sauros —"lagarto") [4] y nasicornis con "cuerno de nariz" (de las palabras latinas nasus —"nariz" y cornu —"cuerno"). [5] Dada la integridad del espécimen, el género recién descrito fue, en ese momento, el terópodo más conocido descubierto en América. En 1898 y 1899, el espécimen fue transferido al Museo Nacional de Historia Natural en Washington, DC , junto con muchos otros fósiles descritos originalmente por Marsh. Solo una parte de este material estaba completamente preparado cuando llegó a Washington. La preparación posterior duró desde 1911 hasta fines de 1918. El embalaje y el envío desde New Haven a Washington causaron algunos daños al espécimen de Ceratosaurus . [2] : 2, 114 En 1920, Charles Gilmore publicó una redescripción extensa de este y los otros especímenes de terópodos recibidos de New Haven, incluido el espécimen casi completo de Allosaurus recuperado de la misma cantera. [2] : 2

En un artículo de 1892, Marsh publicó la primera reconstrucción esquelética de Ceratosaurus , que muestra al animal con 22 pies (6,7 m) de largo y 12 pies (3,7 m) de altura. [6] Como señaló Gilmore en 1920, el tronco se representó demasiado largo en esta reconstrucción, incorporando al menos seis vértebras dorsales de más. Este error se repitió en varias publicaciones posteriores, incluida la primera reconstrucción en vida, que fue dibujada en 1899 por Frank Bond bajo la guía de Charles R. Knight , pero no publicada hasta 1920. Una reconstrucción en vida más precisa, publicada en 1901, fue realizada por Joseph M. Gleeson , nuevamente bajo la supervisión de Knight. El holotipo fue montado por Gilmore en 1910 y 1911. Desde entonces, se exhibió en el Museo Nacional de Historia Natural. La mayoría de las reconstrucciones tempranas muestran a Ceratosaurus en una postura erguida, con la cola arrastrándose por el suelo. [2] : 115–116 El montaje del holotipo de Gilmore, en cambio, estaba muy adelantado a su tiempo. [7] : 276 Inspirado por los huesos del muslo superior, que se encontraron en ángulo contra la parte inferior de la pierna, representó el montaje como un animal corriendo con una postura horizontal y una cola que no hacía contacto con el suelo. Debido al fuerte aplanamiento de los fósiles, Gilmore montó el espécimen, no como un esqueleto independiente, sino como un bajorrelieve dentro de una pared artificial. [2] : 114 Como los huesos estaban parcialmente incrustados en una placa, el acceso científico era limitado. En el curso de la renovación de la exposición de dinosaurios del museo entre 2014 y 2019, el espécimen fue desmantelado y liberado de la placa que lo envolvía. [8] [9] En la nueva exposición, que se inauguraría en 2019, se planeó reemplazar el montaje por un molde independiente y los huesos originales se almacenarían en la colección del museo para permitir el acceso total a los científicos. [9]
.jpg/1280px-Osteology_of_the_carnivorous_Dinosauria_in_the_United_States_National_museum_BHL40623218_edited_(cropped).jpg)
Después del descubrimiento del holotipo de C. nasicornis , no se hizo un hallazgo significativo de Ceratosaurus hasta principios de la década de 1960, cuando el paleontólogo James Madsen y su equipo desenterraron un esqueleto fragmentario y desarticulado que incluía el cráneo (UMNH VP 5278) en la cantera de dinosaurios Cleveland-Lloyd de Utah. Este hallazgo representa uno de los especímenes de Ceratosaurus más grandes conocidos. [10] : 21 Un segundo espécimen articulado que incluye el cráneo (MWC 1) fue descubierto en 1976 por Thor Erikson, hijo del paleontólogo Lance Erikson, cerca de Fruita, Colorado . [4] Un espécimen bastante completo, carece de mandíbulas inferiores, antebrazos y gastralia . El cráneo, aunque razonablemente completo, se encontró desarticulado y está fuertemente aplanado lateralmente. Aunque era un individuo grande, aún no había alcanzado el tamaño adulto, como lo indican las suturas no fusionadas entre los huesos del cráneo. [10] : 2–3 Se produjeron reconstrucciones tridimensionales científicamente precisas del cráneo para su uso en exhibiciones de museos utilizando un proceso complicado que incluía el moldeado y vaciado de los huesos originales individuales, la corrección de deformidades, la reconstrucción de partes faltantes, el ensamblaje de los moldes de los huesos en su posición adecuada y la pintura para que coincidiera con el color original de los huesos. [11]
Tanto los especímenes de Fruita como de Cleveland-Lloyd fueron descritos por Madsen y Samuel Paul Welles en una monografía de 2000, y el espécimen de Utah se asignó a la nueva especie C. dentisulcatus y el espécimen de Colorado se asignó a la nueva especie C. magnicornis . [10] El nombre dentisulcatus se refiere a los surcos paralelos presentes en los lados internos de los dientes premaxilares y los primeros tres dientes de la mandíbula inferior en ese espécimen. Magnicornis apunta al cuerno nasal más grande. [10] : 2, 21 Sin embargo, la validez de ambas especies fue cuestionada en publicaciones posteriores. Brooks Britt y colegas, en 2000, afirmaron que el holotipo de C. nasicornis era de hecho un individuo juvenil, y que las dos especies más grandes representaban el estado adulto de una sola especie. [12] Oliver Rauhut, en 2003, y Matthew Carrano y Scott Sampson, en 2008, consideraron las diferencias anatómicas citadas por Madsen y Welles para apoyar que estas especies adicionales representen una variación ontogenética (relacionada con la edad) o individual. [13] [14] : 192
En 1992 se descubrió otro ejemplar (BYUVP 12893) en la cantera Agate Basin al sureste de Moore, Utah , pero aún espera una descripción. El ejemplar, considerado el más grande conocido del género, incluye la mitad frontal de un cráneo, siete vértebras dorsales pélvicas fragmentarias y una pelvis y un sacro articulados. [14] : 192 [10] : 36 En 1999, Britt informó del descubrimiento de un esqueleto de Ceratosaurus perteneciente a un individuo juvenil. Descubierto en la cantera Bone Cabin en Wyoming, es un 34% más pequeño que el holotipo de C. nasicornis y consta de un cráneo completo, así como del 30% del resto del esqueleto, incluida una pelvis completa. [15]
Además de estos cinco hallazgos esqueléticos, se han reportado restos fragmentarios de Ceratosaurus en varias localidades de las zonas estratigráficas 2 y 4-6 de la Formación Morrison, [16] incluyendo algunos de los principales yacimientos fósiles de la formación. El Monumento Nacional de los Dinosaurios , Utah, arrojó un premaxilar derecho aislado (DNM 972). Se reportó un omóplato grande (escapulocoracoides) en Como Bluff en Wyoming . Otro espécimen proviene de la cantera Dry Mesa de Colorado e incluye un escapulocoracoides izquierdo, así como fragmentos de vértebras y huesos de las extremidades. En la cantera Mygatt Moore, Colorado, el género es conocido a partir de dientes. [10] : 36
Entre 1909 y 1913, las expediciones alemanas del Museo de Naturaleza de Berlín descubrieron una diversa fauna de dinosaurios de la Formación Tendaguru en África Oriental Alemana , en lo que hoy es Tanzania . [17] Aunque comúnmente se considera la localidad de dinosaurios africana más importante, [17] los grandes dinosaurios terópodos solo se conocen a través de unos pocos y muy fragmentarios restos. [18] En 1920, el paleontólogo alemán Werner Janensch asignó varias vértebras dorsales de la cantera "TL" a Ceratosaurus , como Ceratosaurus sp. (de especie incierta). En 1925, Janensch nombró una nueva especie de Ceratosaurus , C. roechlingi , basándose en restos fragmentarios de la cantera "Mw" que abarcan un hueso cuadrado, un peroné, vértebras caudales fragmentarias y otros fragmentos. Este espécimen proviene de un individuo sustancialmente más grande que el holotipo C. nasicornis . [18]

En su monografía de 2000, Madsen y Welles confirmaron la asignación de estos hallazgos a Ceratosaurus . Además, atribuyeron varios dientes al género, que originalmente había sido descrito por Janensch como una posible especie de Labrosaurus , Labrosaurus (?) stechowi . [10] Otros autores cuestionaron la asignación de cualquiera de los hallazgos de Tendaguru a Ceratosaurus , señalando que ninguno de estos especímenes muestra características diagnósticas para ese género. [19] [20] : 66 [14] : 192 [21] En 2011, Rauhut encontró que tanto C. roechlingi como Labrosaurus (?) stechowi eran posibles ceratosáuridos, pero los encontró no diagnósticos a nivel de género y los designó como nomina dubia (nombres dudosos). [21] En 1990, Timothy Rowe y Jacques Gauthier mencionaron otra especie de Ceratosaurus de Tendaguru, Ceratosaurus ingens , que supuestamente fue erigida por Janensch en 1920 y se basó en 25 dientes aislados, muy grandes, de hasta 15 cm (5,9 pulgadas) de longitud. [19] [18] Sin embargo, Janensch asignó esta especie a Megalosaurus , no a Ceratosaurus . Por lo tanto, este nombre podría ser un simple error de copia. [10] : 37 [18] Rauhut, en 2011, demostró que Megalosaurus ingens no estaba estrechamente relacionado ni con Megalosaurus ni con Ceratosaurus , sino que posiblemente representa un carcarodontosáurido . [21]
En 2000 y 2006, los paleontólogos dirigidos por Octávio Mateus describieron un hallazgo de la Formación Lourinhã del centro-oeste de Portugal (ML 352) como un nuevo espécimen de Ceratosaurus , que consiste en un fémur derecho (hueso superior del muslo), una tibia izquierda (hueso de la espinilla) y varios dientes aislados recuperados de los acantilados de la playa de Valmitão, entre los municipios de Lourinhã y Torres Vedras . [22] [23] Los huesos se encontraron incrustados en areniscas de grano fino de color amarillo a marrón, que fueron depositadas por los ríos como depósitos de llanura aluvial y pertenecen a los niveles inferiores del Miembro Porto Novo, que se cree que es de finales del Kimmeridgiano . Huesos adicionales de este individuo (SHN (JJS)-65), incluyendo un fémur izquierdo, una tibia derecha y un peroné izquierdo parcial (hueso de la pantorrilla), fueron expuestos desde entonces debido a la erosión progresiva del acantilado . Aunque inicialmente formaban parte de una colección privada, estos elementos adicionales pasaron a estar oficialmente curados después de que la colección privada fuera donada a la Sociedade de História Natural en Torres Vedras y se describieron en detalle en 2015. [24] El espécimen fue atribuido a la especie Ceratosaurus dentisulcatus por Mateus y colegas en 2006. [23] Una revisión de 2008 realizada por Carrano y Sampson confirmó la asignación a Ceratosaurus , pero concluyó que la asignación a cualquier especie específica no es posible en la actualidad. [14] : 192 En 2015, Elisabete Malafaia y colegas, que cuestionaron la validez de C. dentisulcatus , asignaron el espécimen a Ceratosaurus aff. Ceratosaurus nasicornis . [24]
Otros informes incluyen un solo diente encontrado en Moutier , Suiza . Originalmente nombrado por Janensch en 1920 como Labrosaurus meriani , el diente fue posteriormente asignado a Ceratosaurus sp. (de especie desconocida) por Madsen y Welles. [10] : 35–36 En 2008, Matías Soto y Daniel Perea describieron dientes de la Formación Tacuarembó en Uruguay , incluyendo una presunta corona dental premaxilar. Esta muestra estrías verticales en su lado interno y carece de dentículos en su borde frontal. Estas características, en esta combinación, solo se conocen de Ceratosaurus . Los autores, sin embargo, enfatizaron que una asignación a Ceratosaurus es inviable porque los restos son escasos y señalan que la asignación del material europeo y africano a Ceratosaurus debe verse con cautela. [25] En 2020, Soto y colegas describieron dientes adicionales de Ceratosaurus de la misma formación que respaldan aún más su interpretación anterior. [26]

Ceratosaurus siguió el plan corporal típico de los grandes dinosaurios terópodos. [6] Como bípedo , se movía sobre poderosas patas, mientras que sus brazos eran de tamaño reducido. El espécimen USNM 4735, el primer esqueleto descubierto y holotipo de Ceratosaurus nasicornis , era un individuo de 5,3 m (17 pies) o 5,69 m (18,7 pies) de largo según fuentes separadas. [2] : 115 [7] No está claro si este animal estaba completamente desarrollado. [20] : 66 Othniel Charles Marsh , en 1884, sugirió que este espécimen pesaba aproximadamente la mitad que el contemporáneo Allosaurus . [3] En relatos más recientes, esto se revisó a 418 kilogramos (922 lb), 524 kg (1155 lb) o 670 kg (1480 lb). [27] Tres esqueletos adicionales descubiertos en la segunda mitad del siglo XX eran sustancialmente más grandes. El primero de ellos, UMNH VP 5278, fue estimado por James Madsen en unos 8,8 m (29 pies) de largo, [4] pero más tarde se estimó en 7 m (23 pies) de largo. [28] Su peso se calculó en 980 kg (2160 lb), 452 kg (996 lb) y 700 kg (1540 lb) en trabajos separados. [7] [28] [29] El segundo esqueleto, MWC 1, era algo más pequeño que UMNH VP 5278 y podría haber pesado 275 kg (606 lb). [29] Se afirmó que el tercer espécimen, aún no descrito, BYUVP 12893 era el más grande descubierto hasta ahora, aunque no se han publicado estimaciones. [14] : 192 Otro espécimen, ML 352, descubierto en Portugal en 2000, se estimó en 6 m (20 pies) de longitud y 600 kg (1320 libras). [28]

El cráneo era bastante grande en proporción al resto de su cuerpo. [6] Mide 55 cm (22 pulgadas) de largo en el holotipo de C. nasicornis , medido desde la punta del hocico hasta el cóndilo occipital , que se conecta a la primera vértebra cervical. [2] : 88 El ancho de este cráneo es difícil de reconstruir, ya que está muy distorsionado, y la reconstrucción de Gilmore de 1920 se encontró más tarde que era demasiado ancha. [30] Se estimó que el cráneo bastante completo del espécimen MWC 1 medía 60 cm (24 pulgadas) de largo y 16 cm (6,3 pulgadas) de ancho. Este cráneo era algo más alargado que el del holotipo. [10] : 3 La parte posterior del cráneo tenía una construcción más ligera que en algunos otros terópodos más grandes debido a las amplias aberturas del cráneo, pero las mandíbulas eran profundas para sostener los dientes proporcionalmente grandes. [7] : 277 El hueso lagrimal no sólo formaba el margen posterior de la fenestra antorbital , una gran abertura entre el ojo y la fosa nasal ósea , sino también parte de su margen superior, a diferencia de los miembros de la familia Abelisauridae relacionada . El hueso cuadrado , que estaba conectado a la mandíbula inferior en su extremo inferior para formar la articulación de la mandíbula, estaba inclinado de modo que la articulación de la mandíbula se desplazaba hacia atrás en relación con el cóndilo occipital. Esto también condujo a un ensanchamiento de la base de la fenestra temporal lateral , una gran abertura detrás de los ojos. [20] : 53
La característica más distintiva era un cuerno prominente situado en la línea media del cráneo detrás de las fosas nasales óseas, que se formó a partir de protuberancias fusionadas de los huesos nasales izquierdo y derecho . [2] : 82 Solo el núcleo óseo del cuerno se conoce a partir de fósiles. En el animal vivo, este núcleo habría sostenido una vaina queratinosa . Si bien la base del núcleo del cuerno era lisa, sus dos tercios superiores estaban arrugados y revestidos de surcos que habrían contenido vasos sanguíneos cuando estaba vivo. En el holotipo, el núcleo del cuerno mide 13 cm (5,1 pulgadas) de largo y 2 cm (0,79 pulgadas) de ancho en su base, pero rápidamente se estrecha a solo 1,2 cm (0,47 pulgadas) más arriba, y tiene 7 cm (2,8 pulgadas) de altura. [2] : 82 Es más largo y más bajo en el cráneo de MWC 1. [10] : 3 En el animal vivo, el cuerno probablemente habría sido más alargado debido a su vaina queratinosa. [31] Detrás del cuerno nasal, los huesos nasales formaban un surco ovaluro. Tanto este surco como el cuerno nasal sirven como características para distinguir a Ceratosaurus de géneros relacionados. [14] : 192 Además del gran cuerno nasal, Ceratosaurus poseía crestas óseas semicirculares más pequeñas delante de cada ojo, similares a las de Allosaurus . Estas crestas estaban formadas por los huesos lagrimales. [29] En los juveniles, los tres cuernos eran más pequeños que en los adultos y las dos mitades del núcleo del cuerno nasal aún no estaban fusionadas. [15]
_-_AMNH_-_DSC06292.JPG/1280px-Ceratosaurus_nasicornis_(cast)_-_AMNH_-_DSC06292.JPG)
Los huesos premaxilares , que formaban la punta del hocico, contenían apenas tres dientes en cada lado, menos que en la mayoría de los otros terópodos. [20] : 52 Los huesos maxilares de la mandíbula superior estaban revestidos con 15 dientes en forma de cuchilla en cada lado en el holotipo. Los primeros ocho de estos dientes eran muy largos y robustos, pero a partir del noveno diente en adelante, disminuyen gradualmente de tamaño. Como es típico de los terópodos, presentaban bordes finamente dentados , que contenían unos 10 dentículos por 5 mm (0,20 pulgadas) en el holotipo. [2] : 92 El espécimen MWC 1 solo mostró de 11 a 12 y el espécimen UMNH VP 5278 mostró 12 dientes en cada maxilar. Los dientes eran más robustos y más curvados en el último espécimen. [10] : 3, 27 En todos los especímenes, las coronas dentales de las mandíbulas superiores eran excepcionalmente largas. En el espécimen UMNH VP 5278, medían hasta 9,3 cm (3,7 pulgadas) de largo, lo que equivale a la altura mínima de la mandíbula inferior. En el holotipo, miden 7 cm (2,8 pulgadas) de largo, lo que incluso supera la altura mínima de la mandíbula inferior. En otros terópodos, solo se conoce una longitud de diente comparable del posiblemente estrechamente relacionado Genyodectes . [32] En contraste, varios miembros de Abelisauridae presentan coronas dentales muy cortas. [20] : 92 En el holotipo, cada mitad del dentario , el hueso que lleva los dientes de la mandíbula , estaba equipada con 15 dientes, que, sin embargo, están mal conservados. Ambos especímenes MWC 1 y UMNH VP 5278 muestran solo 11 dientes en cada dentario, que eran, como lo muestra el último espécimen, ligeramente más rectos y menos robustos que los de la mandíbula superior. [10] : 3, 21

El número exacto de vértebras es desconocido debido a varios huecos en la columna vertebral del holotipo Ceratosaurus nasicornis . Al menos 20 vértebras formaban el cuello y la espalda delante del sacro . En la porción media del cuello, los centros (cuerpos) de las vértebras eran tan largos como altos, mientras que en las porciones delantera y trasera del cuello, los centros eran más cortos que su altura. Las espinas neurales que proyectaban hacia arriba eran comparativamente grandes y, en las vértebras dorsales (espalda), eran tan altas como largos eran los centros vertebrales. El sacro, que consta de seis vértebras sacras fusionadas , estaba arqueado hacia arriba, con sus centros vertebrales fuertemente reducidos en altura en su porción media, como es el caso en algunos otros ceratosaurios . [20] : 55–58 La cola comprendía alrededor de 50 vértebras caudales y era aproximadamente la mitad de la longitud total del animal. En el holotipo, se estimó en 2,84 m (9,33 pies). [3] [2] : 115 La cola era profunda de arriba a abajo debido a sus altas espinas neurales y a sus alargados chevrones , huesos ubicados debajo de los centros vertebrales. Como en otros dinosaurios, contrarrestaba el cuerpo y contenía el enorme músculo caudofemoralis , que era responsable del empuje hacia adelante durante la locomoción, tirando del muslo superior hacia atrás cuando se contraía . [20] : 55–58

La escápula (omóplato) se fusionó con el coracoides , formando un solo hueso sin ninguna demarcación visible entre los dos elementos originales. [20] : 58 El holotipo de C. nasicornis fue encontrado con un brazo izquierdo articulado que incluía una mano incompleta. Aunque desarticulado durante la preparación, se había hecho un molde del fósil de antemano para documentar las posiciones relativas originales de los huesos. No se conocían huesos del carpo de ningún espécimen, lo que llevó a algunos autores a sugerir que se perdieron en el género. En un artículo de 2016, Matthew Carrano y Jonah Choiniere sugirieron que probablemente estaban presentes uno o más carpos cartilaginosos (no óseos), como lo indica un espacio presente entre los huesos del antebrazo y los metacarpianos, así como por la textura de la superficie dentro de este espacio visto en el molde. [33] A diferencia de la mayoría de los terópodos más derivados , que mostraban solo tres dígitos en cada mano (dígitos I-III), Ceratosaurus conservaba cuatro dígitos, y el dígito IV se reducía en tamaño. El primer y cuarto metacarpianos eran cortos, mientras que el segundo era ligeramente más largo que el tercero. El metacarpo y especialmente las primeras falanges eran proporcionalmente muy cortos, a diferencia de la mayoría de los otros terópodos basales . Solo las primeras falanges de los dedos II, III y IV se conservan en el holotipo. Se desconoce el número total de falanges y ungueales (huesos de las garras). La anatomía del metacarpiano I indica que originalmente también había falanges presentes en este dedo. El pes (pie) consistía en tres dedos que soportaban peso, numerados del II al IV. El dedo I, que en los terópodos suele reducirse a un espolón que no toca el suelo, no se conserva en el holotipo. Marsh, en su descripción original de 1884, asumió que este dedo se perdió en Ceratosaurus , pero Charles Gilmore , en su monografía de 1920, notó un área de inserción en el segundo metatarsiano que demostraba la presencia de este dedo. [2] : 112
Ceratosaurus , único entre los terópodos, poseía osteodermos (huesos de la piel) pequeños, alargados e irregularmente formados a lo largo de la línea media de su cuerpo. Dichos osteodermos se han encontrado por encima de las espinas neurales de las vértebras cervicales 4 y 5, así como de las vértebras caudales 4 a 10, y probablemente formaban una fila continua que podría haberse extendido desde la base del cráneo hasta la mayor parte de la cola. Como sugirió Gilmore en 1920, su posición en la matriz rocosa probablemente refleja su posición exacta en el animal actual. Los osteodermos por encima de la cola se encontraron separados de las espinas neurales por 25 mm (0,98 pulgadas) a 38 mm (1,5 pulgadas), posiblemente explicando la piel y los músculos presentes en el medio, mientras que los del cuello estaban mucho más cerca de las espinas neurales. Aparte de la línea media del cuerpo, la piel contenía osteodermos adicionales, como lo indica una placa grande, aproximadamente cuadrangular, de 58 mm (2,3 in) por 70 mm (2,8 in) encontrada junto con el holotipo. Sin embargo, se desconoce la posición de esta placa en el cuerpo. [2] : 113–114 El espécimen UMNH VP 5278 también se encontró con varios osteodermos, que se han descrito como de forma amorfa. Aunque la mayoría de estos huesecillos se encontraron a una distancia máxima de 5 m del esqueleto, no estaban directamente asociados con ninguna vértebra, a diferencia del holotipo de C. nasicornis , por lo que su posición original en el cuerpo no se puede inferir a partir de este espécimen. [10] : 32

En su descripción original del holotipo de Ceratosaurus nasicornis y publicaciones posteriores, Marsh señaló una serie de características que eran desconocidas en todos los demás terópodos conocidos en ese momento. [14] : 185 Dos de estas características, la pelvis fusionada y el metatarso fusionado, se conocían de las aves modernas y, según Marsh, demuestran claramente la estrecha relación entre estas últimas y los dinosaurios. [34] Para diferenciar el género de Allosaurus , Megalosaurus y celurosaurios , Marsh convirtió a Ceratosaurus en el único miembro de una nueva familia , Ceratosauridae , y un nuevo infraorden , Ceratosauria. [14] : 185 Esto fue cuestionado en 1892 por Edward Drinker Cope , archirrival de Marsh en las Guerras de los Huesos , quien argumentó que las características distintivas como el cuerno nasal simplemente mostraban que C. nasicornis era una especie distinta, pero eran insuficientes para justificar un género distinto. En consecuencia, asignó a C. nasicornis al género Megalosaurus , creando la nueva combinación Megalosaurus nasicornis . [35]
Aunque Ceratosaurus se mantuvo como un género distinto en todos los análisis posteriores, [2] : 76 sus relaciones siguieron siendo controvertidas durante el siglo siguiente. Tanto Ceratosauridae como Ceratosauria no fueron ampliamente aceptados, con solo unos pocos y poco conocidos miembros adicionales identificados. A lo largo de los años, autores separados clasificaron a Ceratosaurus dentro de Deinodontidae , Megalosauridae , Coelurosauria , Carnosauria y Deinodontoidea . [10] : 2 En su revisión de 1920, Gilmore argumentó que el género era el terópodo más basal conocido después del Triásico , no estando tan estrechamente relacionado con ningún otro terópodo contemporáneo conocido en ese momento. Por lo tanto, garantiza su propia familia: Ceratosauridae. [2] : 76 Sin embargo, no fue hasta el establecimiento del análisis cladístico en la década de 1980 que la afirmación original de Marsh de Ceratosauria como un grupo distinto ganó terreno. En 1985, se descubrió que los géneros sudamericanos recién descubiertos Abelisaurus y Carnotaurus estaban estrechamente relacionados con Ceratosaurus . Gauthier, en 1986, reconoció que Coelophysoidea estaba estrechamente relacionado con Ceratosaurus , aunque este clado queda fuera de Ceratosauria en los análisis más recientes. Desde entonces se han reconocido muchos miembros adicionales de Ceratosauria. [14] : 185
Ceratosauria se separó temprano de la línea evolutiva que condujo a las aves modernas y se considera basal dentro de los terópodos. [36] Ceratosauria en sí contiene un grupo de miembros derivados (no basales) de las familias Noasauridae y Abelisauridae , que están dentro del clado Abelisauroidea , así como varios miembros basales, como Elaphrosaurus , Deltadromeus y Ceratosaurus . La posición de Ceratosaurus dentro de los ceratosaurios basales está en debate. Algunos análisis consideraron a Ceratosaurus como el más derivado de los miembros basales, formando el taxón hermano de Abelisauroidea. [14] : 187 [37] Oliver Rauhut, en 2004, propuso a Genyodectes como el taxón hermano de Ceratosaurus , ya que ambos géneros se caracterizan por dientes excepcionalmente largos en la mandíbula superior. [32] Rauhut agrupó a Ceratosaurus y Genyodectes dentro de la familia Ceratosauridae, [32] lo que fue seguido por varios relatos posteriores. [38] [39] [40] [31]
En 2017, Shuo Wang y sus colegas concluyeron que Noasauridae no estaba anidado dentro de Abelisauroidea como se suponía anteriormente, sino que era más basal que Ceratosaurus . Debido a que los noasáuridos se habían utilizado como un punto fijo para definir los clados Abelisauroidea y Abelisauridae, estos clados incluirían en consecuencia muchos más taxones por definición, incluido Ceratosaurus . En un estudio posterior de 2018, Rafael Delcourt aceptó estos resultados, pero señaló que, como consecuencia, Abelisauroidea tendría que ser reemplazado por el sinónimo más antiguo Ceratosauroidea, que hasta entonces rara vez se usaba. Para Abelisauridae, Delcourt propuso una nueva definición que excluye a Ceratosaurus , lo que permite usar el nombre en su sentido tradicional. Wang y sus colegas encontraron además que Ceratosaurus y Genyodectes forman un clado con el género argentino Eoabelisaurus . [40] Delcourt utilizó el nombre Ceratosauridae para referirse a este mismo clado, y sugirió definir a Ceratosauridae como aquel que contiene todos los taxones que están más estrechamente relacionados con Ceratosaurus que con el abelisáurido Carnotaurus . [31]

El siguiente cladograma que muestra las relaciones de Ceratosaurus se basa en el análisis filogenético realizado por Diego Pol y Oliver Rauhut en 2012: [38]
Un cráneo del Jurásico Medio de Inglaterra aparentemente muestra un cuerno nasal similar al de Ceratosaurus . En 1926, Friedrich von Huene describió este cráneo como Proceratosaurus (que significa "antes de Ceratosaurus "), asumiendo que era un antecedente del Ceratosaurus del Jurásico Tardío . [41] Hoy, Proceratosaurus se considera un miembro basal de Tyrannosauroidea , un clado mucho más derivado de dinosaurios terópodos. [42] El cuerno nasal habría evolucionado independientemente en ambos géneros. [14] : 185 Oliver Rauhut y colegas, en 2010, agruparon a Proceratosaurus dentro de su propia familia, Proceratosauridae . Estos autores también notaron que el cuerno nasal está incompletamente preservado, lo que abre la posibilidad de que representara la porción más delantera de una cresta de la cabeza más extensa, como se ve en algunos otros proceratosáuridos como Guanlong . [42]
Dentro de la Formación Morrison y Lourinhã, los fósiles de Ceratosaurus se encuentran frecuentemente en asociación con los de otros grandes terópodos, incluyendo el megalosáurido Torvosaurus [43] y el alosaurido Allosaurus . La localidad de Garden Park en Colorado contenía, además de Ceratosaurus , fósiles atribuidos a Allosaurus . La cantera Dry Mesa en Colorado, así como la cantera Cleveland-Lloyd y el Monumento Nacional de los Dinosaurios en Utah, presentan, respectivamente, los restos de al menos tres grandes terópodos: Ceratosaurus , Allosaurus y Torvosaurus . [30] [44] Asimismo, Como Bluff y localidades cercanas en Wyoming contenían restos de Ceratosaurus , Allosaurus y al menos un gran megalosáurido. [45] Ceratosaurus era un elemento raro de la fauna de terópodos, ya que es superado en número por Allosaurus a una tasa promedio de 7,5 a 1 en los sitios donde co-ocurren. [46]
Varios estudios intentaron explicar cómo estas especies simpátricas podrían haber reducido la competencia directa. Donald Henderson, en 1998, argumentó que Ceratosaurus co-ocurrió con dos especies potenciales separadas de Allosaurus , que denotó como "morfos": un morfo con un hocico acortado, un cráneo alto y ancho y dientes cortos que se proyectan hacia atrás, y un morfo caracterizado por un hocico más largo, un cráneo más bajo y dientes largos y verticales. En términos generales, cuanto mayor sea la similitud entre las especies simpátricas en cuanto a su morfología, fisiología y comportamiento, más intensa será la competencia entre estas especies. Henderson llegó a la conclusión de que el morfo de hocico corto de Allosaurus ocupaba un nicho ecológico diferente tanto del morfo de hocico largo como del Ceratosaurus . El cráneo más corto en este morfo habría reducido los momentos de flexión que ocurren durante la mordedura, aumentando así la fuerza de mordida, comparable a la condición observada en los gatos. Sin embargo, Ceratosaurus y el otro morfo Allosaurus tenían cráneos con hocico largo, que son mejores en comparación con los de los perros. Los dientes más largos habrían sido utilizados como colmillos para dar mordidas rápidas y cortantes, con la fuerza de la mordida concentrada en un área más pequeña debido al cráneo más estrecho. Según Henderson, las grandes similitudes en la forma del cráneo entre Ceratosaurus y el morfo de hocico largo Allosaurus indican que estas formas participaron en una competencia directa entre sí. Por lo tanto, Ceratosaurus podría haber sido expulsado de los hábitats dominados por el morfo de hocico largo. De hecho, Ceratosaurus es muy raro en la cantera Cleveland-Lloyd, que contiene el morfo de hocico largo Allosaurus , pero parece ser más común tanto en Garden Park como en la cantera Dry Mesa, en la que coexiste con el morfo de hocico corto. [30]

Además, Henderson sugirió que Ceratosaurus podría haber evitado la competencia al preferir presas diferentes. La evolución de sus dientes extremadamente alargados podría haber sido un resultado directo de la competencia con el Allosaurus de hocico largo . Ambas especies también podrían haber preferido partes diferentes de los cadáveres cuando actuaban como carroñeros. Los dientes alargados de Ceratosaurus podrían haber servido como señales visuales que facilitaban el reconocimiento de miembros de la misma especie o para otras funciones sociales. Además, el gran tamaño de estos terópodos habría tendido a disminuir la competencia, ya que el número de presas posibles aumenta con el tamaño. [30]
Foster y Daniel Chure, en un estudio de 2006, coincidieron con Henderson en que Ceratosaurus y Allosaurus generalmente compartían los mismos hábitats y se alimentaban de los mismos tipos de presas, lo que significa que probablemente tenían diferentes estrategias de alimentación para evitar la competencia. Según estos investigadores, esto también se evidencia por las diferentes proporciones del cráneo, los dientes y los brazos. [46] Sin embargo, la distinción entre los dos morfos de Allosaurus fue cuestionada por algunos estudios posteriores. Kenneth Carpenter , en un estudio de 2010, encontró que los individuos de hocico corto de Allosaurus de la cantera Cleveland-Lloyd representan casos de variación individual extrema en lugar de un taxón separado. [47] Además, el cráneo de USNM 4734 de la localidad de Garden Park, que formó la base para el análisis de Henderson del morfo de hocico corto, se descubrió más tarde que había sido reconstruido demasiado corto. [48]
En un estudio de 2004, Robert Bakker y Gary Bir sugirieron que Ceratosaurus estaba especializado principalmente en presas acuáticas como peces pulmonados , cocodrilos y tortugas. Como lo indica un análisis estadístico de dientes caídos de 50 localidades separadas en y alrededor de Como Bluff, los dientes tanto de Ceratosaurus como de megalosáuridos eran más comunes en hábitats dentro y alrededor de fuentes de agua como llanuras aluviales húmedas , márgenes de lagos y pantanos. Ceratosaurus también se presentó ocasionalmente en localidades terrestres. Los alosáuridos, sin embargo, fueron igualmente comunes en hábitats terrestres y acuáticos. A partir de estos resultados, Bakker y Bir concluyeron que Ceratosaurus y los megalosáuridos deben haber cazado predominantemente cerca y dentro de los cuerpos de agua, y que Ceratosaurus también se alimentaba de cadáveres de dinosaurios más grandes en ocasiones. Los investigadores además notaron el cuerpo largo, bajo y flexible de Ceratosaurus y los megalosáuridos. En comparación con otros terópodos de Morrison, Ceratosaurus mostró espinas neurales más altas en las vértebras caudales más delanteras, que eran verticales en lugar de inclinadas hacia atrás. Junto con los huesos en forma de chevron profundos en la parte inferior de la cola, indican una cola profunda, "similar a la de un cocodrilo", posiblemente adaptada para nadar. Por el contrario, los alosáuridos presentan un cuerpo más corto, más alto y más rígido con patas más largas. Habrían estado adaptados para correr rápido en terreno abierto y para cazar grandes dinosaurios herbívoros como saurópodos y estegosaurios, pero como especularon Bakker y Bir, cambiaban estacionalmente a presas acuáticas cuando los grandes herbívoros estaban ausentes. [45] Sin embargo, esta teoría fue desafiada por Yun en 2019, sugiriendo que Ceratosaurus era simplemente más capaz de cazar presas acuáticas que otros terópodos de la Formación Morrison en lugar de ser completamente semiacuático. [49]

En su popular libro de 1986 The Dinosaur Heresies , Bakker argumentó que los huesos de la mandíbula superior estaban unidos de forma laxa a los huesos del cráneo circundantes, lo que permitía cierto grado de movimiento dentro del cráneo, una condición denominada kinesis craneal . Del mismo modo, los huesos de la mandíbula inferior habrían podido moverse unos contra otros y el hueso cuadrado podría oscilar hacia afuera, extendiendo la mandíbula inferior en la articulación de la mandíbula. En conjunto, estas características habrían permitido al animal ensanchar sus mandíbulas para tragar alimentos más grandes. [50] En un estudio de 2008, Casey Holliday y Lawrence Witmer reevaluaron afirmaciones similares realizadas para otros dinosaurios, concluyendo que la presencia de kinesis craneal impulsada por músculos no se puede probar para ninguna especie de dinosaurio y probablemente estaba ausente en la mayoría. [51]
Un pie púbico de Allosaurus muestra marcas de los dientes de otro terópodo, probablemente Ceratosaurus o Torvosaurus . La ubicación del hueso en el cuerpo (a lo largo del margen inferior del torso y parcialmente protegido por las piernas) y el hecho de que estaba entre los más grandes del esqueleto indica que el Allosaurus estaba siendo carroñeado. [52] Un conjunto de huesos en la cantera Mygatt-Moore del Jurásico Superior preserva una ocurrencia inusualmente alta de marcas de mordeduras de terópodos, la mayoría de las cuales pueden atribuirse a Allosaurus y Ceratosaurus , mientras que otras podrían haber sido hechas por Saurophaganax o Torvosaurus dado el tamaño de las estrías. Si bien la posición de las marcas de mordeduras en los dinosaurios herbívoros es consistente con la depredación o el acceso temprano a los restos, las marcas de mordeduras encontradas en el material de Allosaurus sugieren carroñeo, ya sea de los otros terópodos o de otro Allosaurus . La concentración inusualmente alta de marcas de mordeduras de terópodos en comparación con otros conjuntos podría explicarse ya sea por una utilización más completa de los recursos durante la estación seca por parte de los terópodos o por un sesgo de recolección en otras localidades. [53]
En 1884, Marsh consideró que el cuerno nasal de Ceratosaurus era un "arma muy poderosa" tanto para fines ofensivos como defensivos y Gilmore, en 1920, coincidió con esta interpretación. [3] : 331 [2] : 82 El uso del cuerno como arma ahora se considera generalmente improbable. [4] En 1985, David Norman creía que el cuerno "probablemente no era para protección contra otros depredadores", sino que podría haber sido utilizado para el combate intraespecífico entre ceratosaurios machos que competían por derechos de reproducción. [54] Gregory S. Paul , en 1988, sugirió una función similar e ilustró a dos Ceratosaurus involucrados en una competencia de cabezazos no letal. [7] En 1990, Rowe y Gauthier fueron más allá, sugiriendo que el cuerno nasal de Ceratosaurus "probablemente se usó solo con fines de exhibición" y no jugó ningún papel en enfrentamientos físicos. [19] Si se hubiera usado para exhibición, el cuerno probablemente habría sido de colores brillantes. [29] También se propuso una función de visualización para la fila de osteodermos que corre a lo largo de la línea media del cuerpo. [19]

Los metacarpianos y falanges fuertemente acortados de Ceratosaurus plantean la pregunta de si la mano retuvo la función de agarre asumida para otros terópodos basales. Dentro de Ceratosauria, se puede observar una reducción de la mano aún más extrema en los abelisáuridos, donde el brazo perdió su función original , [55] y en Limusaurus . En un artículo de 2016 sobre la anatomía de la mano de Ceratosaurus , Carrano y Jonah Choiniere destacaron la gran similitud morfológica de la mano con las de otros terópodos basales, sugiriendo que aún cumplía su función de agarre original, a pesar de su acortamiento. Aunque solo se conservan las primeras falanges, las segundas falanges habrían sido móviles, como lo indican las superficies articulares bien desarrolladas, y los dígitos probablemente habrían permitido un grado similar de movimiento al de otros terópodos basales. Como en otros terópodos distintos de los abelisáuridos, el primer dígito se habría girado ligeramente hacia adentro cuando se flexionaba . [33]

Marsh realizó un molde de la cavidad cerebral del holotipo bajo su supervisión, probablemente durante la preparación del cráneo, lo que le permitió concluir que el cerebro "era de tamaño mediano, pero comparativamente mucho más grande que el de los dinosaurios herbívoros". Sin embargo, los huesos del cráneo habían sido cementados posteriormente, por lo que la precisión de este molde no pudo verificarse mediante estudios posteriores. [3] [2] : 93
Un segundo cráneo bien conservado había sido encontrado con el espécimen MWC 1 en Fruita, Colorado, y fue escaneado por TC por los paleontólogos Kent Sanders y David Smith, lo que permitió reconstrucciones del oído interno , regiones macroscópicas del cerebro y senos craneales que transportan sangre fuera del cerebro. En 2005, los investigadores concluyeron que Ceratosaurus poseía una cavidad cerebral típica de los terópodos basales y similar a la de Allosaurus . Las impresiones de los bulbos olfatorios , que albergan el sentido del olfato, están bien conservadas. Si bien son similares a los de Allosaurus , eran más pequeños que en Tyrannosaurus , que se cree que estaba equipado con un sentido del olfato muy agudo. Los canales semicirculares , que son responsables del sentido del equilibrio y, por lo tanto, permiten inferencias sobre la orientación habitual de la cabeza y la locomoción, son similares a los encontrados en otros terópodos. En los terópodos, estas estructuras son generalmente conservadoras, lo que sugiere que los requisitos funcionales durante la locomoción han sido similares en todas las especies. El canal semicircular más anterior estaba agrandado, una característica que se encuentra generalmente en animales bípedos. La orientación del canal semicircular lateral indica que la cabeza y el cuello se mantenían horizontales en posición neutra. [56]
El holotipo de C. nasicornis fue encontrado con sus metatarsianos izquierdos II a IV fusionados. [57] Marsh, en 1884, dedicó un breve artículo a esta característica, en ese momento, desconocida en los dinosaurios, señalando la gran semejanza con la condición observada en las aves modernas. [34] La presencia de esta característica en Ceratosaurus se volvió controvertida en 1890, cuando Georg Baur especuló que la fusión en el holotipo era el resultado de una fractura curada . Esta afirmación fue repetida en 1892 por Cope, al tiempo que argumentaba que C. nasicornis debería clasificarse como una especie de Megalosaurus debido a las diferencias anatómicas insuficientes entre estos géneros. [35] Sin embargo, desde entonces se han descrito ejemplos de metatarsianos fusionados en dinosaurios que no son de origen patológico, incluidos taxones más basales que Ceratosaurus . [57] Osborn, en 1920, explicó que no es evidente un crecimiento óseo anormal y que la fusión es inusual, pero probablemente no patológica. [2] : 112 Ronald Ratkevich, en 1976, argumentó que esta fusión había limitado la capacidad de correr del animal, pero esta afirmación fue rechazada por Paul en 1988, quien señaló que la misma característica ocurre en muchos animales de movimiento rápido de la actualidad, incluidos los pájaros terrestres y los ungulados. [7] Un análisis de 1999 de Darren Tanke y Bruce Rothschild sugirió que la fusión era de hecho patológica, lo que confirma la afirmación anterior de Baur. [57] Otros informes de patologías incluyen una fractura por estrés en un hueso del pie asignado al género, [58] así como un diente roto de una especie no identificada de Ceratosaurus que muestra signos de mayor desgaste recibido después de la rotura. [57]

Todos los hallazgos de Ceratosaurus en América del Norte provienen de la Formación Morrison, una secuencia de rocas sedimentarias marinas y (predominantemente) aluviales poco profundas en el oeste de los Estados Unidos y la fuente más fértil de huesos de dinosaurios del continente. Según la datación radiométrica , la edad de la formación varía entre 156,3 millones de años en su base [59] y 146,8 millones de años en la parte superior, [60] lo que la ubica en las etapas Oxfordiana tardía , Kimmeridgiana y Tithoniana temprana del Jurásico tardío. Ceratosaurus es conocido a partir de los estratos Kimmeridgiano y Tithoniano de la formación. [20] : 49 La Formación Morrison se interpreta como un ambiente semiárido con estaciones húmedas y secas diferenciadas . La Cuenca Morrison se extendía desde Nuevo México hasta Alberta y Saskatchewan, y se formó cuando los precursores de la Cordillera Frontal de las Montañas Rocosas comenzaron a empujar hacia el oeste. Los depósitos de sus cuencas de drenaje orientadas al este fueron transportados por arroyos y ríos y depositados en tierras bajas pantanosas , lagos, canales fluviales y llanuras aluviales. [61] Esta formación es similar en edad a la Formación Lourinhã en Portugal y la Formación Tendaguru en Tanzania. [62]
La Formación Morrison registra un ambiente y un tiempo dominados por gigantescos dinosaurios saurópodos. [63] Otros dinosaurios conocidos de Morrison incluyen los terópodos Koparion , Stokesosaurus , Ornitholestes , Allosaurus y Torvosaurus , los saurópodos Apatosaurus , Brachiosaurus , Camarasaurus y Diplodocus , y los ornitisquios Camptosaurus , Dryosaurus , Nanosaurus , Gargoyleosaurus y Stegosaurus . [64] Allosaurus , que representó entre el 70 y el 75% de todos los especímenes de terópodos, estaba en el nivel trófico superior de la red alimentaria de Morrison . [44] Otros vertebrados que compartieron este paleoambiente incluyeron peces con aletas radiadas , ranas , salamandras , tortugas como Uluops , esfenodontos , lagartos , crocodilomorfos terrestres y acuáticos como Hoplosuchus y varias especies de pterosaurios como Harpactognathus y Mesadactylus . También son comunes las conchas de bivalvos y caracoles acuáticos . La flora del período ha sido revelada por fósiles de algas verdes , hongos , musgos , colas de caballo , cícadas , ginkgos y varias familias de coníferas . La vegetación variaba desde bosques ribereños de helechos arborescentes y helechos ( bosques de galería ) hasta sabanas de helechos con árboles ocasionales como la conífera Brachyphyllum , parecida a la Araucaria . [65]
Un espécimen parcial de Ceratosaurus indica la presencia del género en el Miembro Porto Novo portugués de la Formación Lourinhã. Muchos de los dinosaurios de la Formación Lourinhã son del mismo género que los observados en la Formación Morrison o tienen una contraparte cercana. [62] Además de Ceratosaurus , los investigadores también notaron que la presencia de Allosaurus y Torvosaurus en las rocas portuguesas se conoce principalmente del Morrison, mientras que Lourinhanosaurus hasta ahora solo se ha informado de Portugal. Los dinosaurios herbívoros del Miembro Porto Novo incluyen, entre otros, los saurópodos Dinheirosaurus y Zby , así como el estegosaurio Miragaia . [66] [22] [23] Durante el Jurásico Superior, Europa acababa de ser separada de América del Norte por el todavía estrecho Océano Atlántico . Portugal, como parte de la Península Ibérica , todavía estaba separado de otras partes de Europa. Según Mateus y sus colegas, la similitud entre las faunas de terópodos portugueses y norteamericanos indica la presencia de un puente terrestre temporal , lo que permite el intercambio de fauna. [22] [23] Malafaia y sus colegas, sin embargo, argumentaron a favor de un escenario más complejo, ya que otros grupos, como los saurópodos, las tortugas y los cocodrilos, muestran composiciones de especies claramente diferentes en Portugal y Norteamérica. Por lo tanto, la separación incipiente de estas faunas podría haber llevado al intercambio en algunas, pero a la especiación alopátrica en otros grupos. [24]
{{cite book}}: |journal=ignorado ( ayuda )