Serpina
1imv Las serpinas (del inglés, serine protease inhibitors) son una superfamilia de proteínas caracterizadas principalmente por ser inhibidores de proteasa, ampliamente distribuida en los seres vivos ya que se pueden encontrar en todos los eucariotas y en algunos procariotas.
Este inusual mecanismo de acción consiste en inhibir la proteasa de manera irreversible, puesto que al unirse ambas moléculas son sometidas a un gran cambio conformacional que altera sus sitios activos quedando inactivas en forma permanente.
[118][119][120] Estudios en las serpinas de Drosophila revelan que la Serpina-27A inhibe la proteasa de pascua(la proteasa final en el Nudel) Al igual que su rol central en los patrones embriónicos, la señalización también es importante para la respuesta inmune innata en insectos.
[130] Roles alternativos y las proteasas objetivo para las serpinas de las plantas han sido propuestos.
[133][134][135] Otras dos serpinas Arabidopsis , AtSRP2 (At2g14540) y AtSRP3 (At1g64030) parecen estar relacionadas con respuestas al daño del ADN.
Se predice que la celpina es inhibitoria y contiene dos dominion con terminales-N además del dominio de la serpina.
En contraste con las serpinas de mamíferos, estas moléculas poseen gran resistencia a la desnaturalización por calor.
[141][142] Serp1 suprime la respuesta inmune innata de TLR y permite la supervivencia del injerto cardiaco en ratas.
Específicamente, el crmA carece de la hélice-D en la misma proporción que las hélices A y E.
En la práctica esta molécula puede encontrarse en diferentes estados: activa, latente, dividida, delta y polimérica.
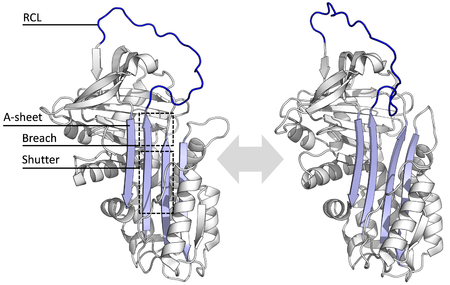
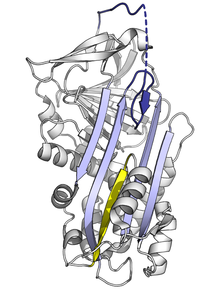
[1] Las diferentes serpinas comparten similitudes en su secuencia que varían entre un 17 a un 95%, sin embargo, su estructura posee secuencias clave comunes que permiten una conformación estable que típicamente está compuesta por tres láminas beta plegadas (denominadas A, B y C) y ocho o nueve hélices alfa (denominadas hA a hI) y un ciclo central reactivo (RCL, por el inglés reactive center loop).
[152] Las serpinas actúan como inhibidores irreversibles y suicidas al atrapar un intermediario del mecanismo catalítico de la proteasa.
[63][156][157] Aproximadamente dos tercios de las serpinas humanas realizan roles extracelulares, inhibiendo proteasas en el flujo sanguíneo para modular sus actividades.
Además muchas serpinas humanas carecen de los equivalentes funcionales precisos en organismos modelos como el ratón.
El cambio conformacional incluye que el RCL se mueva al extremo contrario de la proteína e insertando una β-plegada A, formando una hebra-β extra antiparalela.
Inicialmente, los residuos catalíticos del sitio activo de la tríada realiza un ataque nucleofílico en el enlace peptídico del sustrato, esto libera las nuevas terminales-N y forma un enlace de éster covalente entre la enzima y el sustrato.
[87] En particular, la función de serpinas inhibitorias puede ser regulada por interacciones alostéricas con los factores específicos.
Las estructuras cristalinas en rayos X de la antitrombina, el cofacto heparina II, MENT y la antiquimotripsina revelan que estas serpinas adoptan una conformación cuando en los primeros dos aminoácidos del RCL, están insertadas las β-plegadas A.
El ejemplo arquetípico de esta situación es la antitrombina, que circula en el plasma en un estado relativamente inactivo al estar parcialmente insertado.
La heparina, por lo tanto, también actúa como un templados para la unión de ambas, la proteasa y la serpina, acelerando dramáticamente la interacción entre las dos entidades.
[172] Finalmente, la terminal-N, una serpina de Thermoanaerobacter tengcongensis, se necesita para asegurar la molécula en su estado nativo inhibitorio.
Por ejemplo, ambas variantes enfermas de antitrombina wibble y wobble,[181] promueven la formación del estado latente.
En la conformación- δ, se insertan cuatro residuos del RCL en la parte superior de la β-plegada A.
[182] No está claro si otras serpinas pueden adoptar este confórmero y si esta conformación tiene un tarea funcional, pero se especula que la conformación-δ puede ser adoptada por la globulina fijadora de tiroxina durante la liberación de tiroxinas.
[99] En la ausencia de la serpina requerida, la proteasa que normalmente regularía está hiperactiva, conduciendo a patologías.
[183][26] En algunos casos raros, un solo cambio en los aminoácidos del RCL de la serpina altera su especificidad y ataca a la proteasa incorrecta.
[187][188][189][191] Varios acercamientos terapéuticos están siendo investigados para tratar la serinpatía más común, la deficiencia de antitripsina.
[195] Pequeñas moléculas también han sido diseñadas que bloquean la polimerización de antitripsina in vitro.
El cambio conformacional de S a R también ha sido aaptado por algunas serpinas que se unen para regular la afinidad por sus objetivos.
[43] Como las serpinas controlan procesos celulares tales como la coagulación y la inflamación, son proteínas blanco para investigaciones científicas.