El virus de la influenza A ( IAV ) es unpatógenoque causala gripeen aves y algunosmamíferos, incluidos los humanos.[1]Es unvirus ARNcuyos subtipos han sido aislados de aves silvestres. Ocasionalmente, se transmite de aves silvestres a domésticas, y esto puede causar enfermedades graves, brotes opandemias.[2][3][4]
Cada subtipo de virus incluye una amplia variedad de cepas con diferentes perfiles patogénicos ; algunos pueden causar enfermedades sólo en una especie pero otros en varias. Debido a que el genoma viral está segmentado, los subtipos no son cepas ni linajes, ya que la designación del subtipo se refiere a proteínas codificadas por sólo dos de los ocho segmentos del genoma.
Se ha desarrollado una vacuna contra la influenza A filtrada y purificada para humanos y muchos países la han almacenado para permitir una rápida administración a la población en caso de una pandemia de influenza aviar . En 2011, los investigadores informaron del descubrimiento de un anticuerpo eficaz contra todos los tipos de virus de la influenza A. [5]
H = hemaglutinina , una proteína que permite que el virus se una al ácido siálico y entre en la célula. Su nombre proviene del hecho de que también provoca la aglutinación de los glóbulos rojos , una característica que es fisiológicamente irrelevante para el virus in vivo .
La hemaglutinina es fundamental para que el virus reconozca y se una a las células diana, y también para que luego infecte la célula con su ARN . La neuraminidasa, por otro lado, es fundamental para la posterior liberación de las partículas del virus hijo creadas dentro de la célula infectada para que puedan propagarse a otras células. [ cita necesaria ]
Los diferentes genomas del virus de la influenza codifican diferentes proteínas hemaglutinina y neuraminidasa. Por ejemplo, el virus H5N1 designa un subtipo de influenza A que tiene una proteína hemaglutinina (H) de tipo 5 y una proteína neuraminidasa (N) de tipo 1. Hay 18 tipos conocidos de hemaglutinina y 11 tipos conocidos de neuraminidasa, por lo que, en teoría, son posibles 198 combinaciones diferentes de estas proteínas. [7] [8]
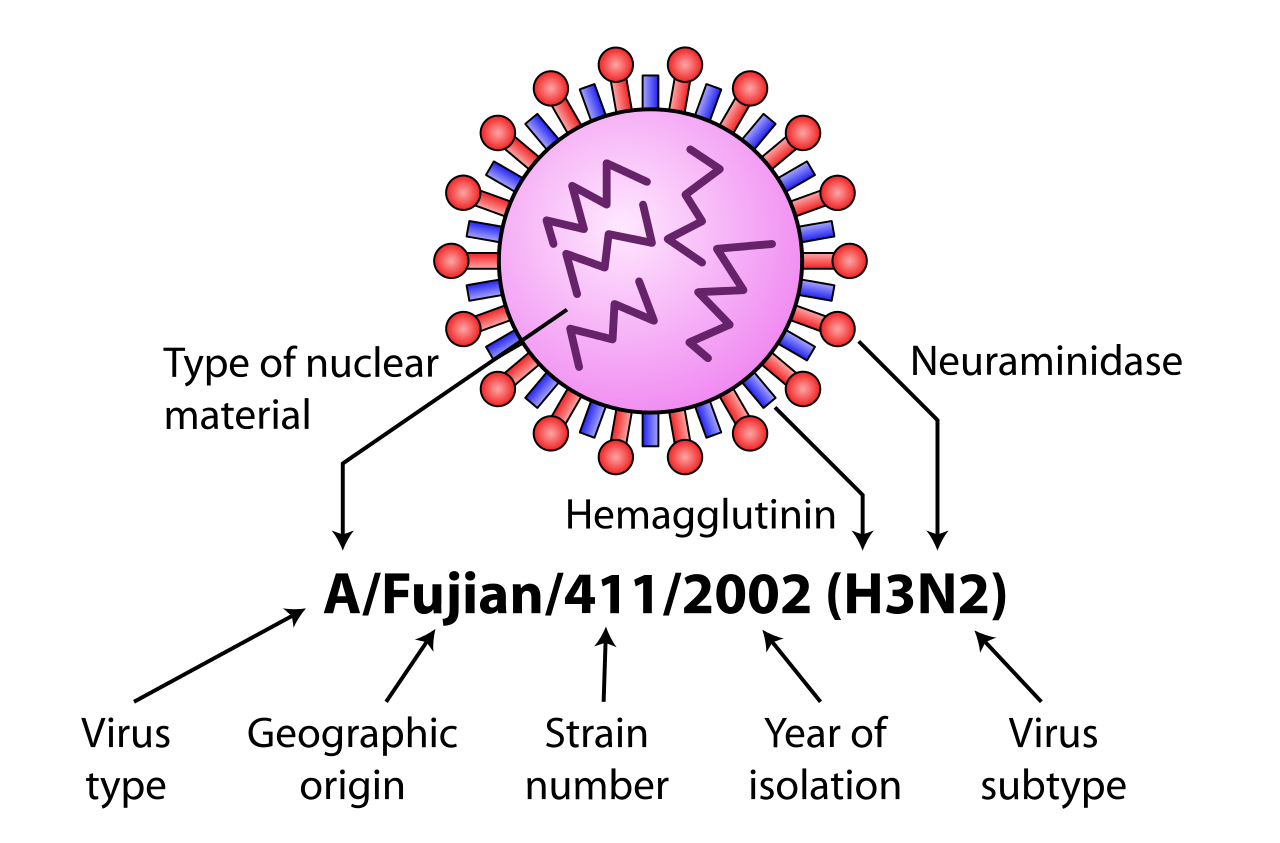
Algunas variantes se identifican y nombran según el aislado al que se parecen, por lo que se presume que comparten linaje (por ejemplo, el virus de la gripe de Fujian ); según su huésped típico (por ejemplo, el virus de la gripe humana ); según su subtipo (ejemplo H3N2); y según su letalidad (ejemplo LP, poco patógeno). Por lo tanto, una gripe causada por un virus similar al aislado A/Fujian/411/2002 (H3N2) se llama gripe de Fujian , gripe humana y gripe H3N2. [ cita necesaria ]
A veces las variantes se nombran según la especie (hospedador) en la que la cepa es endémica o a la que está adaptada. Las principales variantes nombradas usando esta convención son: [ cita necesaria ]
A veces también se han nombrado variantes según su letalidad en las aves de corral, especialmente en los pollos: [ cita necesaria ]
Influenza aviar de baja patogenicidad (LPAI)
Influenza aviar altamente patógena (HPAI), también llamada gripe mortal o gripe mortal
La mayoría de las cepas conocidas son cepas extintas. Por ejemplo, el subtipo de gripe anual H3N2 ya no contiene la cepa que provocó la gripe de Hong Kong . [ cita necesaria ]
gripe anual
La gripe anual (también llamada "gripe estacional" o "gripe humana") en los EE.UU. "provoca aproximadamente 36.000 muertes y más de 200.000 hospitalizaciones cada año. Además de este costo humano, la influenza es responsable anualmente de un costo total de más de $10 mil millones en los EE.UU." [9] A nivel mundial, el costo del virus de la influenza se estima entre 290.000 y 645.000 muertes al año, superando las estimaciones anteriores. [10]
La vacuna trivalente contra la influenza, que se actualiza anualmente, consta de componentes de glicoproteína de superficie hemaglutinina (HA) de los virus de la influenza H3N2 , H1N1 y B. [11]
La resistencia medida a los medicamentos antivirales estándar amantadina y rimantadina en H3N2 ha aumentado del 1% en 1994 al 12% en 2003 al 91% en 2005. [ cita necesaria ]
"Los virus de la gripe humana H3N2 contemporáneos son ahora endémicos en los cerdos del sur de China y pueden reagruparse con los virus aviares H5N1 en este huésped intermediario". [12]
anticuerpo FI6
FI6 , un anticuerpo que se dirige a la proteína hemaglutinina, se descubrió en 2011. FI6 es el único anticuerpo conocido eficaz contra los 16 subtipos del virus de la influenza A. [13] [14] [15]
Estructura y genética
Los virus de la influenza A son virus de ARN segmentados, monocatenarios y de sentido negativo . Los distintos subtipos están etiquetados según un número H (para el tipo de hemaglutinina ) y un número N (para el tipo de neuraminidasa ). Hay 18 antígenos H diferentes conocidos (H1 a H18) y 11 antígenos N diferentes conocidos (N1 a N11). [7] [8] El H17N10 se aisló de murciélagos frugívoros en 2012. [16] [17] El H18N11 se descubrió en un murciélago peruano en 2013. [8]
Una micrografía electrónica de transmisión (TEM) del virus de la influenza pandémica de 1918 reconstruido. La estructura inferior representa restos de membrana de las células utilizadas para amplificar el virus. [18] En la foto se muestran las partículas 'elípticas' que representan las partículas más pequeñas producidas por el virus de la influenza. Las técnicas de purificación a menudo deforman las partículas sin los protocolos de fijación adecuados, lo que da lugar a una apariencia "esférica". [19] Las partículas filamentosas o de tamaño intermedio simplemente se extienden a lo largo del eje longitudinal en el lado opuesto de los segmentos del genoma.
Los virus de la influenza tipo A son muy similares en estructura a los virus de la influenza tipo B, C y D. [20] La partícula del virus (también llamada virión) tiene entre 80 y 120 nanómetros de diámetro, de modo que los viriones más pequeños adoptan una forma elíptica. [21] [19] La longitud de cada partícula varía considerablemente, debido al hecho de que la influenza es pleomórfica y puede exceder muchas decenas de micrómetros, produciendo viriones filamentosos. [22] La confusión sobre la naturaleza de la pleomorfia del virus de la influenza surge de la observación de que las cepas adaptadas al laboratorio generalmente pierden la capacidad de formar filamentos [23] y que estas cepas adaptadas al laboratorio fueron las primeras en ser visualizadas mediante microscopía electrónica. [24] A pesar de estas formas variadas, los viriones de todos los virus de la influenza tipo A son similares en composición. Todos están formados por una envoltura viral que contiene dos tipos principales de proteínas, envueltas alrededor de un núcleo central. [25]
Las dos proteínas grandes que se encuentran en el exterior de las partículas virales son la hemaglutinina (HA) y la neuraminidasa (NA). HA es una proteína que media la unión del virión a las células diana y la entrada del genoma viral en la célula diana. La NA participa en la liberación de los abundantes sitios de unión no productivos presentes en el moco [26] , así como en la liberación de viriones descendientes de las células infectadas. [27] Estas proteínas suelen ser el objetivo de los medicamentos antivirales. [28] Además, también son las proteínas antigénicas a las que los anticuerpos del huésped pueden unirse y desencadenar una respuesta inmune. Los virus de la influenza tipo A se clasifican en subtipos según el tipo de estas dos proteínas en la superficie de la envoltura viral. Se conocen 16 subtipos de HA y 9 subtipos de NA, pero sólo H 1, 2 y 3, y N 1 y 2 se encuentran comúnmente en humanos. [29]
El núcleo central de un virión contiene el genoma viral y otras proteínas virales que empaquetan y protegen el material genético. A diferencia de los genomas de la mayoría de los organismos (incluidos humanos, animales, plantas y bacterias) que están formados por ADN bicatenario, muchos genomas virales están formados por un ácido nucleico monocatenario diferente llamado ARN. Sin embargo, algo inusual para un virus es que el genoma del virus de la influenza tipo A no es una sola pieza de ARN; en cambio, consta de trozos segmentados de ARN de sentido negativo, cada trozo contiene uno o dos genes que codifican un producto genético (proteína). [25] El término ARN de sentido negativo simplemente implica que el genoma de ARN no puede traducirse directamente en proteína; primero debe transcribirse a ARN de sentido positivo antes de poder traducirlo en productos proteicos. La naturaleza segmentada del genoma permite el intercambio de genes completos entre diferentes cepas virales. [25]
Estructura del virus de la influenza A
El genoma completo del virus de la influenza A tiene 13.588 bases de largo y está contenido en ocho segmentos de ARN que codifican al menos 10 pero hasta 14 proteínas, dependiendo de la cepa. La relevancia o presencia de productos genéticos alternativos puede variar: [30]
El segmento 1 codifica la subunidad de la ARN polimerasa (PB2).
El segmento 2 codifica la subunidad de la ARN polimerasa (PB1) y la proteína PB1-F2, que induce la muerte celular, mediante el uso de diferentes marcos de lectura del mismo segmento de ARN.
El segmento 3 codifica la subunidad de la ARN polimerasa (PA) y la proteína PA-X, que desempeña un papel en el cierre de la transcripción del huésped. [31]
El segmento 4 codifica HA (hemaglutinina). Se necesitan alrededor de 500 moléculas de hemaglutinina para formar un virión. HA determina el alcance y la gravedad de una infección viral en un organismo huésped.
El segmento 5 codifica NP, que es una nucleoproteína.
El segmento 6 codifica NA (neuraminidasa). Se necesitan alrededor de 100 moléculas de neuraminidasa para producir un virión.
El segmento 7 codifica dos proteínas de la matriz (M1 y M2) mediante el uso de diferentes marcos de lectura del mismo segmento de ARN. Se necesitan alrededor de 3.000 moléculas de proteína de matriz para producir un virión.
El segmento 8 codifica dos proteínas no estructurales distintas (NS1 y NEP) mediante el uso de diferentes marcos de lectura del mismo segmento de ARN.
Ciclo de replicación del virus de la influenza A
Los segmentos de ARN del genoma viral tienen secuencias de bases complementarias en los extremos terminales, lo que les permite unirse entre sí mediante enlaces de hidrógeno. [27] La transcripción del genoma viral con sentido (-) (ARNv) solo puede continuar después de que la proteína PB2 se une a los ARN cubiertos del huésped, lo que permite que la subunidad PA escinda varios nucleótidos después de la cubierta. Esta tapa derivada del huésped y los nucleótidos que la acompañan sirven como cebador para el inicio de la transcripción viral. La transcripción continúa a lo largo del ARNv hasta que se alcanza un tramo de varias bases de uracilo, lo que inicia una "tartamudez" mediante la cual el ARNm viral naciente se poliadenila, produciendo una transcripción madura para la exportación nuclear y la traducción por parte de la maquinaria del huésped. [32]
La síntesis de ARN tiene lugar en el núcleo celular, mientras que la síntesis de proteínas tiene lugar en el citoplasma. Una vez que las proteínas virales se ensamblan en viriones, los viriones ensamblados abandonan el núcleo y migran hacia la membrana celular. [33] La membrana de la célula huésped tiene parches de proteínas transmembrana virales (HA, NA y M2) y una capa subyacente de la proteína M1 que ayuda a los viriones ensamblados a brotar a través de la membrana, liberando virus envueltos terminados en el líquido extracelular. [33]
Se estima que los subtipos del virus de la influenza A divergieron hace 2.000 años. Se estima que los virus de la influenza A y B divergieron de un solo ancestro hace unos 4.000 años, mientras que se estima que el ancestro de los virus de la influenza A y B y el ancestro del virus de la influenza C divergieron de un ancestro común hace unos 8.000 años. [34]
Reactivación de la multiplicidad
El virus de la influenza puede sufrir una reactivación múltiple después de la inactivación por radiación UV, [35] [36] o por radiación ionizante. [37] Si alguna de las ocho cadenas de ARN que componen el genoma contiene un daño que impide la replicación o expresión de un gen esencial, el virus no es viable cuando infecta por sí solo una célula (una única infección). Sin embargo, cuando dos o más virus dañados infectan la misma célula (infección múltiple), se pueden producir una progenie de virus viables siempre que cada uno de los ocho segmentos genómicos esté presente en al menos una copia no dañada. Es decir, puede ocurrir una reactivación de la multiplicidad. [ cita necesaria ]
Tras la infección, el virus de la influenza induce una respuesta del huésped que implica una mayor producción de especies reactivas de oxígeno, y esto puede dañar el genoma del virus. [38] Si, en condiciones naturales, la supervivencia del virus es normalmente vulnerable al desafío del daño oxidativo, entonces la reactivación de la multiplicidad probablemente sea selectivamente ventajosa como una especie de proceso de reparación genómica. Se ha sugerido que la reactivación de la multiplicidad que involucra genomas de ARN segmentados puede ser similar a la forma evolucionada más temprana de interacción sexual en el mundo del ARN que probablemente precedió al mundo del ADN. [39]
Virus de la influenza humana
Cronología de las pandemias y epidemias de gripe causadas por el virus de la gripe A
El "virus de la influenza humana" generalmente se refiere a aquellos subtipos que se propagan ampliamente entre los humanos. H1N1, H1N2 y H3N2 son los únicos subtipos conocidos del virus de la influenza A que actualmente circulan entre los humanos. [40]
Los factores genéticos para distinguir entre "virus de la gripe humana" y "virus de la gripe aviar" incluyen:
PB2: (ARN polimerasa): Aminoácido (o residuo ) posición 627 en la proteína PB2 codificada por el gen de ARN PB2. Hasta el H5N1, todos los virus de la influenza aviar conocidos tenían un Glu en la posición 627, mientras que todos los virus de la influenza humana tenían una lisina .
HA: (hemaglutinina): el HA de la influenza aviar se une a los receptores de ácido siálico alfa 2–3 , mientras que el HA de la influenza humana se une a los receptores de ácido siálico alfa 2–6. Los virus de la influenza porcina tienen la capacidad de unirse a ambos tipos de receptores de ácido siálico.
Los síntomas de la gripe humana suelen incluir fiebre, tos, dolor de garganta , dolores musculares , conjuntivitis y, en casos graves, problemas respiratorios y neumonía que pueden ser mortales. La gravedad de la infección dependerá en gran parte del estado del sistema inmunológico de la persona infectada y de si la víctima ha estado expuesta anteriormente a la cepa y, por tanto, es parcialmente inmune. Los estudios de seguimiento sobre el impacto de las estatinas en la replicación del virus de la influenza muestran que el tratamiento previo de las células con atorvastatina suprime el crecimiento del virus en cultivo. [41]
La influenza aviar H5N1 altamente patógena en humanos es mucho peor: mata al 50% de los humanos que la contraen. En un caso, un niño con H5N1 experimentó diarrea seguida rápidamente de un coma sin desarrollar síntomas respiratorios o similares a los de la gripe. [42]
Los subtipos del virus de la influenza A que se han confirmado en humanos, ordenados según el número de muertes pandémicas humanas conocidas, son:
El H7N9 es responsable de una epidemia de 2013 en China [43] y el Dr. Michael Greger , autor de How Not to Die , lo considera la mayor amenaza pandémica de los virus de la influenza A [44].
El H7N7 tiene cierto potencial zoonótico : rara vez ha causado enfermedades en humanos [45] [46]
El H1N2 es actualmente endémico en cerdos y rara vez ha causado enfermedad en humanos [47]
Casos humanos y muertes causadas por diferentes subtipos del virus de la influenza AEl H1N1 fue responsable de la pandemia de 2009 tanto en poblaciones humanas como porcinas. Una variante del H1N1 fue responsable de la pandemia de gripe española que mató entre 50 y 100 millones de personas en todo el mundo durante aproximadamente un año en 1918 y 1919. [48] Otra variante fue nombrada amenaza pandémica en la pandemia de gripe de 2009 . La controversia surgió en octubre de 2005, después de que se publicara el genoma del H1N1 en la revista Science , por temor a que esta información pudiera utilizarse con fines bioterroristas . [49]
H1N2
El H1N2 es endémico en las poblaciones de cerdos [50] y se ha documentado en algunos casos humanos. [47]
H2N2
La gripe asiática, un brote pandémico de influenza aviar H2N2, se originó en China en 1957, se extendió por todo el mundo ese mismo año en el que se desarrolló una vacuna contra la influenza, duró hasta 1958 y causó entre uno y cuatro millones de muertes. [ cita necesaria ]
H3N2
H3N2 es actualmente [ ¿cuándo? ] endémico tanto en poblaciones humanas como porcinas. Evolucionó a partir del H2N2 por cambio antigénico y causó la pandemia de gripe de Hong Kong de 1968 y 1969, que mató a hasta 750.000 personas. [51] Una forma grave del virus H3N2 mató a varios niños en los Estados Unidos a finales de 2003. [52]
La cepa dominante de gripe anual en enero de 2006 fue la H3N2. La resistencia medida a los medicamentos antivirales estándar amantadina y rimantadina en el H3N2 aumentó del 1% en 1994 al 12% en 2003 y al 91% en 2005. [53] Los virus de la influenza humana H3N2 son ahora [ ¿ cuándo? ] endémico en cerdos en el sur de China, donde circulan junto con los virus aviares H5N1. [12]
El Ministerio de Salud de Japón dijo en enero de 2006 que los trabajadores de granjas avícolas en la prefectura de Ibaraki podrían haber estado expuestos al H5N2 en 2005. [54] Los títulos de anticuerpos H5N2 de sueros emparejados de 13 sujetos aumentaron cuatro veces o más. [55]
H5N8
En febrero de 2021, Rusia informó de los primeros casos conocidos de H5N8 en humanos. Se confirmó que siete personas estaban infectadas en diciembre de 2020 y desde entonces se han recuperado. [56] No hubo indicios de transmisión de persona a persona. [57]
Se encontró que una persona en Nueva York en 2003 y una persona en Virginia en 2002 tenían evidencia serológica de infección con H7N2. [ cita necesaria ] Ambos completamente recuperados. [59] [ verificación fallida ]
H7N3
En América del Norte, en febrero de 2004 se confirmó la presencia de la cepa H7N3 de influenza aviar en varias granjas avícolas de Columbia Británica. En abril de 2004, 18 granjas habían sido puestas en cuarentena para detener la propagación del virus. En esa región se han confirmado dos casos de humanos con influenza aviar. "Los síntomas incluían conjuntivitis y una enfermedad leve parecida a la gripe". [60] Ambos se recuperaron completamente.
H7N7
El H7N7 tiene un potencial zoonótico inusual. En 2003, en los Países Bajos, se confirmó que 89 personas tenían infección por el virus de la influenza H7N7 luego de un brote en aves de corral en varias granjas. Se registró una muerte.
H7N9
El 2 de abril de 2013, el Centro para la Protección de la Salud (CHP) del Departamento de Salud de Hong Kong confirmó cuatro casos más en la provincia de Jiangsu , además de los tres casos notificados inicialmente el 31 de marzo de 2013. [61] Este virus también tiene la mayor potencial de una pandemia de influenza entre todos los subtipos de influenza A. [62]
H9N2
La infección por influenza aviar A (H9N2) de baja patogenicidad se confirmó en 1999 en China y Hong Kong en dos niños, y en 2003 en Hong Kong en un niño. Los tres se recuperaron completamente. [59] [ verificación fallida ]
H10N7
En 2004, en Egipto , se informó por primera vez del virus H10N7 en humanos. Causó enfermedades en dos bebés en Egipto. El padre de uno de los niños era comerciante de aves de corral. [63]
H10N3
En mayo de 2021, en Zhenjiang , China, se informó por primera vez del virus H10N3 en humanos. Una persona resultó infectada. [64]
Evolución
Evolución genética de los virus de la influenza humana y porcina, 1918-2009
Todas las pandemias de influenza A desde [la pandemia de gripe española], y de hecho casi todos los casos de influenza A en todo el mundo (excepto las infecciones humanas por virus aviares como el H5N1 y el H7N7), han sido causadas por descendientes del virus de 1918, incluido el H1N1 "a la deriva". virus y virus H2N2 y H3N2 reagrupados. Estos últimos están compuestos por genes clave del virus de 1918, actualizados mediante genes de influenza aviar incorporados posteriormente que codifican nuevas proteínas de superficie, lo que convierte al virus de 1918 en la "madre" de todas las pandemias.
Investigadores de los Institutos Nacionales de Salud utilizaron datos del Proyecto de Secuenciación del Genoma de la Influenza y concluyeron que durante el período de diez años examinado, la mayor parte del tiempo el gen de la hemaglutinina en el H3N2 no mostró un exceso significativo de mutaciones en las regiones antigénicas, mientras que una variedad cada vez mayor de cepas acumuladas. Esto dio como resultado que una de las variantes finalmente alcanzara una mayor aptitud, se volviera dominante y, en un breve intervalo de rápida evolución , se extendiera rápidamente por la población y eliminara la mayoría de las otras variantes. [66]
En la evolución a corto plazo del virus de la influenza A, un estudio de 2006 encontró que los procesos estocásticos o aleatorios son factores clave. [67] La evolución antigénica de la HA del virus de la influenza A parece caracterizarse más por saltos puntuados y esporádicos que por una tasa constante de cambio antigénico. [68] Utilizando el análisis filogenético de 413 genomas completos de virus de influenza A humana que se recolectaron en todo el estado de Nueva York, los autores de Nelson et al. 2006 pudieron demostrar que la diversidad genética, y no la deriva antigénica, dio forma a la evolución a corto plazo de la influenza A mediante migración y reordenamiento aleatorios. La evolución de estos virus está dominada más por la importación aleatoria de cepas virales genéticamente diferentes de otras ubicaciones geográficas y menos por la selección natural. Dentro de una temporada determinada, la evolución adaptativa es poco frecuente y tuvo un efecto general débil, como lo demuestran los datos recopilados de los 413 genomas. El análisis filogenético reveló que las diferentes cepas derivaban de material genético recién importado, a diferencia de aislados que habían estado circulando en Nueva York en temporadas anteriores. Por lo tanto, el flujo de genes dentro y fuera de esta población, y no la selección natural, fue más importante a corto plazo. [ cita necesaria ]
Otros animales
Consulte H5N1 para conocer la situación actual [ ¿cuándo? ] epizoótica (una epidemia en no humanos) y panzoótica (una enfermedad que afecta a animales de muchas especies, especialmente en un área amplia) de influenza H5N1
Influenza aviar
Las aves de corral actúan como portadoras naturales asintomáticas del virus de la influenza A. Antes del actual [ ¿cuándo? ] Epizootia H5N1, se había demostrado que las cepas del virus de la influenza A se transmitían de aves silvestres únicamente a aves, cerdos, caballos, focas , ballenas y humanos; y sólo entre humanos y cerdos y entre humanos y aves domésticas; y no otros caminos como el de las aves domésticas al caballo. [69]
Las aves acuáticas silvestres son los huéspedes naturales de una gran variedad de virus de la influenza A. Ocasionalmente, los virus se transmiten de estas aves a otras especies y luego pueden causar brotes devastadores en las aves de corral domésticas o dar lugar a pandemias de influenza humana. [3] [4]
Se ha demostrado que el H5N1 se transmite a tigres, leopardos y gatos domésticos que fueron alimentados con aves domésticas (pollos) crudas con el virus. Los virus H3N8 de los caballos se cruzaron y provocaron brotes en los perros. Se han infectado ratones de laboratorio con éxito con diversos genotipos de gripe aviar. [70]
Los virus de la influenza A se propagan en el aire y en el estiércol y sobreviven más tiempo en climas fríos. También pueden transmitirse a través de alimentos, agua, equipos y ropa contaminados; sin embargo, no hay evidencia de que el virus pueda sobrevivir en carne bien cocida. Los síntomas en los animales varían, pero las cepas virulentas pueden provocar la muerte a los pocos días. Los virus de la influenza aviar que la Organización Mundial de Sanidad Animal y otros analizan para controlar las enfermedades de las aves de corral incluyen H5N1 , H7N2 , H1N7 , H7N3 , H13N6 , H5N9 , H11N6 , H3N8 , H9N2 , H5N2 , H4N8, H10N7 , H2N2 , H8N4, H14N5, H6N5 . y H12N5. [ cita necesaria ]
Brotes conocidos de gripe altamente patógena en aves de corral entre 1959 y 2003 [71]
*Brotes con importante propagación a numerosas explotaciones, provocando grandes pérdidas económicas. La mayoría de los demás brotes implicaron poca o ninguna propagación desde las granjas inicialmente infectadas.
Se registraron más de 400 muertes de focas comunes en Nueva Inglaterra entre diciembre de 1979 y octubre de 1980, por neumonía aguda causada por el virus de la influenza A/Seal/Mass/1/180 (H7N7). [72]
La gripe porcina
La influenza porcina (o "influenza porcina") se refiere a un subconjunto de Orthomyxoviridae que crean influenza y son endémicas en los cerdos. Las especies de Orthomyxoviridae que pueden causar gripe en los cerdos son el virus de la influenza A y el virus de la influenza C , pero no todos los genotipos de estas dos especies infectan a los cerdos. Los subtipos conocidos del virus de la influenza A que crean influenza y son endémicos en los cerdos son H1N1, H1N2, H3N1 y H3N2. En 1997, los virus H3N2 de humanos ingresaron a la población porcina, causando una enfermedad generalizada entre los cerdos. [73]
gripe equina
La gripe equina (o "influenza equina") se refiere a variedades del virus de la influenza A que afectan a los caballos. Los virus de la gripe equina no se aislaron hasta 1956. Los dos tipos principales de virus se llaman equino-1 (H7N7), que comúnmente afecta el músculo cardíaco del caballo, y equino-2 (H3N8), que suele ser más grave. Los virus H3N8 de los caballos han infectado a los perros. [73]
gripe canina
La gripe canina (o "influenza canina") se refiere a variedades del virus de la influenza A que afectan a los perros. En enero de 2004 se descubrió que el virus de la gripe equina H3N8 infectaba y mataba, con enfermedades respiratorias, a perros de carreras de galgos en un hipódromo de Florida.
gripe del murciélago
La gripe del murciélago (o "influenza del murciélago") se refiere a las cepas de virus de la influenza A H17N10 y H18N11 que se descubrieron en murciélagos frugívoros de América Central y del Sur, así como a un virus H9N2 aislado del murciélago frugívoro egipcio. [74] Hasta ahora no está claro si estos virus derivados de murciélagos están circulando en alguna especie que no sea murciélago y si representan una amenaza zoonótica. Sin embargo, la caracterización inicial del subtipo H18N11 sugiere que este virus de la influenza en murciélagos no está bien adaptado a otras especies que no sean los murciélagos. [75]
H3N8
El H3N8 es ahora endémico en aves, caballos y perros.
Lista de subtipos
El virus de la influenza A tiene los siguientes subtipos: [ cita necesaria ]
^ Havers FP, Campbell AJ (2020). "285. Virus de la influenza". En Kliegman RM, St Geme III JW (eds.). Nelson Libro de texto de pediatría (21ª ed.). Filadelfia: Elsevier. págs. 1727-1739. ISBN 978-0-323-56890-6.
^ ab Klenk H, Matrosovich M, Stech J (2008). "Influenza aviar: mecanismos moleculares de patogénesis y rango de huéspedes". En Mettenleiter TC, Sobrino F (eds.). Virus animales: biología molecular. Prensa académica Caister. ISBN978-1-904455-22-6.
^ ab Kawaoka Y, ed. (2006). Virología de la influenza: temas actuales. Prensa académica Caister. ISBN978-1-904455-06-6.
^ Gallagher J (29 de julio de 2011). "'El superanticuerpo combate la gripe ". Noticias de la BBC . Consultado el 29 de julio de 2011 .
^ "Taxonomía". Comité Internacional de Taxonomía de Virus (ICTV) . Consultado el 19 de julio de 2018 .
^ abc Tong S, Zhu X, Li Y, Shi M, Zhang J, Bourgeois M, Yang H, Chen X, Recuenco S, Gomez J, Chen LM, Johnson A, Tao Y, Dreyfus C, Yu W, McBride R, Carney PJ, Gilbert AT, Chang J, Guo Z, Davis CT, Paulson JC, Stevens J, Rupprecht CE, Holmes EC, Wilson IA, Donis RO (octubre de 2013). "Los murciélagos del nuevo mundo albergan diversos virus de la influenza A". Más patógenos . 9 (10): e1003657. doi : 10.1371/journal.ppat.1003657 . PMC 3794996 . PMID 24130481.
^ whitehouse.gov Archivado el 21 de febrero de 2009 en la Estrategia Nacional Wayback Machine para la Influenza Pandémica - Introducción - "Aunque se han logrado avances notables en la ciencia y la medicina durante el siglo pasado, se nos recuerda constantemente que vivimos en un universo de microbios: virus , bacterias, protozoos y hongos que cambian constantemente y se adaptan al huésped humano y a las defensas que los humanos crean. Los virus de la influenza se destacan por su resistencia y adaptabilidad. Mientras que la ciencia ha podido desarrollar vacunas y tratamientos altamente efectivos para muchas enfermedades infecciosas. que amenazan la salud pública, adquirir estas herramientas es un desafío constante con el virus de la influenza. Los cambios en la composición genética del virus requieren que desarrollemos nuevas vacunas anualmente y pronostiquemos qué cepas probablemente predominen. Después de las vacunas anuales, Estados Unidos enfrenta una carga de influenza que resulta en aproximadamente 36.000 muertes y más de 200.000 hospitalizaciones cada año. Además de este costo humano, la influenza es responsable anualmente de un costo total de más de $10 mil millones en los EE. UU. Una pandemia, o un brote mundial de un nuevo virus de influenza, podría eclipsar este impacto al abrumar nuestras capacidades médicas y de salud, lo que podría provocar cientos de miles de muertes, millones de hospitalizaciones y cientos de miles de millones de dólares en costos directos e indirectos. Esta Estrategia guiará nuestras actividades de preparación y respuesta para mitigar ese impacto".
^ Iuliano AD, Roguski KM, Chang HH, Muscatello DJ, Palekar R, Tempia S, Cohen C, Gran JM, Schanzer D, Cowling BJ, Wu P, Kyncl J, Ang LW, Park M, Redlberger-Fritz M, Yu H , Espenhain L, Krishnan A, Emukule G, van Asten L, Pereira da Silva S, Aungkulanon S, Buchholz U, Widdowson MA, Bresee JS (marzo de 2018). "Estimaciones de la mortalidad respiratoria asociada a la influenza estacional global: un estudio de modelado". Lanceta . 391 (10127): 1285–1300. doi :10.1016/s0140-6736(17)33293-2. PMC 5935243 . PMID 29248255.
^ Daum LT, Shaw MW, Klimov AI, Canas LC, Macias EA, Niemeyer D, Chambers JP, Renthal R, Shrestha SK, Acharya RP, Huzdar SP, Rimal N, Myint KS, Gould P (agosto de 2005). "Brote de influenza A (H3N2), Nepal". Enfermedades infecciosas emergentes . 11 (8): 1186–91. doi : 10.3201/eid1108.050302. PMC 3320503 . PMID 16102305. "La temporada de influenza 2003-2004 fue severa en términos de su impacto en la enfermedad debido a la circulación generalizada de virus tipo Fujian de influenza A (H3N2) antigénicamente distintos. Estos virus aparecieron por primera vez a fines de la temporada de influenza 2002-2003 y continuaron persistiendo como la cepa circulante dominante durante la siguiente temporada de influenza 2003-2004, reemplazando a los virus H3N2 similares a A/Panamá/2007/99 (1). De los 172 virus H3N2 caracterizados genéticamente por el Departamento de Defensa en 2003-2004, sólo un aislado (de Tailandia) pertenecía al linaje similar a A/Panamá. En febrero de 2003, la Organización Mundial de la Salud (OMS) cambió el componente H3N2 de la vacuna contra la influenza 2004-2005 para brindar protección contra la aparición generalizada de virus similares a Fujian (2 ). La vacuna trivalente, que se actualiza anualmente, consta de componentes de glicoproteína de superficie hemaglutinina (HA) de los virus de la influenza H3N2, H1N1 y B."
^ ab Mahmoud 2005, pág. 126 "El virus H5N1 es ahora endémico en las aves de corral en Asia (Tabla 2-1) y ha ganado un nicho ecológico arraigado desde el cual puede presentar una amenaza pandémica a largo plazo para los humanos. En la actualidad, estos virus se transmiten poco de las aves de corral a los humanos. y no hay pruebas concluyentes de transmisión de persona a persona. Sin embargo, la exposición prolongada y extensa de la población humana a los virus H5N1 aumenta la probabilidad de que los virus adquieran las características necesarias para una transmisión eficaz de persona a persona mediante mutación genética o recombinación con un virus predominante de influenza humana A. Además, los virus de influenza humana H3N2 contemporáneos ahora son endémicos en cerdos en el sur de China (Peiris et al., 2001) y pueden recombinarse con virus aviares H5N1 en este 'huésped intermedio'. Por lo tanto, es imperativo que los brotes de la enfermedad H5N1 en aves de corral en Asia se controlen de manera rápida y sostenible. Es probable que la estacionalidad de la enfermedad en las aves de corral, junto con las medidas de control ya implementadas, reduzcan temporalmente la frecuencia de los brotes de influenza H5N1 y la probabilidad de infección humana."
^ Gallagher J (29 de julio de 2011). "'El superanticuerpo combate la gripe ". Noticias de la BBC .
^ "Los científicos saludan la perspectiva de una vacuna universal contra la gripe". Independiente.co.uk . 29 de julio de 2011.
^ Chan AL (28 de julio de 2011). "Vacuna universal contra la gripe en el horizonte: los investigadores encuentran un 'superanticuerpo'". Huffpost .
^ "Nuevo virus de la gripe único encontrado en murciélagos". Opciones del NHS. 1 de marzo de 2012. Archivado desde el original el 8 de agosto de 2020 . Consultado el 16 de mayo de 2012 .
^ Tong S, Li Y, Rivailler P, Conrardy C, Castillo DA, Chen LM, Recuenco S, Ellison JA, Davis CT, York IA, Turmelle AS, Moran D, Rogers S, Shi M, Tao Y, Weil MR, Tang K, Rowe LA, Sammons S, Xu X, Frace M, Lindblade KA, Cox NJ, Anderson LJ, Rupprecht CE, Donis RO (marzo de 2012). "Un linaje distinto del virus de la influenza A de los murciélagos". Actas de la Academia Nacional de Ciencias de los Estados Unidos de América . 109 (11): 4269–74. Código Bib : 2012PNAS..109.4269T. doi : 10.1073/pnas.1116200109 . PMC 3306675 . PMID 22371588.
^ "Detalles - Biblioteca de imágenes de salud pública (PHIL)". phil.cdc.gov . Consultado el 24 de abril de 2018 .
^ ab Sugita Y, Noda T, Sagara H, Kawaoka Y (noviembre de 2011). "La ultracentrifugación deforma los viriones de influenza A no fijados". La Revista de Virología General . 92 (parte 11): 2485–93. doi :10.1099/vir.0.036715-0. PMC 3352361 . PMID 21795472.
^ Nakatsu S, Murakami S, Shindo K, Horimoto T, Sagara H, Noda T, Kawaoka Y (marzo de 2018). "Los virus de la influenza C y D contienen ocho complejos de ribonucleoproteínas organizados". Revista de Virología . 92 (6): e02084–17. doi :10.1128/jvi.02084-17. PMC 5827381 . PMID 29321324.
^ Noda T (2011). "Morfología nativa de los viriones de la influenza". Fronteras en Microbiología . 2 : 269. doi : 10.3389/fmicb.2011.00269 . PMC 3249889 . PMID 22291683.
^ Dadonaite B, Vijayakrishnan S, Fodor E, Bhella D, Hutchinson EC (agosto de 2016). "Virus filamentosos de la influenza". La Revista de Virología General . 97 (8): 1755–64. doi :10.1099/jgv.0.000535. PMC 5935222 . PMID 27365089.
^ Seladi-Schulman J, Steel J, Lowen AC (diciembre de 2013). "Los virus de la gripe esféricos tienen una ventaja de aptitud física en huevos embrionados, mientras que las cepas productoras de filamentos se seleccionan in vivo". Revista de Virología . 87 (24): 13343–53. doi :10.1128/JVI.02004-13. PMC 3838284 . PMID 24089563.
^ Mosley VM, Wyckoff RW (marzo de 1946). "Micrografía electrónica del virus de la influenza". Naturaleza . 157 (3983): 263. Bibcode :1946Natur.157..263M. doi : 10.1038/157263a0 . PMID 21016866. S2CID 6478026.
^ abc Bouvier NM, Palese P (septiembre de 2008). "La biología de los virus de la influenza". Vacuna . 26 (Suplemento 4): D49–53. doi :10.1016/j.vaccine.2008.07.039. PMC 3074182 . PMID 19230160.
^ Cohen M, Zhang XQ, Senaati HP, Chen HW, Varki NM, Schooley RT, Gagneux P (noviembre de 2013). "La influenza A penetra la mucosa del huésped escindiendo los ácidos siálicos con neuraminidasa". Revista de Virología . 10 : 321. doi : 10.1186/1743-422x-10-321 . PMC 3842836 . PMID 24261589.
^ ab Suzuki Y (marzo de 2005). "Sialobiología de la influenza: mecanismo molecular de variación del rango de huéspedes de los virus de la influenza". Boletín biológico y farmacéutico . 28 (3): 399–408. doi : 10.1248/bpb.28.399 . PMID 15744059.
^ Wilson JC, von Itzstein M (julio de 2003). "Estrategias recientes en la búsqueda de nuevas terapias antigripales". Objetivos farmacológicos actuales . 4 (5): 389–408. doi :10.2174/1389450033491019. PMID 12816348.
^ Lynch JP, Walsh EE (abril de 2007). "Influenza: estrategias en evolución en tratamiento y prevención". Seminarios de Medicina Respiratoria y de Cuidados Críticos . 28 (2): 144–58. doi :10.1055/s-2007-976487. PMID 17458769. S2CID 260316848.
^ Eisfeld AJ, Neumann G, Kawaoka Y (enero de 2015). "En el centro: ribonucleoproteínas del virus de la influenza A". Reseñas de la naturaleza. Microbiología . 13 (1): 28–41. doi :10.1038/nrmicro3367. PMC 5619696 . PMID 25417656.
^ Khaperskyy DA, Schmaling S, Larkins-Ford J, McCormick C, Gaglia MM (febrero de 2016). "Degradación selectiva de las transcripciones de la ARN polimerasa II del huésped mediante la proteína de apagado del huésped PA-X del virus de la influenza A". Más patógenos . 12 (2): e1005427. doi : 10.1371/journal.ppat.1005427 . PMC 4744033 . PMID 26849127.
^ Te Velthuis AJ, Fodor E (agosto de 2016). "ARN polimerasa del virus de la influenza: conocimientos sobre los mecanismos de síntesis de ARN viral". Reseñas de la naturaleza. Microbiología . 14 (8): 479–93. doi :10.1038/nrmicro.2016.87. PMC 4966622 . PMID 27396566.
^ ab Smith AE, Helenius A (abril de 2004). "Cómo entran los virus en las células animales". Ciencia . 304 (5668): 237–42. Código Bib : 2004 Ciencia... 304.. 237S. doi : 10.1126/ciencia.1094823. PMID 15073366. S2CID 43062708.
^ Yoshiyuki Suzuki, Masatoshi Nei (1 de abril de 2001). "Origen y evolución de los genes de hemaglutinina del virus de la influenza". Biología Molecular y Evolución . Académico de Oxford. 19 (4): 501–509. doi : 10.1093/oxfordjournals.molbev.a004105 . PMID 11919291.
^ Barry RD (agosto de 1961). "La multiplicación del virus de la influenza. II. Reactivación de la multiplicidad del virus irradiado con luz ultravioleta". Virología . 14 (4): 398–405. doi :10.1016/0042-6822(61)90330-0. hdl : 1885/109240 . PMID 13687359.
^ Henle W, Liu OC (octubre de 1951). "Estudios sobre las interacciones huésped-virus en el sistema embrión de pollo-virus de la influenza. VI. Evidencia de reactivación multiplicidad de virus inactivados". La Revista de Medicina Experimental . 94 (4): 305–22. doi :10.1084/jem.94.4.305. PMC 2136114 . PMID 14888814.
^ Gilker JC, Pavilanis V, Ghys R (junio de 1967). "Reactivación de multiplicidad en virus de influenza irradiados con gamma". Naturaleza . 214 (5094): 1235–7. Código bibliográfico : 1967Natur.214.1235G. doi :10.1038/2141235a0. PMID 6066111. S2CID 4200194.
^ Peterhans E (mayo de 1997). "Oxidantes y antioxidantes en enfermedades virales: mecanismos de enfermedad y regulación metabólica". La Revista de Nutrición . 127 (5 suplementos): 962S–965S. doi : 10.1093/jn/127.5.962S . PMID 9164274.
^ Bernstein H, Byerly HC, Hopf FA, Michod RE (octubre de 1984). "Origen del sexo". Revista de Biología Teórica . 110 (3): 323–51. Código Bib : 1984JThBi.110..323B. doi :10.1016/S0022-5193(84)80178-2. PMID 6209512.
^ Datos clave de los CDC sobre la influenza aviar (gripe aviar) y el virus de la influenza aviar A (H5N1)
^ Episcopio D, Aminov S, Benjamin S, Germain G, Datan E, Landazuri J, Lockshin RA, Zakeri Z (abril de 2019). "La atorvastatina restringe la capacidad del virus de la influenza para generar gotitas de lípidos y suprime gravemente la replicación del virus". La Revista FASEB . 33 (8): 9516–9525. doi : 10.1096/fj.201900428RR . PMC 6662987 . PMID 31125254.
^ de Jong MD, Bach VC, Phan TQ, Vo MH, Tran TT, Nguyen BH, Beld M, Le TP, Truong HK, Nguyen VV, Tran TH, Do QH, Farrar J (febrero de 2005). "Influenza aviar A (H5N1) mortal en un niño que presenta diarrea seguida de coma". El diario Nueva Inglaterra de medicina . 352 (7): 686–91. doi : 10.1056/NEJMoa044307 . PMID 15716562. S2CID 17703507.
^ "Nueva cepa de gripe aviar en China" una de las más letales "advierte la OMS". MercoPrensa . Consultado el 6 de diciembre de 2020 .
^ "La próxima pandemia y cómo evitarla: llevar una dieta basada en plantas". Poste matutino del sur de China . 19 de octubre de 2020 . Consultado el 6 de diciembre de 2020 .
^ "Dando el salto". Noticias . 677 Huntington Avenue, Boston, Ma 02115 +1495‑1000. 24 de octubre de 2013 . Consultado el 6 de diciembre de 2020 .
^ Ungchusak K, Auewarakul P, Dowell SF, Kitphati R, Auwanit W, Puthavathana P, Uiprasertkul M, Boonnak K, Pittayawonganon C, Cox NJ, Zaki SR (27 de enero de 2005). "Probable transmisión de persona a persona de la influenza aviar A (H5N1)". Revista de Medicina de Nueva Inglaterra . 352 (4): 333–340. doi : 10.1056/NEJMoa044021 . ISSN 0028-4793. PMID 15668219.
^ ab Komadina N, McVernon J, Hall R, Leder K (2014). "Una perspectiva histórica del virus de la influenza A (H1N2)". Enfermedades infecciosas emergentes . 20 (1): 6-12. doi :10.3201/eid2001.121848. PMC 3884707 . PMID 24377419.{{cite journal}}: Mantenimiento CS1: varios nombres: lista de autores ( enlace )
^ Mahmoud 2005, pag. 7
^ "NOVA | scienceNOW | Reviviendo el virus (no Flash) | PBS". www.pbs.org . Consultado el 6 de diciembre de 2020 .
^ "Virus de la influenza A (H1N2): descripción general". Temas de ScienceDirect . Consultado el 21 de febrero de 2021 .
^ Cuadro detallado de su evolución aquí Archivado el 9 de mayo de 2009 en Wayback Machine en PDF llamado Ecología y evolución de la gripe.
^ Mahmoud 2005, pag. 115 "Existe una presión especial para reconocer y prestar atención a las lecciones de pandemias de gripe pasadas a la sombra de la preocupante temporada de gripe de 2003-2004. Una forma grave y de aparición temprana de gripe A H3N2 ocupó los titulares cuando se cobró la vida de varios niños en Estados Unidos a finales de 2003. Como resultado, la demanda anual de vacuna inactivada contra la gripe, más fuerte de lo normal, superó el suministro de vacunas, de las cuales entre el 10 y el 20 por ciento normalmente no se utilizan. si la temporada 2003-2004 fuera testigo de un cambio significativo en los patrones de mortalidad".
^ Motivo Archivado el 26 de octubre de 2006 en Wayback Machine Altman LK (15 de enero de 2006). "El virus de la gripe de esta temporada es resistente a dos medicamentos estándar". Los New York Times .
^ Artículo de CBS News Decenas de personas en Japón pueden tener gripe aviar leve, enero de 2006.
^ Ogata T, Yamazaki Y, Okabe N, Nakamura Y, Tashiro M, Nagata N, Itamura S, Yasui Y, Nakashima K, Doi M, Izumi Y, Fujieda T, Yamato S, Kawada Y (julio de 2008). "Infección por influenza aviar humana H5N2 en Japón y los factores asociados con un alto título de anticuerpos neutralizantes del H5N2". Revista de Epidemiología . 18 (4): 160–6. doi :10.2188/jea.JE2007446. PMC 4771585 . PMID 18603824. Archivado desde el original el 28 de septiembre de 2018 . Consultado el 14 de marzo de 2009 .
^ "Rusia informa los primeros casos humanos de gripe aviar H5N8". Noticias BNO . 20 de febrero de 2021 . Consultado el 20 de febrero de 2021 .
^ "Rusia registra los primeros casos de infección humana con la cepa H5N8 de la gripe aviar". Noticias del cielo . 20 de febrero de 2021 . Consultado el 21 de febrero de 2021 .
^ QUIÉN
^ ab CDC Infección por influenza aviar en humanos
^ Tweed SA, Skowronski DM, David ST, Larder A, Petric M, Lees W, Li Y, Katz J, Krajden M, Tellier R, Halpert C, Hirst M, Astell C, Lawrence D, Mak A (diciembre de 2004). "Enfermedad humana por influenza aviar H7N3, Columbia Británica". Enfermedades infecciosas emergentes . 10 (12): 2196–9. doi : 10.3201/eid1012.040961. PMC 3323407 . PMID 15663860.
^ Schnirring L (2 de abril de 2013). "China informa 4 infecciones más por H7N9". Noticias CIDRAP . Archivado desde el original el 17 de mayo de 2013 . Consultado el 10 de abril de 2013 .
^ "Virus de la influenza aviar A (H7N9) | Influenza aviar (gripe)". www.cdc.gov . Consultado el 24 de febrero de 2017 .
^ niaid.nih.gov Archivado el 26 de diciembre de 2005 en Wayback Machine Cronología de las pandemias de gripe humana
^ "China informa del primer caso humano de gripe aviar H10N3". Reuters . 1 de junio de 2021 . Consultado el 22 de junio de 2021 .
^ Taubenberger JK, Morens DM (enero de 2006). "Gripe de 1918: la madre de todas las pandemias". Enfermedades infecciosas emergentes . 12 (1): 15-22. doi : 10.3201/eid1201.050979. PMC 3291398 . PMID 16494711.
^ Artículo del Science Daily Un nuevo estudio tiene implicaciones importantes para la vigilancia de la gripe publicado el 27 de octubre de 2006
^ Nelson MI, Simonsen L, Viboud C, Miller MA, Taylor J, George KS, Griesemer SB, Ghedin E, Ghedi E, Sengamalay NA, Spiro DJ, Volkov I, Grenfell BT, Lipman DJ, Taubenberger JK, Holmes EC (diciembre 2006). "Los procesos estocásticos son determinantes clave de la evolución a corto plazo del virus de la influenza a". Más patógenos . 2 (12): e125. doi : 10.1371/journal.ppat.0020125 . PMC 1665651 . PMID 17140286.
^ Smith DJ, Lapedes AS, de Jong JC, Bestebroer TM, Rimmelzwaan GF, Osterhaus AD, Fouchier RA (julio de 2004). "Mapeo de la evolución antigénica y genética del virus de la influenza". Ciencia . 305 (5682): 371–6. Código Bib : 2004 Ciencia... 305.. 371S. doi : 10.1126/ciencia.1097211 . PMID 15218094. S2CID 1258353.
^ Mahmoud 2005, pag. 30
^ Mahmoud 2005, pag. 82 "Curiosamente, los virus de la influenza recombinantes que contienen 1918 HA y NA y hasta tres genes adicionales derivados del virus 1918 (los otros genes se derivan del virus A/WSN/33) fueron todos altamente virulentos en ratones (Tumpey et al. , 2004). Además, el análisis de microarrays de expresión realizado en tejido pulmonar completo de ratones infectados con el recombinante 1918 HA/NA mostró una mayor regulación positiva de los genes implicados en la apoptosis, la lesión tisular y el daño oxidativo (Kash et al., 2004). eran inusuales porque los virus con los genes 1918 no se habían adaptado a los ratones. La finalización de la secuencia del genoma completo del virus 1918 y la reconstrucción y caracterización de los virus con los genes 1918 en condiciones de bioseguridad apropiadas arrojarán más luz sobre estos hallazgos. y debería permitir un examen definitivo de esta explicación. El análisis antigénico de virus recombinantes que poseen HA y NA de 1918 mediante pruebas de inhibición de la hemaglutinación utilizando antisueros de hurón y pollo sugirieron una estrecha relación con el virus A/porcino/Iowa/30 y los virus H1N1 aislados en el década de 1930 (Tumpey et al., 2004), lo que respalda datos adicionales de Shope de la década de 1930 (Shope, 1936). Curiosamente, cuando se inmunizaron ratones con diferentes cepas del virus H1N1, los estudios de exposición utilizando virus similares a 1918 revelaron una protección parcial con este tratamiento, lo que sugiere que la actual [ ¿cuándo? ] Las estrategias de vacunación son adecuadas contra un virus similar al de 1918 (Tumpey et al., 2004)".
^ "Influenza aviar A (H5N1) - actualización 31: Situación (aves de corral) en Asia: necesidad de una respuesta a largo plazo, comparación con brotes anteriores". Alerta y Respuesta ante Epidemias y Pandemias (EPR) . OMS. 2004. Archivado desde el original el 7 de marzo de 2004. Brotes conocidos de gripe altamente patógena en aves de corral 1959-2003.
^ Geraci JR, St Aubin DJ, Barker IK, Webster RG, Hinshaw VS, Bean WJ, Ruhnke HL, Prescott JH, Early G, Baker AS, Madoff S, Schooley RT (febrero de 1982). "Mortalidad masiva de focas comunes: neumonía asociada al virus de la influenza A". Ciencia . 215 (4536): 1129–31. Código Bib : 1982 Ciencia... 215.1129G. doi : 10.1126/ciencia.7063847. PMID 7063847. Más de 400 focas comunes, la mayoría de ellas inmaduras, murieron a lo largo de la costa de Nueva Inglaterra entre diciembre de 1979 y octubre de 1980 de neumonía aguda asociada con el virus de la influenza A/Seal/Mass/1/180 (H7N7). El virus tiene características aviares, se replica principalmente en mamíferos y causa enfermedades respiratorias leves en focas infectadas experimentalmente. La infección concurrente con un micoplasma no descrito previamente o condiciones ambientales adversas pueden haber desencadenado la epizootia. Las similitudes entre esta epizootia y otras mortalidades de focas en el pasado sugieren que estos eventos pueden estar relacionados por factores biológicos y ambientales comunes.
^ ab Centros CDC para el Control y la Prevención de Enfermedades - Transmisión de los virus de la influenza A entre animales y personas
^ Kandeil A, Gomaa MR, Shehata MM, El Taweel AN, Mahmoud SH, Bagato O (enero de 2019). "Aislamiento y caracterización de un virus de la influenza A distinto de murciélagos egipcios". Revista de Virología . 93 (2): e01059-18. doi :10.1128/JVI.01059-18. PMC 6321940 . PMID 30381492.
^ Ciminski K, Ran W, Gorka M, Lee J, Schinköthe J, Eckley M, Murrieta MA, Aboellail TA, Campbell CL, Ebel GD, Ma J, Pohlmann A, Franzke K, Ulrich R, Hoffmann D, García-Sastre A , Ma W, Schountz T, Beer M, Schwemmle M (2019). "Los virus de la influenza en murciélagos se transmiten entre murciélagos, pero están mal adaptados a especies que no son murciélagos". Microbiología de la naturaleza . 4 (12): 2298–2309. doi :10.1038/s41564-019-0556-9. PMC 7758811 . PMID 31527796. S2CID 202580293.
Sitio web de información sobre la influenza aviar del gobierno de EE. UU.
Centro Europeo para la Prevención y el Control de Enfermedades ( ECDC ) Estocolmo, Suecia
información general
"La gripe aviar y usted" Póster a todo color proporcionado por el Centro de Tecnología y Política de Seguridad Nacional de la Universidad de Defensa Nacional , en colaboración con el Centro de Políticas de Salud de Seguridad Nacional
Beigel JH, Farrar J, Han AM, Hayden FG, Hyer R, de Jong MD, Lochindarat S, Nguyen TK, Nguyen TH, Tran TH, Nicoll A, Touch S, Yuen KY (septiembre de 2005). "Infección por influenza aviar A (H5N1) en humanos". El diario Nueva Inglaterra de medicina . 353 (13): 1374–85. CiteSeerX 10.1.1.730.7890 . doi :10.1056/NEJMra052211. PMID 16192482.
Influenza pandémica: Esfuerzos de preparación nacional Informe del Servicio de Investigación del Congreso sobre preparación para una pandemia.
Una guía sobre la gripe aviar y sus síntomas de BBC Health
Mahmud (2005). Stacey L. Knobler, Alison Mack, Mahmoud A, Stanley M. Lemon (eds.). La amenaza de una gripe pandémica: ¿estamos preparados? : resumen del taller / preparado para el Foro sobre Amenazas Microbianas, Junta de Salud Global . Prensa de las Academias Nacionales. pag. 285.ISBN 0-309-09504-2. El virus de la influenza aviar altamente patógeno está en cada lista de los diez principales agentes disponibles como posibles armas biológicas agrícolas.
Mahmoud AA, Instituto de Medicina, Knobler S, Mack A (2005). La amenaza de la gripe pandémica: ¿Estamos preparados?: Resumen del taller. Washington, DC: Prensa de Academias Nacionales. ISBN 978-0-309-09504-4.
Enlaces a fotografías de la gripe aviar (Hardin MD/Univ de Iowa)
Kawaoka Y (2006). Virología de la influenza: temas actuales . Pr. Académico Caister. ISBN 978-1-904455-06-6.
Sobrino F, Mettenleiter T (2008). Virus animales: biología molecular . Pr. Académico Caister. ISBN 978-1-904455-22-6.
enlaces externos
Base de datos de investigación de influenza: base de datos de secuencias genómicas de influenza e información relacionada.





.jpg/1280px-Genetic_Relationships_Among_Human_and_Swine_Influenza_Viruses,_1918-2009_(7704014350).jpg)