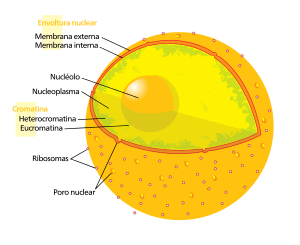
Núcleo celular
La función del núcleo es mantener la integridad de esos genes y controlar las actividades celulares regulando la expresión génica.
No obstante, Hertwig confirmó su observación en otros grupos animales, como por ejemplo en anfibios y moluscos.
[14] Las hormonas esteroideas como el cortisol y la aldosterona, así como otras moléculas pequeñas hidrosolubles implicadas en la señalización celular, pueden difundir a través de la membrana celular y en el citoplasma, donde se unen a proteínas que actúan como receptores nucleares que son conducidas al núcleo.
[7] En las células animales existen dos redes de filamentos intermedios que proporcionan soporte mecánico al núcleo: la lámina nuclear forma una trama organizada en la cara interna de la envoltura, mientras que en la cara externa este soporte es menos organizado.
[25] Se han asociado anticuerpos a ciertos tipos de organización cromatínica, en particular los nucleosomas con varias enfermedades autoinmunes como el lupus eritematoso sistémico.
[26] Estos son conocidos como anticuerpos antinucleares (ANA) y también se han observado en concierto con la esclerosis múltiple en el contexto de una disfunción inmune generalizada.
Estas subunidades ribosomales ensambladas son las estructuras más grandes que pasan a través de los poros nucleares.
[29] Además del nucléolo, el núcleo contiene una cierta cantidad de cuerpos delimitados no membranosos.
Esta enfermedad se produce típicamente por mutaciones en el gen de la actina, y los bastones en sí mismos están constituidos por la actina producida a partir de tales genes mutantes, así como otras proteínas del citoesqueleto.
[40] Los paraspeckles son estructuras dinámicas que se alteran en respuesta a cambios en la actividad celular metabólica.
A diferencia de otros cuerpos nucleares, aparecen solamente durante determinados periodos del ciclo celular.
Este mecanismo regulador tiene lugar en el caso de la glucólisis, una ruta celular en la que se utiliza la glucosa para producir energía.
[47] El ARNm contiene intrones que se deben retirar antes de ser traducidos para producir proteínas funcionales.
La expresión génica implica en primer lugar la transcripción, en la que el ADN se utiliza como molde para producir ARN.
Puesto que los ribosomas se localizan fuera del núcleo, el ARNm sintetizado debe ser exportado.
Posteriormente se deben someter a modificación post-transcripcional en el núcleo antes de ser exportados al citoplasma.
La capacidad de las importinas y las exportinas para transportar su carga está regulada por GTPasas, enzimas que hidrolizan GTP liberando energía.
Estas señales únicamente están presentes en las proteínas nucleares, consisten en una secuencia corta que va entre 4 y 8 aminoácidos.
Su clasificación depende de la dirección del transporte para el que fueron inicialmente descritas, se les ha clasificado como importinas y exportinas.
Durante estos acontecimientos, los componentes estructurales del núcleo —la envoltura y la lámina— son sistemáticamente degradados.
[51] Por tanto, en los estadios tempranos del ciclo celular, comenzando en profase y hasta casi la prometafase, se desmantela la membrana nuclear.
Los eritrocitos maduran gracias a la eritropoyesis en la médula ósea, donde pierden su núcleo, orgánulos y ribosomas.
[59] En los seres humanos, el músculo esquelético posee células, llamadas miocitos, que se convierten en polinucleadas durante su desarrollo.
[61] Al ser la mejor característica que define la célula eucariota, el origen evolutivo del núcleo ha sido objeto de mucha especulación.
Entre las teorías propuestas, se pueden considerar cuatro como las principales, aunque ninguna de ellas ha encontrado un amplio apoyo.
[62] La teoría conocida como "modelo sintrófico" propone que una relación simbiótica entre arqueas y bacterias creó la primera célula eucariota nucleada.
Esta teoría es análoga a teoría aceptada del origen de las mitocondrias y cloroplastos eucariotas, de los que se piensa que se han desarrollado por una relación endosimbionte similar entre protoeucariotas y bacterias aerobias.
[63] El origen arqueano del núcleo está apoyado por la circunstancia de que tanto arqueas como eucariotas tienen genes similares en ciertas proteínas, incluyendo las histonas.
Una versión de esta propuesta sugiere que el núcleo evolucionó concertadamente con la fagocitosis para dar lugar a un depredador celular primitivo.
[70] Finalmente, una propuesta muy reciente sugiere que las variantes tradicionales de las teorías endosimbiontes son insuficientes para explicar el origen del núcleo eucariota.