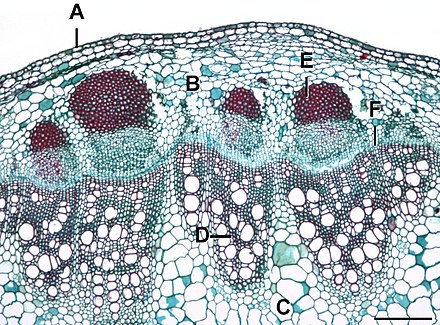
Cámbium
La segunda, hacia afuera, es otro tipo de tejido el floema, que transporta savia elaborada en dirección a las raíces.
Durante el crecimiento secundario, las células de los rayos medulares, en una línea (como se ve en sección; en tres dimensiones, es una lámina) entre haces vasculares vecinos, se convierten en meristemos y forman un nuevo cambium interfascicular (entre haces vasculares).
Actualmente, tanto las hormonas como los péptidos cortos han sido identificados como portadores de información en estos sistemas.
Las señales recibidas desde el exterior del meristemo actúan para regular a la baja los factores internos, lo que promueve la proliferación y diferenciación celular.
Se ha demostrado que los mutantes sin auxina mostrarán un mayor espacio entre los cámbium interfasciculares y un menor crecimiento de los haces vasculares.
Los niveles de etileno son elevados en las plantas con una zona cambial activa y todavía se están estudiando.
El cambium se crea a partir de las células del parénquima interfascicular[9] que sufren desdiferenciación.
En algunas plantas, especialmente en las dicotiledóneas leñosas, se forma un anillo cambial tan pronto como cesa el crecimiento en longitud, a veces después de unos diez días, comienza la estructura secundaria, el felógeno se iniciará más tarde.
[14] El líber de los siguientes árboles es comestible y se ha consumido:[14] sauce (Salix alba), haya (Fagus sylvatica, Fagus orientalis), arce (todos), abedul (Betula pendula [15]), tilleul (todos), olmo (Ulmus glabra, Ulmus minor/campestris, Ulmus fulva), Álamo (Populus alba, Populus nigra, Populus tremula), Roble de Pensilvania,[16] y Pinaceae (todos).
[19] Tampoco sabemos si el cambium tiene biosensores y mecanismos intrínsecos para regular la producción de madera, o si esta regulación es el resultado de fitohormonas u otras señales aún por explorar.
[20] En 2015, la tesis de Félix Hartmann propuso un modelo biofísico del cambium y su funcionamiento.