Si bien un estudio genético indica que R1a se originó hace 25.000 [2] años, su subclado M417 (R1a1a1) se diversificó c. Hace 5.800 años. [4] El lugar de origen del subclado juega un papel en el debate sobre los orígenes de los protoindoeuropeos .
La mutación del SNP R-M420 se descubrió después de R-M17 (R1a1a), lo que resultó en una reorganización del linaje, en particular estableciendo un nuevo paragrupo (designado R-M420*) para los linajes relativamente raros que no están en el R-SRY10831. .2 (R1a1) ramal que conduce a la R-M17.
Orígenes
Orígenes de R1a
Se estima que la divergencia genética de R1a (M420) ocurrió hace 25.000 [2] años, que es el momento del último máximo glacial . Un estudio de 2014 realizado por Peter A. Underhill et al., utilizando 16.244 individuos de más de 126 poblaciones de toda Eurasia, concluyó que había "un caso convincente para el Medio Oriente, posiblemente cerca del actual Irán, como origen geográfico del hg R1a". ". [2] El antiguo registro de ADN ha mostrado el primer R1a durante el Mesolítico en los cazadores-recolectores del este (de Europa del Este, hace c. 13.000 años), [5] [6] y el caso más antiguo de R* entre el Norte antiguo del Paleolítico superior. Euroasiáticos , [7] de los cuales los cazadores-recolectores orientales derivan predominantemente su ascendencia. [8]
Diversificación de R1a1a1 (M417) y migraciones antiguas.
Orígenes de R1a (Underhill 2009; [3] orígenes de R1a1a (Pamjav et al. 2012); posible migración de R1a a la costa báltica; y expansión más antigua y de mayor frecuencia de R1a1a (Underhill et al. 2014)
Según Underhill et al. (2014), el subclado R1a-M417 aguas abajo se diversificó en Z282 y Z93 hace alrededor de 5.800 años "en las cercanías de Irán y el este de Turquía". [4] [nota 1] Aunque R1a ocurre como un haplogrupo del cromosoma Y entre varias lenguas como el eslavo y el indoiraní , la cuestión de los orígenes de R1a1a es relevante para el debate en curso sobre el urheimat del protoindo-iraní. Pueblo europeo , y también puede ser relevante para los orígenes de la civilización del valle del Indo . R1a muestra una fuerte correlación con las lenguas indoeuropeas del sur y oeste de Asia , Europa central y oriental y, en cierta medida , Escandinavia [10] [3] , siendo más prevalente en Europa del este , Asia occidental y Asia del sur . En Europa, Z282 prevalece especialmente, mientras que en Asia domina Z93. La conexión entre el ADN-Y R-M17 y la difusión de las lenguas indoeuropeas fue notada por primera vez por T. Zerjal y sus colegas en 1999. [11]
Dispersión esteparia propuesta de R1a1a
Semino et al. (2000) propusieron orígenes ucranianos y una expansión posglacial del haplogrupo R1a1 durante el Máximo Glacial Tardío , posteriormente magnificada por la expansión de la cultura Kurgan hacia Europa y hacia el este. [12] Spencer Wells propone orígenes en Asia Central, sugiriendo que la distribución y edad de R1a1 apunta a una antigua migración correspondiente a la expansión del pueblo Kurgan en su expansión desde la estepa euroasiática . [13] Según Pamjav et al. (2012), R1a1a se diversificó en las estepas euroasiáticas o en la región de Medio Oriente y el Cáucaso:
Asia interior y central es una zona de superposición para los linajes R1a1-Z280 y R1a1-Z93 [lo que] implica que una zona de diferenciación temprana de R1a1-M198 posiblemente ocurrió en algún lugar dentro de las estepas euroasiáticas o en la región de Medio Oriente y el Cáucaso, ya que se encuentran entre el Sur y el Sur. Asia y Europa central y oriental. [14]
Tres estudios genéticos realizados en 2015 respaldaron la teoría de Kurgan de Gimbutas con respecto al Urheimat indoeuropeo . Según esos estudios, los haplogrupos R1b y R1a, ahora los más comunes en Europa (R1a también es común en el sur de Asia) se habrían expandido desde las estepas del Póntico-Caspio, junto con las lenguas indoeuropeas; también detectaron un componente autosómico presente en los europeos modernos que no estaba presente en los europeos del Neolítico, que habría sido introducido con los linajes paternos R1b y R1a, así como con las lenguas indoeuropeas. [15] [16] [17]
Silva et al. (2017) señalaron que R1a en el sur de Asia "probablemente se propagó a partir de un único grupo de fuentes de Asia central ; parece haber al menos tres y probablemente más clados fundadores de R1a dentro del subcontinente indio , lo que es consistente con múltiples oleadas de llegada". [18] Según Martin P. Richards, coautor de Silva et al. (2017), la prevalencia de R1a en la India fue "una prueba muy poderosa de una migración sustancial de la Edad del Bronce desde Asia central que probablemente trajo hablantes indoeuropeos a la India". [19] [nota 2]
Orígenes propuestos del sur de Asia
Kivisild et al. (2003) han propuesto Asia meridional u occidental , [20] [nota 3] mientras que Mirabal et al. (2009) ven apoyo tanto para Asia meridional como central. [10] Sengupta y cols. (2006) han propuesto orígenes indios. [21] Thanseem y otros. (2006) han propuesto Asia meridional o central. [22] Sahoo et al. (2006) han propuesto Asia meridional u occidental. [23] Thangaraj et al. (2010) también han propuesto un origen del sur de Asia. [24] Sharma et al. (2009) teorizan la existencia de R1a en la India más allá de 18.000 años a posiblemente 44.000 años en su origen. [1]
Las poblaciones del sur de Asia tienen la mayor diversidad de STR dentro de R1a1a, [25] [26] [10] [3] [1] [27] y dataciones TMRCA más antiguas posteriores , [ cita necesaria ] y R1a1a está presente entre las dos castas superiores ( brahmanes ) y castas inferiores, aunque la frecuencia es mayor entre las castas brahmanes. Sin embargo, las dataciones TMRCA más antiguas del haplogrupo R1a se producen en la tribu Saharia , una casta registrada de la región de Bundelkhand en la India central . [1] [27] A partir de estos hallazgos, algunos investigadores han concluido que R1a1a se originó en el sur de Asia, [26] [1] [nota 4] [nota 5] excluyendo una afluencia genética más reciente, aunque menor, de inmigrantes indoeuropeos en regiones del noroeste como Afganistán, Baluchistán, Punjab y Cachemira. [26] [25] [3]
Sin embargo, esta diversidad, y las posteriores dataciones TMRCA más antiguas, también pueden explicarse por las cifras de población históricamente altas, [nota 6] , lo que aumenta la probabilidad de diversificación y variación de microsatélites . [19] [18] Según Sengupta et al. (2006), "[R1a1 y R2] podrían haber llegado varias veces al sur de la India desde una región de origen del suroeste de Asia". [25] [nota 7] Sin embargo, Sengupta también describió en este artículo:
Descubrimos que la influencia de Asia Central en el acervo genético preexistente era menor. Las edades de variación acumulada de microsatélites en la mayoría de los haplogrupos indios superan los 10.000 a 15.000 años, lo que atestigua la antigüedad de la diferenciación regional. Por lo tanto, nuestros datos no respaldan modelos que invoquen un aporte genético reciente pronunciado de Asia Central para explicar la variación genética observada en el sur de Asia.
En la publicación del MIT, "La desigualdad, una teoría genética", se afirma que "la reconstrucción filogenética de R1a no respalda un origen del sur de Asia". A pesar de que la fuente (es decir, el MIT) tiene credibilidad, la reconstrucción filogenética a la que se refiere el autor no se cita, excluyendo también el hecho de que las reconstrucciones filogenéticas tienen grandes márgenes de error que se atribuyen principalmente a la "incertidumbre del modelo", ya que se hacen suposiciones sobre la evolución/mutación. proceso de antemano (Aquí se referiría a Diferentes modelos de evolución pueden producir diferentes árboles filogenéticos, lo que lleva a incertidumbre en las relaciones inferidas), excluyendo también los errores generados debido al muestreo, la medición y, por supuesto, la incertidumbre estadística. La posición predominante entre los genetistas es que el haplogrupo R1a no se originó en el sur de Asia, y que los portadores del haplogrupo R1a se trasladaron del oeste de Asia al sur de Asia algún tiempo después del establecimiento de la civilización del valle del Indo . [29] [30]
Orígenes propuestos de Yamnaya
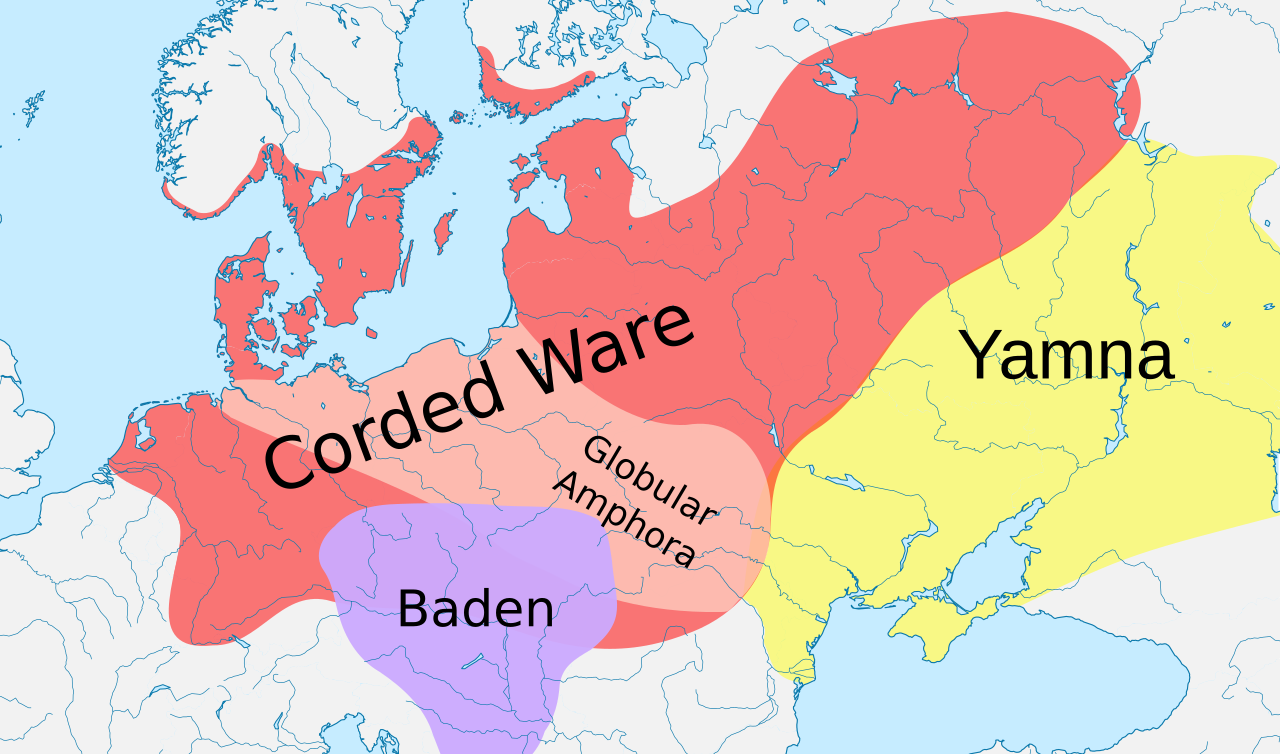
Neolítico medio europeo. Cultura Comb Ware c. 4200 – c. 2000 a. C.Cultura Corded Ware (c. 2900 - c. 2350 a. C.)
David Anthony considera que la cultura Yamnaya es el Urheimat indoeuropeo . [31] [32] Según Haak et al. (2015), se produjo una migración masiva de la cultura Yamnaya hacia el norte c. 2500 a. C., lo que representa el 75% de la ascendencia genética de la cultura Corded Ware , señalando que R1a y R1b pueden haberse "extendido a Europa desde el Este después del 3000 a. C.". [33] Sin embargo, las siete muestras de Yamnaya pertenecían al subclado R1b-M269 , [33] pero no se ha encontrado ningún R1a1a en sus muestras de Yamnaya. Esto plantea la pregunta de dónde vino el R1a1a en la cultura Corded Ware, si no fue de la cultura Yamnaya. [34]
Según Marc Haber, la ausencia del haplogrupo R1a-M458 en Afganistán no respalda un origen estepario del Póntico-Caspio para los linajes R1a en las poblaciones modernas de Asia Central. [35]
Según Leo Klejn , la ausencia del haplogrupo R1a en los restos de Yamnaya (a pesar de su presencia en las poblaciones eneolíticas de Samara y de cazadores-recolectores del este ) hace que sea poco probable que los europeos heredaran el haplogrupo R1a de Yamnaya. [36]
El arqueólogo Barry Cunliffe ha dicho que la ausencia del haplogrupo R1a en especímenes de Yamnaya es una debilidad importante en la propuesta de Haak de que R1a tiene un origen Yamnaya. [37]
Semenov y Bulat (2016) defienden un origen Yamnaya de R1a1a en la cultura Corded Ware, y señalan que varias publicaciones apuntan a la presencia de R1a1 en la cultura Comb Ware . [38] [nota 8]
Orígenes propuestos de Transcaucasia y Asia occidental y posible influencia en la civilización del valle del Indo
Haak et al. (2015) encontraron que parte de la ascendencia Yamnaya derivaba del Medio Oriente y que las técnicas neolíticas probablemente llegaron a la cultura Yamnaya desde los Balcanes . [nota 9] La cultura Rössen (4.600-4.300 a. C.), que estaba situada en Alemania y es anterior a la cultura Corded Ware, todavía se puede encontrar un antiguo subclado de R1a, a saber, L664. [nota 10]
Parte de la ascendencia genética del sur de Asia deriva de poblaciones de Eurasia occidental, y algunos investigadores han insinuado que Z93 pudo haber llegado a la India a través de Irán [40] y expandirse allí durante la civilización del valle del Indo . [2] [41]
Mascarenhas et al. (2015) propusieron que las raíces de Z93 se encuentran en Asia occidental, y propusieron que "Z93 y L342.2 se expandieron en dirección sureste desde Transcaucasia hacia el sur de Asia ", [40] señalando que tal expansión es compatible con "los registros arqueológicos de la expansión hacia el este de las poblaciones de Asia occidental en el cuarto milenio a. C. que culminó con las llamadas migraciones Kura-Araxes en el período posterior a Uruk IV . [40] Sin embargo, Lazaridis señaló que la muestra I1635 de Lazaridis et al. (2016), su muestra armenia de Kura-Araxes, portaba el haplogrupo Y R1 b 1-M415(xM269) [nota 11] (también llamado R1b1a1b-CTS3187). [42] [ ¿ fuente poco confiable? ]
Según Underhill et al. (2014) la diversificación de Z93 y la "urbanización temprana dentro del Valle del Indo... ocurrió [hace 5.600 años] y la distribución geográfica de R1a-M780 (Figura 3d [nota 12] ) puede reflejar esto". [2] [nota 13] Poznik et al. (2016) señalan que se produjeron "expansiones sorprendentes" dentro de R1a-Z93 en c. Hace 4.500 a 4.000 años, que "es anterior en unos pocos siglos al colapso de la civilización del valle del Indo". [41] [nota 14]
Sin embargo, según Narasimhan et al. (2018), los pastores esteparios son una fuente probable de R1a en la India. [44] [nota 15]
Filogenia
El árbol genealógico R1a ahora tiene tres niveles principales de ramificación, con el mayor número de subclados definidos dentro de la rama dominante y más conocida, R1a1a (que se encontrará con varios nombres como "R1a1" en literatura relativamente reciente, pero no en la más reciente). .
Topología
La topología de R1a es la siguiente (códigos [entre paréntesis] códigos que no son isogg): [9] [45] [ se necesita verificación ] [46] [2] [47] Tatiana et al. (2014) "El rápido proceso de diversificación de K-M526 probablemente ocurrió en el sudeste asiático , con posteriores expansiones hacia el oeste de los ancestros de los haplogrupos R y Q ". [48]
R1a se distingue por varios marcadores únicos, incluida la mutación M420. Es un subclado del Haplogrupo R-M173 (anteriormente llamado R1). R1a tiene los subclados hermanos Haplogrupo R1b -M343 y el paragrupo R-M173*.
R-M420 (R1a)
R-M420, definida por la mutación M420, tiene dos ramas: R-SRY1532.2, definida por la mutación SRY1532.2, que constituye la gran mayoría; y R-M420*, el paragrupo , definido como M420 positivo pero SRY1532.2 negativo. (En el esquema de 2002, esta minoría negativa SRY1532.2 era una parte del grupo relativamente raro clasificado como paragrupo R1*). Las mutaciones que se entienden equivalentes a M420 incluyen M449, M511, M513, L62 y L63. [3] [51]
Este paragrupo ahora se conoce como R1a2 (R-YP4141). Luego tiene dos ramas R1a2a (R-YP5018) y R1a2b (R-YP4132).
R-SRY1532.2 (R1a1)
R1a1 está definido por SRY1532.2 o SRY10831.2 (se entiende que siempre incluye SRY10831.2, M448, L122, M459 y M516 [3] [52] ). Esta familia de linajes está dominada por M17 y M198. Por el contrario, el paragrupo R-SRY1532.2* carece de los marcadores M17 o M198.
El paragrupo R-SRY1532.2* es aparentemente menos raro que el R1*, pero sigue siendo relativamente inusual, aunque se ha probado en más de una encuesta. Underhill et al. (2009) informaron 1/51 en Noruega , 3/305 en Suecia , 1/57 macedonios griegos , 1/150 iraníes, 2/734 armenios étnicos y 1/141 kabardianos . [3] Sahoo et al. (2006) informaron R-SRY1532.2* para 1/15 muestras de Himachal Pradesh Rajput. [26]
R-M17/M198 (R1a1a)
Los siguientes SNP están asociados con R1a1a:
R-M417 (R1a1a1)
R1a1a1 (R-M417) es el subclado más ampliamente encontrado, en dos variaciones que se encuentran respectivamente en Europa (R1a1a1b1 (R-Z282) ([R1a1a1a*] (R-Z282) (Underhill 2014) [2] ) y Central y Sur de Asia (R1a1a1b2 (R-Z93) ([R1a1a2*] (R-Z93) Underhill 2014) [2] ).
R-Z282 (R1a1a1b1a) (Europa del Este)
Este gran subclado parece abarcar la mayor parte de los R1a1a que se encuentran en Europa. [14]
R1a1a1b1a [R1a1a1a* (Underhill (2014))] (R-Z282*) ocurre en el norte de Ucrania, Bielorrusia y Rusia con una frecuencia de c. 20%. [2]
R1a1a1b1a3 [R1a1a1a1 (Underhill (2014))] (R-Z284) ocurre en el noroeste de Europa y alcanza su punto máximo en c. 20% en Noruega. [2]
R1a1a1c (M64.2, M87, M204) es aparentemente raro: se encontró en 1 de 117 machos tipificados en el sur de Irán. [53]
R-M458 (R1a1a1b1a1)
Distribución de frecuencia del R-M458
R-M458 es un SNP principalmente eslavo , caracterizado por su propia mutación, y fue llamado por primera vez grupo N. Underhill et al. (2009) descubrieron que estaba presente en las poblaciones europeas modernas aproximadamente entre la cuenca del Rin y los Montes Urales y lo rastrearon hasta "un efecto fundador que... cae en el período del Holoceno temprano, 7,9 ± 2,6 KYA". (Velocidades de Zhivotovsky, 3 veces sobrevaluadas) [3] M458 fue encontrado en un esqueleto de una tumba del siglo XIV en Usedom , Mecklemburgo-Pomerania Occidental, Alemania. [54] El artículo de Underhill et al. (2009) también informa una frecuencia sorprendentemente alta de M458 en algunas poblaciones del norte del Cáucaso (18% entre Ak Nogai , [55] 7,8% entre Qara Nogai y 3,4% entre Abazas ). [56]
R-L260 (R1a1a1b1a1a)
R1a1a1b1a1a (R-L260), comúnmente conocido como eslavo occidental o polaco , es un subclado del grupo parental más grande R-M458, y fue identificado por primera vez como un grupo STR por Pawlowski et al. 2002. En 2010 se verificó que era un haplogrupo identificado por su propia mutación (SNP). [57] Aparentemente representa alrededor del 8% de los hombres polacos, lo que lo convierte en el subclado más común en Polonia. Fuera de Polonia es menos común. [58] Además de Polonia, se encuentra principalmente en la República Checa y Eslovaquia , y se considera "claramente eslavo occidental". Se estima que el antepasado fundador de R-L260 vivió hace entre 2.000 y 3.000 años, es decir, durante la Edad del Hierro , con una importante expansión demográfica hace menos de 1.500 años. [59]
R-M334
R-M334 ([R1a1a1g1], [47] un subclado de [R1a1a1g] (M458) [47] cq R1a1a1b1a1 (M458) [46] ) fue encontrado por Underhill et al. (2009) solo en un hombre estonio y puede definir un clado pequeño y de fundación muy reciente. [3]
R1a1a1b1a2 (S466/Z280, S204/Z91)
R1a1a1b1a2b3* (Grupo K de Gwozdz)
R1a1a1b1a2b3* (M417+, Z645+, Z283+, Z282+, Z280+, CTS1211+, CTS3402, Y33+, CTS3318+, Y2613+) (Clúster K de Gwozdz) [ 45] [ se necesita verificación ] es un grupo basado en STR que es R-M17(xM458). Este grupo es común en Polonia pero no exclusivo de Polonia. [59]
R1a1a1b1a2b3a (R-L365)
R1a1a1b1a2b3a (R-L365) [46] se llamó inicialmente Grupo G. [ cita necesaria ]
R-Z93* o R1a1a1b2* (R1a1a2* en Underhill (2014)) es más común (>30%) en la región rusa de Altai en el sur de Siberia, apareciendo en Kirguistán (6%) y en todas las poblaciones iraníes (1-8 %). [2]
R-Z2125 ocurre con mayor frecuencia en Kirguistán y en los pastunes afganos (>40%). Con una frecuencia >10%, también se observa en otros grupos étnicos afganos y en algunas poblaciones del Cáucaso e Irán. [2]
R-M434 es un subclado de Z2125. Se detectó en 14 personas (de 3667 personas examinadas), todas en un rango geográfico restringido desde Pakistán hasta Omán . Esto probablemente refleja un evento de mutación reciente en Pakistán. [3]
R-M560 es muy raro y solo se observó en cuatro muestras: dos hablantes de burushaski (norte de Pakistán), un hazara (Afganistán) y un azerbaiyano iraní. [2]
R-M780 ocurre con alta frecuencia en el sur de Asia: India, Pakistán, Afganistán y el Himalaya. Turquía comparte sublinajes R1a (12,1%). [60] Los romaníes de Eslovaquia comparten el 3% de R1a. [61] El grupo también se encuentra en >3% en algunas poblaciones iraníes y está presente en >30% en los romaníes de Croacia y Hungría. [2]
Distribución geográfica de R1a1a
Distribución de R1a (morado) y R1b (rojo)
Prehistórico
En la Europa mesolítica, R1a es característico de los cazadores-recolectores del este (EHG). [62] Un EHG masculino de la cultura Veretye enterrado en Peschanitsa cerca del lago Lacha en el Óblast de Arkhangelsk , Rusia c. 10.700 a. C. se descubrió que era portador del haplogrupo paterno R1a5-YP1301 y del haplogrupo materno U4a . [63] [64] [62] Un hombre mesolítico de Karelia c. Se ha descubierto que entre el 8.800 a. C. y el 7.950 a. C. porta el haplogrupo R1a. [65] Un hombre mesolítico enterrado en Deriivka c. 7000 a. C. a 6700 a. C. portaban el haplogrupo paterno R1a y el materno U5a2a . [17] Otro macho de Karelia de c. 5.500 a 5.000 aC, que era considerado un EHG, portaba el haplogrupo R1a. [15] Un macho de la cultura Comb Ceramic en Kudruküla c. Se ha determinado que entre el 5.900 a. C. y el 3.800 a. C. era portador de R1a y del U2e1 materno . [66] Según el arqueólogo David Anthony, el R1a-Z93 paterno fue encontrado en el río Oskol cerca de un koljoz "Alejandría" que ya no existe, Ucrania c. 4000 a. C., "la muestra más antigua conocida que muestra la adaptación genética a la persistencia de la lactasa (13910-T)". [67] R1a se ha encontrado en la cultura Corded Ware , [68] [69] en la que es predominante. [70] Los machos examinados de la cultura Fatyanovo de la Edad del Bronce pertenecen enteramente al R1a, específicamente al subclado R1a-Z93. [62] [63] [71]
Posteriormente se ha encontrado el haplogrupo R1a en fósiles antiguos asociados con la cultura Urnfield ; [72] así como el entierro de los restos de las culturas Sintashta , [16] Andronovo , [73] Pazyryk , [74] Tagar , [73] Tashtyk , [ 73] y Srubnaya , los habitantes de la antigua Tanais , [ 75] en las momias de Tarim , [76] y la aristocracia de Xiongnu . [77] Los restos esqueléticos de un padre y sus dos hijos, de un sitio arqueológico descubierto en 2005 cerca de Eulau (en Sajonia-Anhalt , Alemania ) y que datan aproximadamente del 2600 a. C., dieron positivo para el marcador Y-SNP SRY10831.2. El número de búsqueda de los restos de Eulau es 2C46S. Por tanto, el clado ancestral estuvo presente en Europa hace al menos 4600 años, en asociación con un sitio de la extendida cultura Corded Ware . [68]
Hay una presencia importante en pueblos de ascendencia escandinava , con niveles más altos en Noruega e Islandia , donde entre el 20 y el 30% de los hombres están en R1a1a. [84] [85] Los vikingos y normandos también pueden haber llevado el linaje R1a1a más lejos, representando al menos parte de la pequeña presencia en las Islas Británicas , las Islas Canarias y Sicilia . [86] [87] En Alemania Oriental , el haplogrupo R1a1a promedia entre el 20 y el 30%, con un pico en Rostock del 31,3%. [88]
En el sur de Europa, R1a1a no es común, pero se han encontrado niveles significativos en zonas como el Valle del Pas en el norte de España , áreas de Venecia y Calabria en Italia . [89] [ se necesita mejor fuente ] Los Balcanes muestran una amplia variación entre áreas con niveles significativos de R1a1a, por ejemplo, 36-39% en Eslovenia , [90] 27-34% en Croacia , [81] [91] [92] [ 93] [94] y más del 30% en la Macedonia griega , pero menos del 10% en Albania , Kosovo y partes de Grecia al sur del desfiladero del Olimpo. [95] [82] [12]
R1a está prácticamente compuesto únicamente por el subclado Z284 en Escandinavia . En Eslovenia, el subclado principal es Z282 (Z280 y M458), aunque el subclado Z284 se encontró en una muestra de un esloveno. Hay una representación insignificante de Z93 en Turquía , el 12,1% [60] [2] Los eslavos occidentales y los húngaros se caracterizan por una alta frecuencia del subclado M458 y una baja Z92, un subclado de Z280. Cientos de muestras eslovenas y checas carecen del subclado Z92 de Z280, mientras que los polacos, eslovacos, croatas y húngaros sólo muestran una frecuencia muy baja de Z92. [2] Los bálticos , los eslavos orientales , los serbios , los macedonios , los búlgaros y los rumanos presentan una proporción Z280>M458 y una proporción alta, hasta predominante, de Z92. [2] Los bálticos y los eslavos orientales tienen los mismos subclados y frecuencias similares en una filogenia más detallada de los subclados. [96] [97] El genetista ruso Oleg Balanovsky especuló que existe un predominio del sustrato preeslavo asimilado en la genética de las poblaciones eslavas orientales y occidentales, según él, la estructura genética común que contrasta a los eslavos orientales y bálticos de otras poblaciones. puede sugerir la explicación de que el sustrato preeslavo de los eslavos orientales y occidentales estaba formado principalmente por hablantes del Báltico, que en un momento fueron anteriores a los eslavos en las culturas de la estepa euroasiática según referencias arqueológicas y toponímicas. [nota 16]
Asia
Asia Central
Zerjal et al. (2002) encontraron R1a1a en el 64% de una muestra de tayikos de Tayikistán y en el 63% de una muestra de kirguís de Kirguistán . [98]
Haber et al. (2012) encontraron R1a1a-M17 en el 26,0% (53/204) de un conjunto de muestras de Afganistán , incluido el 60% (3/5) de una muestra de nuristanis , el 51,0% (25/49) de una muestra de pastunes, 30,4% (17/56) de una muestra de tayikos, 17,6% (3/17) de una muestra de uzbekos, 6,7% (4/60) de una muestra de hazaras y en el único individuo turcomano de la muestra. [99]
Di Cristofaro et al. (2013) encontraron R1a1a-M198/M17 en el 56,3% (49/87) de un par de muestras de pastunes de Afganistán (incluido 20/34 o 58,8% de una muestra de pastunes de Baghlan y 29/53 o 54,7% de una muestra de pastunes de Kunduz ), 29,1% (37/127) de un conjunto de muestras de uzbekos de Afganistán (incluido 28/94 o 29,8% de una muestra de uzbekos de Jawzjan , 8/28 o 28,6% de una muestra de uzbekos de Sar-e Pol , y 1/5 o 20% de una muestra de uzbekos de Balkh ), el 27,5% (39/142) de un conjunto de muestras de tayikos de Afganistán (incluidos 22/54 o 40,7% de una muestra de Tayikos de Balkh , 9/35 o 25,7% de una muestra de tayikos de Takhar , 4/16 o 25,0% de una muestra de tayikos de Samangan , y 4/37 o 10,8% de una muestra de tayikos de Badakhshan ), 16,2% (12/74) de una muestra de turcomanos de Jawzjan , y el 9,1% (7/77) de un par de muestras de hazara de Afganistán (incluido 7/69 o 10,1% de una muestra de hazara de Bamiyan y 0/8 o 0% de una muestra de Hazara de Balkh ). [100]
Malyarchuk et al. (2013) encontraron R1a1-SRY10831.2 en el 30,0% (12/40) de una muestra de tayikos de Tayikistán. [101]
Ashirbekov et al. (2017) encontraron R1a-M198 en el 6,03% (78/1294) de un conjunto de muestras de kazajos de Kazajstán . R1a-M198 se observó con una frecuencia superior a la media en las muestras del estudio de las siguientes tribus kazajas: 13/41 = 31,7 % de una muestra de Suan, 8/29 = 27,6 % de una muestra de Oshaqty, 6/30 = 20,0 % de una muestra de Qozha, 4/29 = 13,8% de una muestra de Qypshaq, 1/8 = 12,5% de una muestra de Tore, 9/86 = 10,5% de una muestra de Jetyru, 4/50 = 8,0% de una muestra de Argyn, 1/13 = 7,7% de una muestra de Shanyshqyly, 8/122 = 6,6% de una muestra de Alimuly, 3/46 = 6,5% de una muestra de Alban. R1a-M198 también se observó en 5/42 = 11,9% de una muestra de kazajos de afiliación tribal no declarada. [102]
Asia del Sur
En el sur de Asia, R1a1a se ha observado a menudo en varios grupos demográficos. [26] [25]
Además de estos, los estudios muestran altos porcentajes en grupos regionalmente diversos como los Manipuris (50%) [3] en el extremo noreste y entre los punjabis (47%) [20] en el extremo noroeste.
Un artículo chino publicado en 2018 encontró R1a-Z94 en el 38,5% (15/39) de una muestra de uigures keriyalik de Darya Boyi / Darya Boye Village, condado de Yutian , Xinjiang (于田县达里雅布依乡), R1a-Z93. en el 28,9% (22/76) de una muestra de Dolan uigures del municipio de Horiqol, condado de Awat , Xinjiang (阿瓦提县乌鲁却勒镇), y R1a-Z93 en el 6,3% (4/64) de una muestra de Loplik uigures de Karquga / Qarchugha Village, condado de Yuli , Xinjiang (尉犁县喀尔曲尕乡). R1a(xZ93) se observó sólo en uno de los 76 uigures de Dolan. [109] Tenga en cuenta que Darya Boyi Village está ubicada en un oasis remoto formado por el río Keriya en el desierto de Taklamakan . Un estudio de ADN-Y de 2011 encontró Y-dna R1a1 en el 10% de una muestra de personas Hui del sur de Yunnan, el 1,6% de una muestra de personas tibetanas de Xizang ( Región Autónoma del Tíbet ), el 1,6% de una muestra de personas Xibe de Xinjiang , 3,2% de una muestra de hui del norte de Ningxia , 9,4% de una muestra de hazak ( kazajos ) de Xinjiang, y tasas de 24,0%, 22,2%, 35,2%, 29,2% en 4 muestras diferentes de uigures de Xinjiang, 9,1% en una muestra de mongoles de Mongolia Interior . También se encontró un subclado diferente de R1 en el 1,5% de una muestra del norte de Hui de Ningxia. [110] en el mismo estudio no se detectaron casos de R1a en 6 muestras de chinos Han en Yunnan , 1 muestra de Han en Guangxi , 5 muestras de Han en Guizhou , 2 muestras de Han en Guangdong , 2 muestras de Han en Fujian , 2 muestras de Han en Zhejiang , 1 muestra de Han en Shanghai , 1 muestra de Han en Jiangxi , 2 muestras de Han en Hunan , 1 muestra de Han en Hubei , 2 muestras de Han en Sichuan , 1 muestra de Han en Chongqing , 3 muestras de Han en Shandong , 5 muestras de Han en Gansu , 3 muestras de Han en Jilin y 2 muestras de Han en Heilongjiang . [111] 40% de los salars, 45,2% de los tayikos de Xinjiang , 54,3% de los dongxiang, 60,6% de los tártaros y 68,9% de los kirguís.En Xinjiang, en el noroeste de China, probado en una muestra tenía R1a1-M17. Bao'an (Bonan) tenía la mayor diversidad de haplogrupos de 0,8946 ± 0,0305, mientras que las otras minorías étnicas en el noroeste de China tenían una alta diversidad de haplogrupos como los asiáticos centrales, de 0,7602 ± 0,0546. [112]
En Siberia Oriental , R1a1a se encuentra entre ciertos grupos étnicos indígenas, incluidos los kamchatkanos y chukotkanos , y alcanza un máximo en Itel'man con un 22%. [113]
Asia occidental
R1a1a se ha encontrado en diversas formas, en la mayor parte de Asia occidental , en concentraciones muy variables, desde casi ninguna presencia en áreas como Jordania , hasta niveles mucho más altos en partes de Kuwait e Irán . La tribu beduina Shimar ( Shammar ) de Kuwait muestra la frecuencia más alta en Oriente Medio con un 43%. [114] [115] [116]
Wells 2001 señaló que en la parte occidental del país, los iraníes muestran niveles bajos de R1a1a, mientras que los hombres de las partes orientales de Irán portaban hasta un 35% de R1a1a. Nasidze et al. 2004 encontró R1a1a en aproximadamente el 20% de los hombres iraníes de las ciudades de Teherán e Isfahán . Regueiro 2006, en un estudio de Irán , observó frecuencias mucho más altas en el sur que en el norte.
Un estudio más reciente ha encontrado un 20,3% de R-M17* entre muestras kurdas tomadas en la provincia de Kurdistán en el oeste de Irán, un 19% entre azerbaiyanos en Azerbaiyán occidental , un 9,7% entre mazandaraníes en el norte de Irán en la provincia de Mazandaran , un 9,4% entre gilaks. en la provincia de Gilan , 12,8% entre los persas y 17,6% entre los zoroastrianos en Yazd , 18,2% entre los persas en Isfahan , 20,3% entre los persas en Khorasan , 16,7% afroiraníes, 18,4% Qeshmi "Gheshmi", 21,4% entre el pueblo persa Bandari en Hormozgan y el 25% entre los baluchis en la provincia de Sistán y Baluchistán . [117]
Di Cristofaro et al. (2013) encontraron el haplogrupo R1a en el 9,68% (18/186) de un conjunto de muestras de Irán, aunque con una gran variación que va desde el 0% (0/18) en una muestra de iraníes de Teherán hasta el 25% (5/20 ) en una muestra de iraníes de Khorasan y el 27% (3/11) en una muestra de iraníes de procedencia desconocida. Todos los individuos iraníes R1a portaban las mutaciones M198 y M17, excepto un individuo en una muestra de iraníes de Gilan ( n = 27), que se informó que pertenecía a R1a-SRY1532.2 (xM198, M17). [100]
Malyarchuk et al. (2013) encontraron R1a1-SRY10831.2 en el 20,8% (16/77) de una muestra de persas recolectada en las provincias de Khorasan y Kerman en el este de Irán, pero no encontraron ningún miembro de este haplogrupo en una muestra de 25 kurdos. recolectados en la provincia de Kermanshah en el oeste de Irán. [101]
Más al norte de estas regiones de Asia occidental, por el contrario, los niveles de R1a1a comienzan a aumentar en el Cáucaso , también de forma desigual. Varias poblaciones estudiadas no han mostrado signos de R1a1a, mientras que los niveles más altos descubiertos hasta ahora en la región parecen pertenecer a hablantes de la lengua Karachay-Balkar, entre los cuales aproximadamente una cuarta parte de los hombres examinados hasta ahora están en el haplogrupo R1a1a. [3]
Denominación histórica de R1a
El sistema de denominación histórico comúnmente utilizado para R1a era inconsistente en diferentes fuentes publicadas, porque cambiaba con frecuencia; esto requiere alguna explicación.
En 2002, el Consorcio del Cromosoma Y (YCC) propuso un nuevo sistema de denominación para haplogrupos (YCC 2002), que ahora se ha convertido en estándar. En este sistema, los nombres con el formato "R1" y "R1a" son nombres " filogenéticos ", destinados a marcar posiciones en un árbol genealógico. Los nombres de las mutaciones de SNP también se pueden utilizar para nombrar clados o haplogrupos. Por ejemplo, como M173 es actualmente la mutación que define a R1, R1 también es R-M173, un nombre de clado "mutacional". Cuando se descubre una nueva ramificación en un árbol, algunos nombres filogenéticos cambiarán, pero por definición todos los nombres mutacionales seguirán siendo los mismos.
El haplogrupo de amplia aparición definido por la mutación M17 se conocía con varios nombres, como "Eu19", como se utiliza en (Semino et al. 2000) en los sistemas de denominación más antiguos. La propuesta del YCC de 2002 asignó el nombre R1a al haplogrupo definido por la mutación SRY1532.2. Esto incluía a Eu19 (es decir, R-M17) como subclado, por lo que Eu19 se denominó R1a1. Tenga en cuenta que SRY1532.2 también se conoce como SRY10831.2 [ cita necesaria ] El descubrimiento de M420 en 2009 ha provocado una reasignación de estos nombres filogenéticos (Underhill et al. 2009 e ISOGG 2012) R1a ahora está definido por la mutación M420: en este árbol actualizado, el subclado definido por SRY1532.2 se ha movido de R1a a R1a1, y Eu19 (R-M17) de R1a1 a R1a1a.
Las actualizaciones más recientes registradas en la página web de referencia de ISOGG involucran sucursales de R-M17, incluida una sucursal principal, R-M417.
^ Según Family Tree, [ ¿quién? ] se diversificaron c. Hace 5.000 años. [9]
^ Ver también: "dispersiones de población 'fuertemente sesgadas por sexo' en el subcontinente indio (Silva et al. 2017)" . Blog de Eurogenes . 28 de marzo de 2017.[ fuente autoeditada? ]
^ Kivisild y col. (2003): "El haplogrupo R1a, anteriormente asociado con la supuesta invasión indo-aria, se encontró en su mayor frecuencia en Punjab, pero también con una frecuencia relativamente alta (26%) en la tribu Chenchu. Este hallazgo, junto con el mayor R1a "La diversidad de repeticiones en tándem cortas asociadas en India e Irán en comparación con Europa y Asia central, sugiere que el sur y el oeste de Asia podrían ser la fuente de este haplogrupo". [20]
^
Sahoo y col. (2006): "... uno debería esperar observar una variación genética dramáticamente menor entre los linajes Rla indios. De hecho, ocurre lo contrario: la diversidad de haplotipos STR en el contexto de R1a en Asia Central (y también en Europa del Este) ha Ya se ha demostrado que es menor que en la India (6). Más bien, la alta incidencia de R1* y Rla en las poblaciones europeas de Asia Central (sin R2 y R* en la mayoría de los casos) se explica de manera más parsimoniosa por el flujo de genes en la dirección opuesta. , posiblemente con un efecto fundador temprano en el sur o el oeste de Asia [28] .
^
Sharma y col. (2009): "Una observación peculiar de la frecuencia más alta (hasta 72,22%) del haplogrupo Y R1a1* en los brahmanes insinuó su presencia como un linaje fundador de este grupo de castas. Además, la observación de R1a1* en diferentes grupos de población tribales , la existencia del haplogrupo Y R1a* en ancestros y los análisis filogenéticos extendidos del conjunto de datos agrupados de 530 indios, 224 paquistaníes y 276 asiáticos centrales y euroasiáticos que portaban el haplogrupo R1a1* respaldaron el origen autóctono del linaje R1a1 en la India y un vínculo tribal con el linaje indio. Brahmanes. Sin embargo, es importante descubrir nuevos marcadores binarios del cromosoma Y para una resolución más alta de R1a1* y confirmar las presentes conclusiones". [1]
^ durante la mayor parte de la historia, el subcontinente indio fue la región más poblada del mundo
^ Sengupta y col. (2006): "La distribución geográfica generalizada de HG R1a1-M17 en Eurasia y la ausencia actual de subdivisiones informativas definidas por marcadores binarios dejan incierto el origen geográfico de HG R1a1-M17. Sin embargo, el mapa de contorno de la varianza de R1a1-M17 muestra la mayor variación en la región noroeste de la India... La pregunta sigue siendo qué tan distintiva es la historia de L1 en relación con algunos o todos los representantes de R1a1 y R2. Esta incertidumbre neutraliza las conclusiones previas de que la intrusión de los HG R1a1 y R2 desde el noroeste en Las tribus del sur de habla dravídica se pueden atribuir a un único evento reciente. [R1a1 y R2] podrían haber llegado al sur de la India desde una región de origen del suroeste de Asia varias veces, con algunos episodios considerablemente antes que otros. Existe evidencia arqueológica considerable sobre la presencia de Pueblos mesolíticos de la India (Kennedy 2000), algunos de los cuales podrían haber ingresado al subcontinente desde el noroeste durante la época del Pleistoceno tardío. La alta varianza de R1a1 en la India (tabla 12), la distribución de frecuencia espacial de las líneas de varianza de los microsatélites R1a1 (fig. 4) y el tiempo de expansión (tabla 11) respaldan esta opinión". [25 ]
^ Semenov y Bulat (2016) hacen referencia a las siguientes publicaciones:
Haak, Wolfgang (2015). "La migración masiva desde la estepa es una fuente de lenguas indoeuropeas en Europa". Naturaleza . 522 (7555): 207–211. arXiv : 1502.02783 . Código Bib :2015Natur.522..207H. bioRxiv 10.1101/013433 . doi : 10.1038/NATURALEZA14317 . PMC 5048219 . PMID 25731166.
Mathieson, Iain (2015). "Ocho mil años de selección natural en Europa". bioRxiv 10.1101/016477 .
Chekunova Е.М., Yartseva NV, Chekunov М.К., Мazurkevich А.N. Los primeros resultados del genotipado de los aborígenes y los restos óseos humanos de los monumentos arqueológicos del Alto Podvin'e. // Arqueología de los asentamientos lacustres del IV—II milenio antes de Cristo: la cronología de las culturas y el entorno natural y los ritmos climáticos. Actas de la conferencia internacional dedicada a la investigación de 50 años de los asentamientos de pilotes en el noroeste de Rusia. San Petersburgo, 13 al 15 de noviembre de 2014.
Jones, ER; González-Fortes, G; Connell, S; Siska, V; Eriksson, A; Martiniano, R; McLaughlin, RL; Gallego Llorente, M; Cassidy, LM; Gamba, C; Meshveliani, T; Bar-Yosef, O; Müller, W; Belfer-Cohen, A; Matskevich, Z; Jakeli, N; Higham, TF; Currat, M; Lordkipanidze, D; Hofreiter, M; Manica, A; Pinhasi, R; Bradley, Director General (2015). "Los genomas del Paleolítico superior revelan raíces profundas de los euroasiáticos modernos". Comuna Nacional . 6 : 8912. Código Bib : 2015NatCo...6.8912J. doi : 10.1038/ncomms9912. PMC 4660371 . PMID 26567969.
^ Sin embargo, Haak et al. También declara explícitamente: "un tipo de ascendencia del Cercano Oriente diferente de la que introdujeron los primeros agricultores". [ se necesita aclaración ] [39]
^ Según Family Tree DNA, L664 se formó hace 4700 años, es decir, 2700 a. [9]
^ Lazaridis, Twitter, 18 de junio de 2016: "I1635 (Armenia_EBA) es R1b1-M415(xM269). Nos aseguraremos de incluirlo en la revisión. ¡Gracias a la persona que se dio cuenta! #ILovePreprints". [ fuente poco confiable? ] Véase también "El gran problema de 2016: el territorio del actual Irán no puede ser la patria indoeuropea". Blog de Eurogenes . 26 de noviembre de 2016,[ fuente poco confiable? ] para una discusión sobre el mismo tema.
^ Consulte el mapa de distribución de M780 en el blog de antropología de Dieneke, nuevo artículo importante sobre los orígenes profundos del haplogrupo Y R1a (Underhill et al. 2014) [43]
^ Según Family Tree DNA, M780 se formó hace 4700 ybp. [9] Esta datación coincide con el movimiento hacia el este entre 2800 y 2600 a. C. de la cultura Yamnaya hacia la región de la cultura Poltavka , predecesora de la cultura Sintashta , de la que se originaron los indoiraníes. M780 se concentra en el valle del Ganges, el lugar de la sociedad védica clásica .
^ Poznik y col. (2016) calculan con un tiempo de generación de 30 años; un tiempo de generación de 20 años produce otros resultados.
^ "La evidencia de que el grupo Steppe_MLBA [Edad del Bronce Medio a Final] es una fuente plausible de la ascendencia esteparia en el sur de Asia también está respaldada por evidencia del cromosoma Y, como el haplogrupo R1a, que es del subtipo Z93 común en el sur de Asia hoy en día [Underhill et al. (2014), Silva et al. (2017)] fue de alta frecuencia en Steppe_MLBA (68%) (16), pero raro en Steppe_EMBA [Edad del Bronce Temprano a Medio] (ausente en nuestros datos)". [44]
^ Балановский (2015), pág. 208 (en ruso) Прежде всего, это преобладание в славянских популяциях дославянского субстрата — двух ассимилированных ими генетиче ских компонентов – восточноевропейского для западных и восточных славян и южноевропейского для южных славян... предположить, что ассимилированный субстратмог быть представлен по преимуществу балтоязычными популяциями . Actualmente, la arqueología está formada por un grupo de expertos de la comunidad esclava. Балтскийсубстрату славян (правда, наряду с финно-угорским) выявляли а антропологи. Полученные нами генетические данные — и на графиках генетических взаимоотношений, и по доле общих фрагментов генома — указывают, что современные балтские народы являются ближайшими генетически ми соседями восточных славян. При этом балты являются и лингвистически ближайшими род ственниками славян. И можно полагать, что к моменту ассимиляции и х генофонда не так сильно отличался от генофонда начавших свое широкое расселе ние славян. Después de todo, es necesario asimilar el uso de baltos de primera calidad, por lo que es posible comprobarlo. и сходство современных славянских и балтских народов друг с другом, и и и отличия от окружающих их не балто-славянских гр упп Европы...В работе высказывается осторожное предположение, что El sustrato asimilado puede ser muy popular entre las personas más populares. Actualmente, la arqueología está integrada por un grupo de investigación balcánico formado por un grupo de expertos eslavos. Балтский субстрат у славян (правда, наряду с финно-угорским) выявляли и антропологи. Полученные в этой работе генетические данные — и на графиках генетических взаимоотношений, и по доле общих фрагментов генома — указывают, что современные балтские народы являются ближайшими генетическими соседями восточных славян.
Referencias
^ abcdef Sharma y col. 2009.
^ abcdefghijklmnopqrstu Underhill et al. 2014.
^ abcdefghijklmnopqr Underhill et al. 2009.
^ ab Underhill y col. 2014, pág. 130.
^ Saag, Lehti; Vasiliev, Sergey V.; Varul, Liivi; Kosorukova, Natalia V.; Gerasimov, Dmitri V.; Oshibkina, Svetlana V.; Griffith, Samuel J.; Solnik, Anu; Saag, Lauri; D'Atanasio, Eugenia; Metspalu, Ene (enero de 2021). "Cambios de ascendencia genética en la transición de la Edad de Piedra a la Edad del Bronce en la llanura de Europa del Este". Avances científicos . 7 (4): eabd6535. Código Bib : 2021SciA....7.6535S. doi :10.1126/sciadv.abd6535. PMC 7817100 . PMID 33523926.
^ Haak, Wolfgang; Lazaridis, José; Patterson, Nick; Rohland, Nadin; Mallick, Swapan; Llamas, Bastien; Brandt, Guido; Nordenfelt, Susanne; Harney, Eadaoin; Stewardson, Kristin; Fu, Qiaomei (10 de febrero de 2015). "La migración masiva desde la estepa es una fuente de lenguas indoeuropeas en Europa". bioRxiv : 013433. arXiv : 1502.02783 . doi :10.1101/013433. S2CID 196643946.
^ Raghavan, Maanasa; Skoglund, Ponto; Graf, Kelly E.; Metspalu, Mait; Albrechtsen, Anders; Moltke, Ida; Rasmussen, Simón; Stafford Jr, Thomas W.; Orlando, Ludovic; Metspalu, Ene; Karmin, Monika (enero de 2014). "El genoma del Paleolítico superior de Siberia revela una doble ascendencia de los nativos americanos". Naturaleza . 505 (7481): 87–91. Código Bib :2014Natur.505...87R. doi : 10.1038/naturaleza12736. PMC 4105016 . PMID 24256729.
^ Narasimhan, Vagheesh M.; Patterson, Nick; Moorjani, Priya; Rohland, Nadin; Bernardos, Rebeca; Mallick, Swapan; Lazaridis, José; Nakatsuka, Nathan; Olalde, Íñigo; Lipson, Marcos; Kim, Alexander M. (6 de septiembre de 2019). "La formación de poblaciones humanas en Asia central y meridional". Ciencia . 365 (6457): comer7487. doi : 10.1126/science.aat7487. PMC 6822619 . PMID 31488661. Los tipos de haplogrupo del cromosoma Y R1b o R1a no están representados en Irán y Turan en este período ...
^ abcdefghij "árbol R1a". YCompleto .
^ abc Mirabal et al. 2009.
^ Zerjal, T.; et al. (1999). "El uso de la variación del ADN del cromosoma Y para investigar la historia de la población: reciente propagación masculina en Asia y Europa". En Papiha, SS; Deka, R. y Chakraborty, R. (eds.). Diversidad genómica: aplicaciones en genética de poblaciones humanas . Nueva York: Kluwer Academic/Plenum Publishers. págs. 91-101. ISBN978-0-3064-6295-5.
^ abcd Semino y col. 2000.
^ ab Wells 2001.
^ a b C Pamjav et al. 2012.
^ ab Haak y col. 2015.
^ ab Allentoft y col. 2015.
^ ab Mathieson y col. 2015.
^ ab Silva y col. 2017.
^ ab Joseph, Tony (16 de junio de 2017). "Cómo la genética está zanjando el debate sobre la migración aria". El hindú .
^ abcd Kivisild et al. 2003.
^ Sengupta S, Zhivotovsky LA, King R, Mehdi SQ, Edmonds CA, Chow CE, et al. (febrero de 2006). "La polaridad y la temporalidad de las distribuciones del cromosoma y de alta resolución en la India identifican expansiones tanto indígenas como exógenas y revelan una influencia genética menor de los pastores de Asia Central". Revista Estadounidense de Genética Humana . 78 (2): 202–221. doi :10.1086/499411. PMC 1380230 . PMID 16400607. "Aunque el impacto cultural considerable en la jerarquía social y el lenguaje en el sur de Asia es atribuible a la llegada de pastores nómadas de Asia Central, los datos genéticos (mitocondrial y cromosómico Y) han producido inferencias dramáticamente contradictorias sobre los orígenes genéticos de las tribus y castas del sur de Asia. Intentamos resolver este conflicto utilizando datos de alta resolución sobre 69 marcadores binarios informativos del cromosoma Y y 10 marcadores microsatélites de un gran conjunto de grupos étnicos geográfica, social y lingüísticamente representativos del sur de Asia. "El acervo genético preexistente era menor. Las edades de variación microsatélite acumulada en la mayoría de los haplogrupos indios superan los 10.000 a 15.000 años, lo que atestigua la antigüedad de la diferenciación regional. Por lo tanto, nuestros datos no respaldan modelos que invoquen una pronunciada genética reciente. Los análisis de microsatélites asociados de los cromosomas del haplogrupo R1A1 de alta frecuencia indican historias recientes independientes del valle del INDUS y de Asia central para explicar la variación genética observada en el sur de Asia. LA REGIÓN INDÍGENA PENINSULAR."
^ Thanseem I, Thangaraj K, Chaubey G, Singh VK, Bhaskar LV, Reddy BM y otros. (Agosto de 2006). "Afinidades genéticas entre las castas inferiores y grupos tribales de la India: inferencia del cromosoma Y y el ADN mitocondrial". Genética BMC . 7 : 42. doi : 10.1186/1471-2156-7-42 . PMC 1569435 . PMID 16893451.
^ Sahoo S, Singh A, Himabindu G, Banerjee J, Sitalaximi T, Gaikwad S, et al. (Enero de 2006). "Una prehistoria de los cromosomas Y indios: evaluación de escenarios de difusión démica". Actas de la Academia Nacional de Ciencias de los Estados Unidos de América . 103 (4): 843–848. Código Bib : 2006PNAS..103..843S. doi : 10.1073/pnas.0507714103 . PMC 1347984 . PMID 16415161.
^ Thangaraj K, Naidu BP, Crivellaro F, Tamang R, Upadhyay S, Sharma VK y col. (Diciembre de 2010). Cordaux R (ed.). "La influencia de las barreras naturales en la configuración de la estructura genética de las poblaciones de Maharashtra". MÁS UNO . 5 (12): e15283. Código Bib : 2010PLoSO...515283T. doi : 10.1371/journal.pone.0015283 . PMC 3004917 . PMID 21187967.
^ abcdef Sengupta 2006.
^ abcde Sahoo y col. 2006.
^ ab Thangaraj et al. 2010.
^ Sahoo y col. 2006, pág. 845-846.
^ Pande, A. (2022). Control de la natalidad: reproducción selectiva y eugenesia neoliberal en Sudáfrica e India. Gobernando las intimidades en el Sur Global. Prensa de la Universidad de Manchester. pag. 3-PA1976-3-PA1979. ISBN978-1-5261-6053-9. Consultado el 16 de julio de 2023 ."La perspectiva dominante es que los R1a se originaron 'fuera' de la India y fueron llevados al país por un grupo de inmigrantes ('los arios') que llegaron después de la civilización del valle del Indo".
^ Lalueza-Fox, C. (2022). Desigualdad: una historia genética. Prensa del MIT. págs. 81–82. ISBN978-0-262-04678-7. Consultado el 16 de julio de 2023 ."Hace algunos años, los científicos locales apoyaron la opinión de que la existencia de un cromosoma Y R1a no era atribuible a un flujo de genes extraños, sino que este linaje había surgido en el subcontinente y se había extendido desde allí. Pero la reconstrucción filogenética de este haplogrupo no apoyar esta opinión."
^ Antonio 2007.
^ Antonio y Ringe 2015.
^ ab Haak y col. 2015, pág. 5.
^ Semenov y Bulat 2016.
^ Haber y col. 2012 "R1a1a7-M458 estuvo ausente en Afganistán, lo que sugiere que R1a1a-M17 no admite, como se pensaba anteriormente [47], expansiones desde la estepa póntica [3], llevando las lenguas indoeuropeas a Asia central y la India".
^ Klejn, Leo S. (22 de abril de 2017). "La hipótesis de la estepa de orígenes indoeuropeos aún no se ha demostrado". Acta Arqueológica . 88 (1): 193–204. doi :10.1111/j.1600-0390.2017.12184.x. ISSN 0065-101X."En cuanto al cromosoma Y, ya se observó en Haak, Lazaridis et al. (2015) que los Yamnaya de Samara tenían cromosomas Y que pertenecían al R-M269 pero no pertenecían al clado común en Europa occidental (p. . 46 del suplemento). Además, ni un solo R1a en Yamnaya a diferencia de Corded Ware (dominado por R1a)".
^ Koch, John T.; Cunliffe, Barry (2016). Celtas de Occidente 3: la Europa atlántica en la Edad de los Metales. Libros Oxbow. pag. 634.ISBN _978-1-78570-228-0.
^ Semenov y Bulat 2016, pag. 41.
^ Haak y col. 2015, pág. 4.
^ abc Mascarenhas et al. 2015, pág. 9.
^ ab Poznik et al. 2016, pág. 5.
^ Blog en inglés de Arame, Y ADN del antiguo Cercano Oriente
^ "Blog de antropología de Dienekes: nuevo artículo importante sobre los orígenes profundos del haplogrupo Y R1a (Underhill et al. 2014)". 27 de marzo de 2014 . Consultado el 20 de diciembre de 2019 .[ fuente poco confiable? ]
^ ab Narasimhan et al. 2018.
^ a b "Acerca de nosotros". ADN del árbol genealógico . Consultado el 20 de diciembre de 2019 .
^ abcdefghijklmnopqrstu vwxyz aa "ISOGG 2017 Y-DNA Haplogroup R". isogg.org . Consultado el 20 de diciembre de 2019 .
^ abcdefghijk "Haplogrupo R (ADN-Y) - SNPedia". www.snpedia.com . Consultado el 20 de diciembre de 2019 .
^ Karafet y otros. 2014.
^ abcdefghijklm Underhill y col. 2014, pág. 125.
^ "R1a en Yamnaya". Blog de Eurogenes . 21 de marzo de 2016. Archivado desde el original el 5 de mayo de 2018 . Consultado el 20 de diciembre de 2019 .
^ "Y-DNA Haplogrupo R y sus subcladas". Sociedad Internacional de Genealogía Genética (ISOGG) . Consultado el 8 de enero de 2011 .
^ Krahn, Thomas. "Borrador de árbol del cromosoma Y". ADN del árbol genealógico . Archivado desde el original el 26 de mayo de 2013 . Consultado el 7 de diciembre de 2012 .
^ Regueiro 2006.
^ Freder, Janine (2010). Die mittelalterlichen Skelette von Usedom: Anthropologische Bearbeitung unter besonderer Berücksichtigung des ethnischen Hintergrundes [ Investigación antropológica teniendo debidamente en cuenta el origen étnico ] (Tesis) (en alemán). Universidad Libre de Berlín. pag. 86. doi :10.17169/refubium-8995.
^ Underhill, Pensilvania; et al. (2009). "Separación de la coascendencia posglacial de los cromosomas y europeos y asiáticos dentro del haplogrupo R1a". Revista europea de genética humana . 18 (4): 479–484. doi :10.1038/ejhg.2009.194. PMC 2987245 . PMID 19888303.
^ Gwozdz, Peter (6 de agosto de 2018). "Clados polacos de ADN-Y".
^ Pawlowski y col. 2002.
^ ab Gwozdz 2009.
^ ab Kars, YO; Başak, AN; Onat, OE; Bilguvar, K.; Choi, J.; Itán, Y.; Çağlar, C.; Palvadeau, R.; Casanova, JL; Cooper, DN; Stenson, PD; Yavuz, A.; Buluş, H.; Gunel, M.; Friedman, JM; Özçelik, T. (2021). "La estructura genética de la población turca revela altos niveles de variación y mezcla". Actas de la Academia Nacional de Ciencias de los Estados Unidos de América . 118 (36): e2026076118. Código Bib : 2021PNAS..11826076K. doi : 10.1073/pnas.2026076118 . PMC 8433500 . PMID 34426522.
^ Petrejcíková, EVA; Soták, Miroslav; Bernasovská, Jarmila; Bernasovský, Ivan; Sovicová, Adriana; Bôziková, Alexandra; Boronová, Iveta; Švícková, Petra; Gabriková, Dana; MacEková, Sona (2009). "Frecuencias del haplogrupo Y en la población romaní eslovaca". Ciencia Antropológica . 117 (2): 89–94. doi : 10.1537/ase.080422 .
^ abc Saag y col. 2020, pág. 5.
^ ab Saag y col. 2020, pág. 29, Tabla 1.
^ Saag y col. 2020, Datos complementarios 2, Fila 4.
^ Fu y col. 2016.
^ Saag y col. 2017.
^ Antonio 2019, págs.16, 17.
^ ab Haak y col. 2008.
^ Brandit y col. 2013.
^ Malmström y col. 2019, pág. 2.
^ Saag y col. 2020, Datos complementarios 2, filas 5-49.
^ Schweitzer, D. (23 de marzo de 2008). "Análisis de datos de la cueva de Lichtenstein" (PDF) . dirkschweitzer.net. Archivado desde el original (PDF) el 14 de agosto de 2011.Resumen en inglés de Schilz (2006).
^ abc Keyser y col. 2009.
^ Ricaut y col. 2004.
^ Korniyenko, IV; Vodolazhsky DI "Использование нерекомбинантных маркеров Y-хромосомы в исследованиях древних популяций (на примере поселения Танаис)" [El uso de no recomendados marcadores binantes del cromosoma Y en el estudio de poblaciones antiguas (en el ejemplo del asentamiento de Tanais)] . Материалы Донских антропологических чтений [Materiales de las lecturas antropológicas del Don]. Rostov del Don: Instituto de Investigación de Oncología de Rostov, 2013.
^ Chunxiang Li y otros. 2010.
^ Kim y col. 2010.
^ Balanovsky y col. 2008.
^ Behar y col. 2003.
^ Kasperaviciūte, Kucinskas y Stoneking 2005.
^ ab Battaglia et al. 2008.
^ ab Rosser y col. 2000.
^ Tambets y col. 2004.
^ Bowden y col. 2008.
^ Dupuy y otros. 2005.
^ Passarino y col. 2002.
^ Capelli y col. 2003.
^ Kayser y col. 2005.
^ Scozzari y col. 2001.
^ Underhill, Peter A. (1 de enero de 2015). "La estructura filogenética y geográfica del haplogrupo R1a del cromosoma Y". Revista europea de genética humana . 23 (1): 124-131. doi :10.1038/ejhg.2014.50. PMC 4266736 . PMID 24667786.
^ L. Barac; et al. (2003). "Herencia cromosómica Y de la población croata y sus aislamientos insulares" (PDF) . Revista europea de genética humana . 11 (7): 535–42. doi : 10.1038/sj.ejhg.5200992 . PMID 12825075. S2CID 15822710.
^ S. Rootsi; et al. (2004). "La filogeografía del haplogrupo I del cromosoma Y revela dominios distintos del flujo de genes prehistóricos en Europa" (PDF) . Revista Estadounidense de Genética Humana . 75 (1): 128-137. doi :10.1086/422196. PMC 1181996 . PMID 15162323.
^ M. Pericic; et al. (2005). "El análisis filogenético de alta resolución del sureste de Europa rastrea episodios importantes de flujo de genes paternos entre las poblaciones eslavas". Biología Molecular y Evolución . 22 (10): 1964–75. doi : 10.1093/molbev/msi185 . PMID 15944443.
^ M. Pericic; et al. (2005). "Revisión del patrimonio genético croata revelado por el ADN mitocondrial y los linajes cromosómicos Y". Revista médica croata . 46 (4): 502–513. PMID 16100752.
^ Pericić y col. 2005.
^ "Sin título". pereformat.ru (en ruso).
^ "Sin título". www.rodstvo.ru .
^ Zerjal y col. 2002.
^ Haber y col. 2012.
^ ab Di Cristofaro et al. 2013.
^ ab Malyarchuk et al. 2013.
^ Ashirbekov y col. 2017.
^ Sha 2011.
^ Arunkumar 2012.
^ Toomas Kivisild; Siiri Rootsi; Mait Metspalu; Ene Metspalu; Juri Parik; Katrin Kaldma; Esien Usanga; Sarabjit Mastana; Surinder S. Papiha; Richard Villems. "La genética del lenguaje y la agricultura se difundieron en la India" (PDF) . En P. Bellwwood; C. Renfrew (eds.). "Examinar la hipótesis de la dispersión agrícola y lingüística ". Monografías del Instituto McDonald. Universidad de Cambridge. págs. 215–222 . Consultado el 20 de diciembre de 2019 .
^ Fornarino y col. 2009.
^ Wang y otros. 2003.
^ Zhou y col. 2007.
^ Liu Shu-hu y otros. 2018.
^ Zhong y otros. 2011.
^ Zhong, Hua; Shi, Hong; Qi, Xue-Bin; Duan, Zi-Yuan; Bronceado, ping-ping; Jin, Li; Su, Bing; Ma, Runlin Z. (2011). "La investigación ampliada del cromosoma Y sugiere migraciones posglaciales de humanos modernos hacia el este de Asia a través de la ruta del norte". Biología Molecular y Evolución . 28 (1): 717–727. doi : 10.1093/molbev/msq247 . PMID 20837606.
^ Shou, Wei-Hua; Qiao, Wn-Fa; Wei, Chuan-Yu; Dong, Yong-Li; Tan, Si-Jie; Shi, Hong; Tang, Wen-Ru; Xiao, Chun-Jie (2010). "Las distribuciones del cromosoma Y entre las poblaciones del noroeste de China identifican una contribución significativa de los pastores de Asia central y una menor influencia de los euroasiáticos occidentales". Revista de genética humana . 55 (5): 314–322. doi : 10.1038/jhg.2010.30 . PMID 20414255. S2CID 23002493.
^ Lell y col. 2002.
^ Mohamed y otros. 2009.
^ Nasidze y col. 2004.
^ Nasidze y col. 2005.
^ Grugni y col. 2012.
Fuentes
Allentoft, Morten E.; Sikora, Martín; Sjögren, Karl-Göran; Rasmussen, Simón; Rasmussen, Morten; Stenderup, Jesper; Damgaard, Peter B.; Schroeder, Hannes; et al. (2015). "Genómica de poblaciones de Eurasia de la Edad del Bronce". Naturaleza . 522 (7555): 167–172. Código Bib :2015Natur.522..167A. doi : 10.1038/naturaleza14507. PMID 26062507. S2CID 4399103.
Anthony, David W. (2007), El caballo, la rueda y el lenguaje. Cómo los jinetes de la Edad del Bronce de las estepas euroasiáticas dieron forma al mundo moderno , Princeton University Press
Anthony, David (primavera-verano de 2019). "Arqueología, genética y lengua en las estepas: un comentario sobre Bomhard". Revista de estudios indoeuropeos . 47 (1–2) . Consultado el 9 de enero de 2020 .
Antonio, David; Ringe, Don (2015), "La patria indoeuropea desde perspectivas lingüísticas y arqueológicas", Annual Review of Linguistics , 1 : 199–219, doi : 10.1146/annurev-linguist-030514-124812
Shah, AM; Tamang, R.; Moorjani, P.; Rani, DS; Govindaraj, P.; Kulkarni, G.; Bhattacharya, T.; Mustak, MS; Bhaskar, LVKS; Reddy, AG; Gadhvi, D.; Gai, PB; Chaubey, G.; Patterson, N.; Reich, D.; Tyler-Smith, C.; Singh, L.; Thangaraj, K. (2011). "Siddis indios: descendientes africanos con mezcla india". La Revista Estadounidense de Genética Humana . 89 (1): 154–61. doi :10.1016/j.ajhg.2011.05.030. PMC 3135801 . PMID 21741027.
ArunKumar, G; Soria-Hernanz, DF; Kavitha, VJ; Arun, VS; Syama, A; Ashokan, KS (2012). "La diferenciación de la población de los linajes masculinos del sur de la India se correlaciona con las expansiones agrícolas anteriores al sistema de castas". MÁS UNO . 7 (11): e50269. Código Bib : 2012PLoSO...750269A. doi : 10.1371/journal.pone.0050269 . PMC 3508930 . PMID 23209694.
Ashirbekov, EE; et al. (2017). "Distribución de haplogrupos del cromosoma Y de los kazajos de las regiones del sur de Kazajstán, Zhambyl y Almaty" (PDF) . Informes de la Academia Nacional de Ciencias de la República de Kazajstán . 6 (316): 85–95. Archivado desde el original (PDF) el 23 de septiembre de 2021 . Consultado el 4 de junio de 2020 .
Balanovsky O, Rootsi S, Pshenichnov A, Kivisild T, Churnosov M, Evseeva I, Pocheshkhova E, Boldyreva M, et al. (2008). "Dos fuentes de la herencia patrilineal rusa en su contexto euroasiático". Revista Estadounidense de Genética Humana . 82 (1): 236–250. doi :10.1016/j.ajhg.2007.09.019. PMC 2253976 . PMID 18179905.
Балановский, О. P. (30 de noviembre de 2015). Генофонд Европы (en ruso). Prensa científica KMK. ISBN 9785990715707.
Battaglia V, Fornarino S, Al-Zahery N, Olivieri A, Pala M, Myres NM, King RJ, Rootsi S, et al. (2008). "Evidencia del cromosoma Y de la difusión cultural de la agricultura en el sudeste de Europa". Revista europea de genética humana . 17 (6): 820–30. doi :10.1038/ejhg.2008.249. PMC 2947100 . PMID 19107149.
Behar D, Thomas MG, Skorecki K, Hammer MF, Bulygina E, Rosengarten D, Jones AL, Held K, et al. (2003). "Múltiples orígenes de los levitas asquenazíes: evidencia del cromosoma Y para ascendencia europea y del Cercano Oriente" (PDF) . Revista Estadounidense de Genética Humana . 73 (4): 768–779. doi : 10.1086/378506 . PMC 1180600 . PMID 13680527.
Bowden GR, Balaresque P, King TE, Hansen Z, Lee AC, Pergl-Wilson G, Hurley E, Roberts SJ, et al. (2008). "Excavación de estructuras de población pasadas mediante muestreo basado en apellidos: el legado genético de los vikingos en el noroeste de Inglaterra". Biología Molecular y Evolución . 25 (2): 301–309. doi :10.1093/molbev/msm255. PMC 2628767 . PMID 18032405.
Brandit, G.; et al. (El Consorcio Genográfico) (2013). "El ADN antiguo revela etapas clave en la formación de la diversidad genética mitocondrial de Europa Central". Ciencia . 342 (6155): 257–261. Código Bib : 2013 Ciencia... 342..257B. doi : 10.1126/ciencia.1241844. PMC 4039305 . PMID 24115443.
Capelli C, Redhead N, Abernethy JK, Gratrix F, Wilson JF, Moen T, Hervig T, Richards M, et al. (2003). "Censo del cromosoma AY de las Islas Británicas" (PDF) . Biología actual . 13 (11): 979–84. doi :10.1016/S0960-9822(03)00373-7. PMID 12781138. S2CID 526263.también en el "University College London" (PDF) .
Chunxiang Li; Hongjie Li; Yinqiu Cui; Chengzhi Xie; Dawei Cai; Wenying Li; Víctor H Mair; Zhi Xu; et al. (2010). "Evidencia de que una población mixta de Oeste a Este vivía en la cuenca del Tarim ya en la Edad del Bronce". Biología BMC . 8 (1): 15. doi : 10.1186/1741-7007-8-15 . PMC 2838831 . PMID 20163704.
Di Cristofaro J, Pennarun E, Mazières S, Myres NM, Lin AA, Temori SA, Metspalu M, Metspalu E, et al. (2013). "Afghan Hindu Kush: donde convergen los flujos genéticos del subcontinente euroasiático". MÁS UNO . 8 (10). e76748. Código Bib : 2013PLoSO...876748D. doi : 10.1371/journal.pone.0076748 . PMC 3799995 . PMID 24204668.
Dupuy BM, Stenersen M, Lu TT, Olaisen B (2005). "Heterogeneidad geográfica de los linajes del cromosoma Y en Noruega" (PDF) . Internacional de Ciencias Forenses . 164 (1): 10-19. doi :10.1016/j.forsciint.2005.11.009. PMID 16337760.
Fornarino, Simona; Pala, María; Battaglia, Vincenza; Maranta, Ramona; Aquiles, Alejandro; Modiano, Guido; Torroní, Antonio; Semino, Ornella; et al. (2009). "Diversidad mitocondrial y del cromosoma Y de Tharus (Nepal): un reservorio de variación genética". Biología Evolutiva del BMC . 9 (1): 154. Código bibliográfico : 2009BMCEE...9..154F. doi : 10.1186/1471-2148-9-154 . PMC 2720951 . PMID 19573232.
Fu, Qiaomei; et al. (2 de mayo de 2016). "La historia genética de la Europa de la Edad del Hielo". Naturaleza . 534 (7606): 200–205. Código Bib :2016Natur.534..200F. doi : 10.1038/naturaleza17993. hdl :10211.3/198594. PMC 4943878 . PMID 27135931.
Grugni V, Battaglia V, Kashani BH, Parolo S, Al-Zahery N, Achilli A, Olivieri A, Gandini F, Houshmand M, Sanati MH, Torroni A, Semino O (2012). "Antiguos acontecimientos migratorios en el Medio Oriente: nuevas pistas de la variación del cromosoma Y de los iraníes modernos". MÁS UNO . 7 (7). e41252. Código Bib : 2012PLoSO...741252G. doi : 10.1371/journal.pone.0041252 . PMC 3399854 . PMID 22815981.
Gwozdz (2009). "Montañas Y-STR en el haploespacio, parte II: aplicación a clados polacos comunes" (PDF) . Revista de genealogía genética . 5 (2).
Haak, W.; Brandt, G.; Jong, HN d.; Meyer, C.; Ganslmeier, R.; Heyd, V.; Hawkesworth, C.; Lucio, AWG; et al. (2008). "El ADN antiguo, los isótopos de estroncio y los análisis osteológicos arrojan luz sobre la organización social y de parentesco de la Edad de Piedra posterior". Procedimientos de la Academia Nacional de Ciencias . 105 (47): 18226–18231. Código bibliográfico : 2008PNAS..10518226H. doi : 10.1073/pnas.0807592105 . PMC 2587582 . PMID 19015520.
Haak, Wolfgang; Lazaridis, José; Patterson, Nick; Rohland, Nadin; Mallick, Swapan; Llamas, Bastien; Brandt, Guido; Nordenfelt, Susanne; et al. (2015). "La migración masiva desde la estepa es una fuente de lenguas indoeuropeas en Europa". Naturaleza . 522 (7555): 207–211. arXiv : 1502.02783 . Código Bib :2015Natur.522..207H. bioRxiv 10.1101/013433 . doi :10.1038/NATURALEZA14317. PMC 5048219 . PMID 25731166.
Haber M, Platt DE, Ashrafian Bonab M, Youhanna SC, Soria-Hernanz DF, Martínez-Cruz B, Douaihy B, Ghassibe-Sabbagh M, et al. (2012). "Los grupos étnicos de Afganistán comparten una herencia cromosómica Y estructurada por acontecimientos históricos". MÁS UNO . 7 (3). e34288. Código Bib : 2012PLoSO...734288H. doi : 10.1371/journal.pone.0034288 . PMC 3314501 . PMID 22470552.
Karafet, Tatiana M.; Méndez, Fernando L.; Sudoyo, Herawati; Lansing, J. Stephen; Martillo, Michael F. (2014). "Resolución filogenética mejorada y rápida diversificación del haplogrupo K-M526 del cromosoma Y en el sudeste asiático". Naturaleza . 23 (3): 369–373. doi : 10.1038/ejhg.2014.106 . PMC 4326703 . PMID 24896152.
Kasperaviciūte, D.; Kucinskas, V.; Stoneking, M. (2005). "Variación del cromosoma Y y del ADN mitocondrial en lituanos". Anales de genética humana . 68 (5): 438–452. doi :10.1046/j.1529-8817.2003.00119.x. PMID 15469421. S2CID 26562505.
Kayser M, Lao O, Anslinger K, Augustin C, Bargel G, Edelmann J, Elias S, Heinrich M, et al. (2005). "La diferenciación genética significativa entre Polonia y Alemania sigue las fronteras políticas actuales, como lo revela el análisis del cromosoma Y" (PDF) . Genética Humana . 117 (5): 428–443. doi :10.1007/s00439-005-1333-9. PMID 15959808. S2CID 11066186. Archivado desde el original (PDF) el 4 de marzo de 2009.
Keyser, Christine; Bouakaze, Carolina; Crubézy, Eric; Nikolaev, Valery G.; Montagnon, Daniel; Reyes, Tatiana; Ludes, Bertrand (2009). "El ADN antiguo proporciona nuevos conocimientos sobre la historia del pueblo Kurgan del sur de Siberia". Genética Humana . 126 (3): 395–410. doi :10.1007/s00439-009-0683-0. PMID 19449030. S2CID 21347353.
Kim, Kijeong; Brenner, Charles H.; Mair, Víctor H.; Lee, Kwang-Ho; Kim, Jae-Hyun; Gelegdorj, Eregzen; Batbold, Natsag; Canción, Yi-Chung; et al. (2010). "Un hombre de Eurasia occidental se encuentra en el cementerio de élite Xiongnu de 2000 años de antigüedad en el noreste de Mongolia". Revista Estadounidense de Antropología Física . 142 (3): 429–440. doi :10.1002/ajpa.21242. PMID 20091844.
Kivisild, T; Rootsi, S; Metspalu, M; Mastana, S; Kaldma, K; Parik, J; Metspalu, E; Adojaan, M; et al. (2003). "La herencia genética de los primeros colonos persiste tanto en las poblaciones tribales como en las castas indias". AJHG . 72 (2): 313–32. doi :10.1086/346068. PMC 379225 . PMID 12536373.
Lazaridis, José; et al. (2016). "Conocimientos genómicos sobre el origen de la agricultura en el antiguo Cercano Oriente". Naturaleza . 536 (7617): 419–424. Código Bib :2016Natur.536..419L. doi : 10.1038/naturaleza19310. PMC 5003663 . PMID 27459054.
Lell JT, Sukernik RI, Starikovskaya YB, Su B, Jin L, Schurr TG, Underhill PA, Wallace DC (2002). "El origen dual y las afinidades siberianas de los cromosomas Y de los nativos americanos" (PDF) . Revista Estadounidense de Genética Humana . 70 (1): 192–206. doi :10.1086/338457. PMC 384887 . PMID 11731934. Archivado desde el original (PDF) el 22 de abril de 2003.
Liu Shu-hu; Nizam Yilihamu; Hornear Rabiyamu; Abdukeram Bupatima; Dolkun Matyusup (2018). "Un estudio de la diversidad genética de tres poblaciones aisladas en Xinjiang utilizando Y-SNP". Acta Antropológica Sínica . 37 (1): 146-156.
Carlos Quiles (10 de septiembre de 2018). "Un estudio de la diversidad genética de tres poblaciones aisladas en Xinjiang utilizando Y-SNP". Indoeuropeo .
Malmstrom, Helena; Günther, Torsten; Svensson, Emma M.; Juras, Anna; Fraser, Magdalena; Munters, Arielle R.; Pospieszny, Łukasz; Torv, Mari; et al. (9 de octubre de 2019). "La ascendencia genómica del pueblo escandinavo de la cultura del hacha de batalla y su relación con el horizonte más amplio de Corded Ware". Actas de la Royal Society B. 286 (1912). doi : 10.1098/rspb.2019.1528 . PMC 6790770 . PMID 31594508.
Malyarchuk, Boris; Derenko, Miroslava; Wozniak, Marcin; Grzybowski, Tomasz (2013). "Variación del cromosoma Y en tayikos e iraníes". Anales de biología humana . 40 (1): 48–54. doi :10.3109/03014460.2012.747628. PMID 23198991. S2CID 2752490.
Mascarenhas, Desmond D.; Raina, Anupuma; Aston, Christopher E.; Sanghera, Dharambir K. (2015). "Reconstrucción genética y cultural de la migración de un linaje antiguo". Investigación BioMed Internacional . 2015 : 651415. doi : 10.1155/2015/651415 . PMC 4605215 . PMID 26491681.
Mathieson, Iain; Lazaridis, José; Rohland, Nadin; Mallick, Swapan; Patterson, Nick; Alpaslan Roodenberg, Songul; Harney, Eadaoin; Stewardson, Kristin; et al. (2015). "Ocho mil años de selección natural en Europa". bioRxiv 10.1101/016477 .
Mirabal, Sheyla; Regueiro, M; Cadenas, AM; Cavalli-Sforza, LL; Underhill, Pensilvania; Verbenko, DA; Limborska, SA; Herrera, RJ; et al. (2009). "Distribución del cromosoma Y dentro del paisaje geolingüístico del noroeste de Rusia". Revista europea de genética humana . 17 (10): 1260-1273. doi :10.1038/ejhg.2009.6. PMC 2986641 . PMID 19259129.
Mohammad T, Xue Y, Evison M, Tyler-Smith C (2009). "Estructura genética de los beduinos nómadas de Kuwait". Herencia . 103 (5): 425–433. doi : 10.1038/hdy.2009.72 . PMC 2869035 . PMID 19639002.
Narasimhan, Vagheesh M.; Antonio, David; Mallory, James; Reich, David (2018). "La formación genómica de Asia central y meridional". bioRxiv 10.1101/292581 .
Nasidze I, Ling EY, Quinque D, Dupanloup I, Cordaux R, Rychkov S, Naumova O, Zhukova O, et al. (2004). "ADN mitocondrial y variación del cromosoma Y en el Cáucaso" (PDF) . Anales de genética humana . 68 (parte 3): 205–221. doi :10.1046/j.1529-8817.2004.00092.x. PMID 15180701. S2CID 27204150. Archivado desde el original (PDF) el 30 de octubre de 2004.
Nasidze I, Quinque D, Ozturk M, Bendukidze N, Stoneking M (2005). "Variación del ADNmt y del cromosoma Y en grupos kurdos" (PDF) . Anales de genética humana . 69 (parte 4): 401–412. doi :10.1046/j.1529-8817.2005.00174.x. PMID 15996169. S2CID 23771698. Archivado desde el original (PDF) el 23 de agosto de 2009.
Pamjav, Horolma; Feher, Tibor; Németh, Endre; Pádár, Zsolt (2012), "Breve comunicación: los nuevos marcadores binarios del cromosoma Y mejoran la resolución filogenética dentro del haplogrupo R1a1", American Journal of Physical Anthropology , 149 (4): 611–615, doi :10.1002/ajpa.22167, PMID 23115110 , S2CID 4820868
Passarino G, Cavalleri GL, Lin AA, Cavalli-Sforza LL, Børresen-Dale AL, Underhill (2002). "Diferentes componentes genéticos en la población noruega revelados por el análisis de polimorfismos del ADNmt y del cromosoma Y". Revista europea de genética humana . 10 (9): 521–529. doi : 10.1038/sj.ejhg.5200834 . PMID 12173029.
Pathak, Ajai K.; Kadiano, Anurag; Kushniarevich, Alena; Montinaro, Francesco; Mondal, Mayukh; Ongaró, Linda; Singh, Manvendra; Kumar, Pramod; et al. (6 de diciembre de 2018). "La ascendencia genética de las poblaciones modernas del valle del Indo del noroeste de la India". La Revista Estadounidense de Genética Humana . 103 (6): 918–929. doi :10.1016/j.ajhg.2018.10.022. PMC 6288199 . PMID 30526867.
Pawlowski, R; Dettlaff-Kakol, A; MacIejewska, A; Paszkowska, R; Reichert, M; Jezierski, G (2002). "Genética de poblaciones de loci STR del cromosoma 9 Y en el norte de Polonia". Arco. Medicina. Sadowej Kryminol . 52 (4): 261–277. PMID 14669672.
Pericić M, Lauc LB, Klarić IM, Rootsi S, Janićijević B, Rudan I, Terzić R, Colak I, et al. (2005). "El análisis filogenético de alta resolución del sureste de Europa rastrea episodios importantes de flujo de genes paternos entre las poblaciones eslavas". Mol. Biol. Evolución . 22 (10): 1964–75. doi : 10.1093/molbev/msi185 . PMID 15944443.
Poznik GD, et al. (2016). "Estallidos puntuados en la demografía masculina humana inferidos de 1244 secuencias del cromosoma Y en todo el mundo". Genética de la Naturaleza . 48 (6): 593–599. doi :10.1038/ng.3559. hdl :11858/00-001M-0000-002A-F024-C. PMC 4884158 . PMID 27111036.
Regueiro, M; Cadenas, AM; Gayden, T; Underhill, Pensilvania; Herrera, RJ (2006). "Irán: nexo tricontinental para la migración impulsada por el cromosoma Y". Hum Hered . 61 (3): 132-143. doi : 10.1159/000093774. PMID 16770078. S2CID 7017701.
Ricaut F, Keyser-Tracqui C, Bourgeois I, Crubézy E, Ludes B (2004). "Análisis genético de un esqueleto escito-siberiano y sus implicaciones para las antiguas migraciones de Asia Central". Biología humana . 76 (1): 109–25. doi :10.1353/hub.2004.0025. PMID 15222683. S2CID 35948291.
Rosser ZH, Zerjal T, Hurles ME, Adojaan M, Alavantic D, Amorim A, Amos W, Armenteros M, et al. (2000). "La diversidad del cromosoma Y en Europa es clinal y está influenciada principalmente por la geografía, más que por el idioma". Revista Estadounidense de Genética Humana . 67 (6): 1526-1543. doi : 10.1086/316890 . PMC 1287948 . PMID 11078479.
Saag, Lehti; Varul, Liivi; Scheib, Christiana Lyn; Stenderup, Jesper; Allentoft, Morten E.; Saag, Lauri; Pagani, Luca; Reidla, Maere; et al. (24 de julio de 2017). "La agricultura extensiva en Estonia comenzó a través de una migración desde la estepa con sesgo sexual". Biología actual . Prensa celular . 27 (14): 2185–2193. doi : 10.1016/j.cub.2017.06.022 . PMID 28712569.
Saag, Lehti; Vasiliev, Sergey V.; Varul, Liivi; Kosorukova, Natalia V.; Gerasimov, Dmitri V.; Oshibkina, Svetlana V.; Griffith, Samuel J.; Solnik, Anu; et al. (3 de julio de 2020). "Cambios de ascendencia genética en la transición de la Edad de Piedra a la Edad del Bronce en la llanura de Europa del Este". bioRxiv 10.1101/2020.07.02.184507 .
Sahoo, S; Singh, A; Himabindu, G; Banerjee, J; Sitalaximi, T; Gaikwad, S; Trivedi, R; Endicott, P; et al. (2006). "Una prehistoria de los cromosomas Y indios: evaluación de escenarios de difusión démica". Procedimientos de la Academia Nacional de Ciencias . 103 (4): 843–848. Código Bib : 2006PNAS..103..843S. doi : 10.1073/pnas.0507714103 . PMC 1347984 . PMID 16415161.
Scozzari R, Cruciani F, Pangrazio A, Santolamazza P, Vona G, Moral P, Latini V, Varesi L, et al. (2001). "Variación del cromosoma Y humano en el área del Mediterráneo occidental: implicaciones para el poblamiento de la región" (PDF) . Inmunología humana . 62 (9): 871–84. CiteSeerX 10.1.1.408.4857 . doi :10.1016/S0198-8859(01)00286-5. PMID 11543889.
Semenov, Alexander S.; Bulat, Vladimir V. (2016), "Paleo-ADN antiguo de Europa nororiental anterior a la era del cobre: establecimiento de las huellas de migración del haplogrupo de ADN-Y R1a1", Revista europea de biotecnología molecular , 11 (1): 40–54 , doi :10.13187/ejmb.2016.11.40, S2CID 172131289
Semino, O; Passarino, G; Oefner, PJ; Lin, AA; Arbuzova, S; Beckman, LE; De Benedictis, G; Francalacci, P; et al. (2000). "El legado genético del Homo sapiens sapiens paleolítico en los europeos existentes: perspectiva del cromosoma AY" (PDF) . Ciencia . 290 (5494): 1155-1159. Código Bib : 2000 Ciencia... 290.1155S. doi : 10.1126/ciencia.290.5494.1155. PMID 11073453. Archivado desde el original (PDF) el 25 de noviembre de 2003.
Sengupta, S; Zhivotovsky, LA; Rey, R; Mehdi, SQ; Edmonds, California; Chow, CE; Lin, AA; Mitra, M; et al. (2006). "La polaridad y la temporalidad de las distribuciones del cromosoma Y de alta resolución en la India identifican expansiones tanto indígenas como exógenas y revelan una influencia genética menor de los pastores de Asia central". Revista Estadounidense de Genética Humana . 78 (2): 202–21. doi :10.1086/499411. PMC 1380230 . PMID 16400607.
Sharma, S; Rai, E; Sharma, P; Jena, M; Singh, S; Darvishi, K; Bhat, Alaska; Bhanwer, AJ; et al. (2009). "El origen indio del haplogrupo paterno R1a1 (*) corrobora el origen autóctono de los brahmanes y el sistema de castas". Revista de genética humana . 54 (1): 47–55. doi : 10.1038/jhg.2008.2 . PMID 19158816.
Schilz, Félix (2006). Molekulargenetische Verwandtschaftsanalysen am prähistorischen Skelettkollektiv der Lichtensteinhöhle [ Análisis de parentesco genético molecular sobre el colectivo de esqueletos prehistóricos de la cueva de Lichtenstein ] (PDF) (Disertación) (en alemán). Gotinga: Mathematisch-Naturwissenschaftlichen Fakultäten der Georg-August-Universität.
Silva, Marina; et al. (2017). "Una cronología genética para el subcontinente indio apunta a dispersiones fuertemente sesgadas por el sexo". Biología Evolutiva del BMC . 17 (1): 88. Código bibliográfico : 2017BMCEE..17...88S. doi : 10.1186/s12862-017-0936-9 . PMC 5364613 . PMID 28335724.
Tambets K, Rootsi S, Kivisild T, Help H, Serk P, Loogväli EL, Tolk HV, Reidla M, et al. (2004). "Las raíces occidentales y orientales de los saami: la historia de los 'valores atípicos' genéticos contados por el ADN mitocondrial y los cromosomas Y". Revista Estadounidense de Genética Humana . 74 (4): 661–682. doi : 10.1086/383203 . PMC 1181943 . PMID 15024688.
Thangaraj, Kumarasamy; Naidu, B. Prathap; Crivellaro, Federica; Tamang, Rakesh; Upadhyay, Shashank; Sharma, Varun Kumar; Reddy, Alla G.; Walimbe, SR; et al. (2010). Cordaux, Richard (ed.). "La influencia de las barreras naturales en la configuración de la estructura genética de las poblaciones de Maharashtra". MÁS UNO . 5 (12): e15283. Código Bib : 2010PLoSO...515283T. doi : 10.1371/journal.pone.0015283 . PMC 3004917 . PMID 21187967.
Underhill, Pensilvania; Myres, Nuevo México; Rootsi, S; Metspalu, M; Zhivotovsky, LA; Rey, RJ; Lin, AA; Chow, CE; et al. (4 de noviembre de 2009). "Separación de la coascendencia posglacial de los cromosomas Y europeos y asiáticos dentro del haplogrupo R1a". Revista europea de genética humana (publicada en abril de 2010). 18 (4): 479–84. doi : 10.1038/ejhg.2009.194 . PMC 2987245 . PMID 19888303.
Underhill, Peter A.; et al. (26 de marzo de 2014). "La estructura filogenética y geográfica del haplogrupo R1a del cromosoma Y". Revista europea de genética humana (publicada en enero de 2015). 23 (1): 124-131. doi : 10.1038/ejhg.2014.50 . PMC 4266736 . PMID 24667786. "PDF" (PDF) . Archivado desde el original (PDF) el 16 de agosto de 2016 . Consultado el 12 de junio de 2016 .
Wang, Wei; Sabia, Cheryl; Baric, Tom; Negro, Michael L.; Bittles, Alan H. (2003). "Los orígenes y la estructura genética de tres poblaciones musulmanas chinas co-residentes: los Salar, Bo'an y Dongxiang". Genética Humana . 113 (3): 244–52. doi :10.1007/s00439-003-0948-y. PMID 12759817. S2CID 11138499.
Wells, RS (2001), "El corazón de Eurasia: una perspectiva continental sobre la diversidad del cromosoma Y", Actas de la Academia Nacional de Ciencias de EE. UU. , 98 (18): 10244–10249, Bibcode :2001PNAS...9810244W, doi : 10.1073/pnas.171305098 , PMC 56946 , PMID 11526236
Yan, Shi; Wang, Chuan-Chao; Zheng, Hong-Xiang; Wang, Wei; Qin, Zhen-Dong; Wei, Lan-Hai; Wang, Yi; Pan, Xue-Dong; et al. (29 de agosto de 2014). "Los cromosomas Y del 40% de los chinos descienden de tres superabuelos neolíticos". MÁS UNO . 9 (8). e105691. arXiv : 1310.3897 . Código Bib : 2014PLoSO...9j5691Y. doi : 10.1371/journal.pone.0105691 . PMC 4149484 . PMID 25170956.
Consorcio del Cromosoma Y "YCC" (2002). "Un sistema de nomenclatura para el árbol de haplogrupos binarios del cromosoma Y humano". Investigación del genoma . 12 (2): 339–348. doi :10.1101/gr.217602. PMC 155271 . PMID 11827954.
Zerjal, Tatiana; Wells, R. Spencer; Yuldasheva, Nadira; Ruzibakiev, Ruslan; Tyler-Smith, Chris (2002). "Un paisaje genético remodelado por acontecimientos recientes: conocimientos del cromosoma Y en Asia central". La Revista Estadounidense de Genética Humana . 71 (3): 466–82. doi :10.1086/342096. PMC 419996 . PMID 12145751.
Zhong H, Shi H, Qi XB, Duan Y, Tan PP, Jin L, SU B, Ma RZ (enero de 2011). "La investigación ampliada del cromosoma Y sugiere migraciones posglaciares de humanos modernos hacia el este de Asia a través de la ruta del norte". Biología Molecular y Evolución . 28 (1): 717–27. doi : 10.1093/molbev/msq247 . PMID 20837606.
Zhou, Ruixia; An, Lizhe; Wang, Xunling; Shao, Wei; Lin, Gonghua; Yu, Weiping; Yi, Lin; Xu, Shijian; et al. (2007). "Prueba de la hipótesis del origen de un antiguo soldado romano del pueblo Liqian en el noroeste de China: una perspectiva del cromosoma Y". Revista de genética humana . 52 (7): 584–91. doi : 10.1007/s10038-007-0155-0 . PMID 17579807.
Otras lecturas
Gimbutas (1970). Indoeuropeos e indoeuropeos . Filadelfia, PA: Univ. de Prensa de Pensilvania. págs. 155-195.
Patrizia Malaspina; Andréi I. Kozlov; Fulvio Cruciani; Piero Santolamazza; Nejat Akar; Dimiter Kovatchev; Marina G. Kerimova; Juri Parik; Richard Villems; Rosana Scozzari; Andrea Novelletto (2003). "Análisis de la variación del cromosoma Y en poblaciones modernas en la frontera entre Europa y Asia" (PDF) . En K. Boyle; C. Renfrew; M. Levine (eds.). Interacciones antiguas: este y oeste en Eurasia . Monografías del Instituto McDonald. Cambridge: Prensa de la Universidad de Cambridge. págs. 309–313.
Parpola, Asko (2015). Las raíces del hinduismo. Los primeros arios y la civilización del Indo . Prensa de la Universidad de Oxford.
Sharma, S. (2007). "1344/T: El origen autóctono y un vínculo tribal de los brahmanes indios: evaluación a través de marcadores genéticos moleculares". 57ª reunión anual de la Sociedad Estadounidense de Genética Humana, 23 al 27 de octubre de 2007; San Diego, California (PDF) (Resumen). pag. 273. Archivado desde el original (PDF) el 26 de junio de 2008.
Trautmann, Thomas (2005). El debate ario . Prensa de la Universidad de Oxford.
Varzari, Alejandro (2006). Historia de la población de los Dniéster-Cárpatos: evidencia de la inserción de Alu y polimorfismos del cromosoma Y (PDF) (Disertación). Múnich: Universidad Ludwig-Maximilians.
Wells, Spencer (2002). El viaje del hombre: una odisea genética . Prensa de la Universidad de Princeton. ISBN 978-0-691-11532-0.
enlaces externos
Wikimedia Commons tiene medios relacionados con el haplogrupo R1a del ADN-Y .
Árbol de ADN
Proyecto de haplogrupo del cromosoma Y FTDNA R1a
Proyecto R1a1a1 y Subclades Y-DNA - Antecedentes del árbol genealógico ADN R1a1a1
TMRCA
TMRCA = Tiempo hasta el ancestro común más reciente
Varios
Proyecto de ADN regional de Demes danés: Haplogrupo R1a de ADN-Y Archivado el 20 de agosto de 2008 en Wayback Machine.
Blog de Eurogenes, El caso atípico de Poltovka
Avotaynu Online, La huella digital del ADN-Y de Shpoler Zeida, un tzadik que conmovió al mundo

_and_R1a1a_oldest_expansion_and_highest_frequency_(2014).jpg/1280px-R1a_origins_(Underhill_2010)_and_R1a1a_oldest_expansion_and_highest_frequency_(2014).jpg)