La ecolocalización , también llamada biosónar , es un sonar biológico activo utilizado por varios grupos de animales , tanto en el aire como bajo el agua. Los animales ecolocalizadores emiten llamadas y escuchan los ecos de esas llamadas que regresan de varios objetos cercanos a ellos. Utilizan estos ecos para localizar e identificar los objetos. La ecolocalización se utiliza para la navegación , la búsqueda de alimento y la caza de presas .
Las llamadas de ecolocalización pueden ser de frecuencia modulada (FM, que varía de tono durante la llamada) o de frecuencia constante (CF). La FM ofrece una discriminación precisa del rango para localizar la presa, a costa de un rango operativo reducido. La CF permite detectar tanto la velocidad de la presa como sus movimientos mediante el efecto Doppler . La FM puede ser mejor para entornos cerrados y desordenados, mientras que la CF puede ser mejor en entornos abiertos o para cazar mientras se está posado.
Los animales que utilizan la ecolocalización incluyen a los mamíferos, especialmente a los odontocetos (ballenas dentadas) y algunas especies de murciélagos , y, utilizando formas más simples, a especies de otros grupos como las musarañas . Unas pocas especies de aves de dos grupos de aves que habitan en cuevas utilizan la ecolocalización, a saber, las salanganas cavernícolas y el guácharo .
Algunas presas que cazan murciélagos ecolocalizadores toman contramedidas activas para evitar ser capturadas. Estas incluyen evitar a los depredadores, desviar los ataques y usar chasquidos ultrasónicos , que han desarrollado múltiples funciones, entre ellas el aposematismo, el mimetismo de especies que se defienden químicamente y el bloqueo de la ecolocalización.
El término ecolocalización fue acuñado en 1944 por el zoólogo estadounidense Donald Griffin , quien, junto con Robert Galambos , demostró por primera vez el fenómeno en murciélagos. [1] [2] Como describió Griffin en su libro, [3] el científico italiano del siglo XVIII Lazzaro Spallanzani había concluido, mediante una serie de experimentos elaborados, que cuando los murciélagos vuelan de noche, dependen de algún sentido además de la vista, pero no descubrió que el otro sentido fuera el oído. [4] [5] El médico y naturalista suizo Louis Jurine repitió los experimentos de Spallanzani (usando diferentes especies de murciélagos) y concluyó que cuando los murciélagos cazan de noche, dependen del oído. [6] [7] [8] En 1908, Walter Louis Hahn confirmó los hallazgos de Spallanzani y Jurine. [9]
En 1912, el inventor Hiram Maxim propuso de forma independiente que los murciélagos utilizaban sonidos por debajo del rango auditivo humano para evitar obstáculos. [10] En 1920, el fisiólogo inglés Hamilton Hartridge propuso correctamente en cambio que los murciélagos utilizaban frecuencias por encima del rango de la audición humana . [11] [12]
La ecolocalización en los odontocetos (ballenas dentadas) no fue descrita apropiadamente hasta dos décadas después del trabajo de Griffin y Galambos, por Schevill y McBride en 1956. [13] Sin embargo, en 1953, Jacques Yves Cousteau sugirió en su primer libro, El mundo silencioso , que las marsopas tenían algo así como un sonar , a juzgar por sus habilidades de navegación. [14]
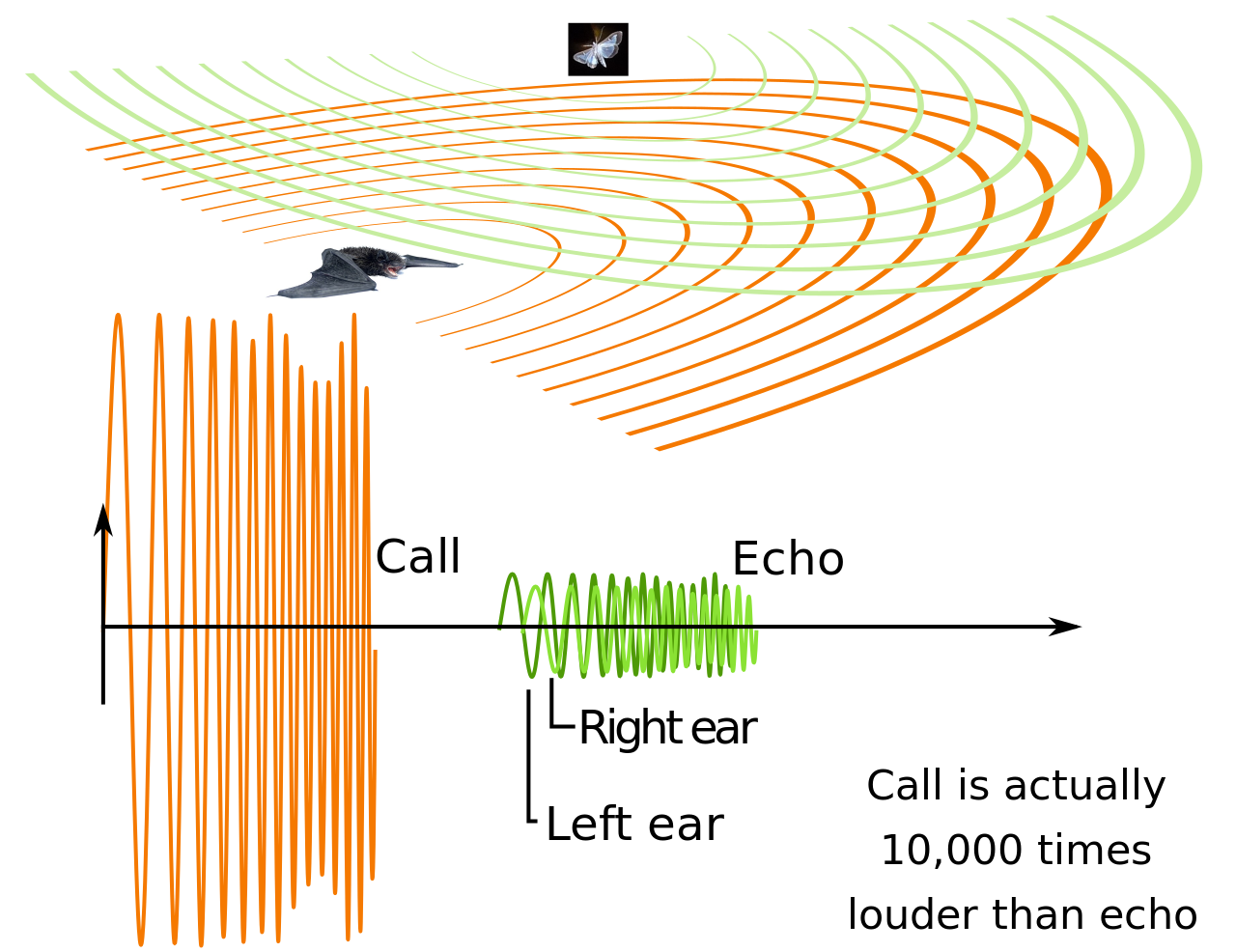
La ecolocalización es un sonar activo que utiliza sonidos emitidos por el propio animal. La determinación de la distancia se logra midiendo el tiempo de retardo entre la emisión de sonido del propio animal y los ecos que regresan del entorno. La intensidad relativa del sonido recibido en cada oído, así como el tiempo de retardo entre la llegada a los dos oídos, proporcionan información sobre el ángulo horizontal (acimut) desde el que llegan las ondas sonoras reflejadas. [15]
A diferencia de algunos sonares fabricados por el hombre que dependen de muchos haces extremadamente estrechos y muchos receptores para localizar un objetivo ( sonares multihaz ), la ecolocalización animal tiene solo un transmisor y dos receptores (las orejas) colocados ligeramente separados. Los ecos que regresan a las orejas llegan en diferentes momentos y con diferentes intensidades, dependiendo de la posición del objeto que genera los ecos. Los animales utilizan las diferencias de tiempo y volumen para percibir la distancia y la dirección. Con la ecolocalización, el murciélago u otro animal puede saber, no solo hacia dónde se dirige, sino también qué tan grande es otro animal, qué tipo de animal es y otras características. [16] [17]
Para describir la diversidad de los llamados de ecolocalización es necesario examinar la frecuencia y las características temporales de los llamados. Son las variaciones en estos aspectos las que producen llamados de ecolocalización adecuados para diferentes entornos acústicos y comportamientos de caza. Los llamados de los murciélagos han sido los más estudiados, pero los principios se aplican a todos los llamados de ecolocalización. [18] [19]
Las frecuencias de llamada de los murciélagos varían desde tan solo 11 kHz hasta tan altas como 212 kHz. [20] Los murciélagos insectívoros de caza aérea, aquellos que persiguen presas al aire libre, tienen una frecuencia de llamada entre 20 kHz y 60 kHz, porque es la frecuencia que brinda el mejor alcance y agudeza de imagen y los hace menos visibles para los insectos. [21] Sin embargo, las frecuencias bajas son adaptativas para algunas especies con presas y entornos diferentes. Euderma maculatum , una especie de murciélago que se alimenta de polillas , utiliza una frecuencia particularmente baja de 12,7 kHz que las polillas no pueden escuchar. [22]
Los llamados de ecolocalización pueden estar compuestos por dos tipos diferentes de estructura de frecuencia: barridos de frecuencia modulada (FM) y tonos de frecuencia constante (CF). Un llamado en particular puede constar de una, la otra o ambas estructuras. Un barrido de FM es una señal de banda ancha, es decir, contiene un barrido descendente a través de un rango de frecuencias. Un tono CF es una señal de banda estrecha: el sonido se mantiene constante en una frecuencia durante toda su duración. [23]
Se han medido llamadas de ecolocalización en murciélagos a intensidades que van desde los 60 a los 140 decibeles . [24] Algunas especies de murciélagos pueden modificar la intensidad de su llamada a mitad de la llamada, disminuyendo la intensidad a medida que se acercan a objetos que reflejan el sonido con fuerza. Esto evita que el eco de retorno ensordezca al murciélago. [19] Las llamadas de alta intensidad, como las de los murciélagos que cazan desde el aire (133 dB), son adaptativas a la caza en cielos abiertos. Sus llamadas de alta intensidad son necesarias incluso para tener una detección moderada de los alrededores porque el aire tiene una alta absorción de ultrasonidos y porque el tamaño de los insectos solo proporciona un objetivo pequeño para la reflexión del sonido. [25] Además, los llamados "murciélagos susurrantes" han adaptado la ecolocalización de baja amplitud para que sus presas, las polillas, que pueden escuchar llamadas de ecolocalización, sean menos capaces de detectar y evitar un murciélago que se aproxima. [22] [26]
Una sola llamada de ecolocalización (una llamada que es un solo rastro continuo en un espectrograma de sonido , y una serie de llamadas que comprenden una secuencia o pase) puede durar desde menos de 3 a más de 50 milisegundos de duración. La duración del pulso es de alrededor de 3 milisegundos en murciélagos FM como Phyllostomidae y algunos Vespertilionidae; entre 7 y 16 milisegundos en murciélagos de frecuencia casi constante (QCF) como otros Vespertilionidae, Emballonuridae y Molossidae; y entre 11 milisegundos (Hipposideridae) y 52 milisegundos (Rhinolophidae) en murciélagos CF. [27] La duración también depende de la etapa del comportamiento de captura de presas en la que se encuentre el murciélago, generalmente disminuye cuando el murciélago está en las etapas finales de la captura de presas; esto le permite al murciélago llamar más rápidamente sin superposición de llamada y eco. La reducción de la duración se produce a costa de tener menos sonido total disponible para reflejarse en los objetos y ser escuchado por el murciélago. [20]
El intervalo de tiempo entre llamadas de ecolocalización posteriores (o pulsos) determina dos aspectos de la percepción de un murciélago. En primer lugar, establece la rapidez con la que se actualiza la información auditiva del murciélago sobre la escena. Por ejemplo, los murciélagos aumentan la frecuencia de repetición de sus llamadas (es decir, disminuyen el intervalo de pulso) a medida que se acercan a un objetivo. Esto permite al murciélago obtener nueva información sobre la ubicación del objetivo a un ritmo más rápido cuando más la necesita. En segundo lugar, el intervalo de pulso determina el rango máximo en el que los murciélagos pueden detectar objetos. Esto se debe a que los murciélagos solo pueden seguir el rastro de los ecos de una llamada a la vez; tan pronto como hacen otra llamada, dejan de escuchar los ecos de la llamada realizada anteriormente. Por ejemplo, un intervalo de pulso de 100 ms (típico de un murciélago que busca insectos) permite que el sonido viaje en el aire aproximadamente 34 metros, por lo que un murciélago solo puede detectar objetos a una distancia de hasta 17 metros (el sonido tiene que viajar de ida y vuelta). Con un intervalo de pulso de 5 ms (típico de un murciélago en los momentos finales de un intento de captura), el murciélago solo puede detectar objetos a una distancia de hasta 85 cm. Por lo tanto, el murciélago tiene que elegir constantemente entre obtener nueva información actualizada rápidamente o detectar objetos lejanos. [28]
La principal ventaja que ofrece una señal FM es la discriminación de rango extremadamente precisa, o localización , del objetivo. JA Simmons demostró este efecto con una serie de experimentos que mostraban cómo los murciélagos que usaban señales FM podían distinguir entre dos objetivos separados incluso cuando los objetivos estaban separados por menos de medio milímetro. Esta capacidad se debe al barrido de banda ancha de la señal, que permite una mejor resolución del retardo de tiempo entre el llamado y el eco de retorno, mejorando así la correlación cruzada de los dos. Si se agregan frecuencias armónicas a la señal FM, entonces esta localización se vuelve aún más precisa. [29] [30] [31]
Una posible desventaja de la señal FM es la disminución del alcance operativo del llamado. Debido a que la energía del llamado se distribuye entre muchas frecuencias, la distancia a la que el murciélago FM puede detectar objetivos es limitada. [32] Esto se debe en parte a que cualquier eco que regrese a una frecuencia particular solo puede evaluarse durante una breve fracción de milisegundo, ya que el rápido barrido descendente del llamado no permanece en ninguna frecuencia durante mucho tiempo. [30]
La estructura de una señal CF es adaptativa, ya que permite al murciélago CF detectar tanto la velocidad de un objetivo como el aleteo de las alas de un objetivo como frecuencias desplazadas por el efecto Doppler. Un desplazamiento Doppler es una alteración en la frecuencia de la onda sonora y se produce en dos situaciones relevantes: cuando el murciélago y su objetivo se mueven uno con respecto al otro y cuando las alas del objetivo oscilan de un lado a otro. Los murciélagos CF deben compensar los desplazamientos Doppler, reduciendo la frecuencia de su llamada en respuesta a los ecos de frecuencia elevada; esto garantiza que el eco de retorno permanezca en la frecuencia a la que los oídos del murciélago están más finamente sintonizados. La oscilación de las alas de un objetivo también produce cambios de amplitud, lo que proporciona al murciélago CF una ayuda adicional para distinguir un objetivo volador de uno estacionario. [33] [29] Los murciélagos de herradura cazan de esta manera. [34]
Además, debido a que la energía de la señal de una llamada CF se concentra en una banda de frecuencia estrecha, el rango operativo de la llamada es mucho mayor que el de una señal FM. Esto se basa en el hecho de que los ecos que regresan dentro de la banda de frecuencia estrecha se pueden sumar a lo largo de toda la duración de la llamada, que mantiene una frecuencia constante durante hasta 100 milisegundos. [30] [32]
Un componente FM es excelente para cazar presas mientras se vuela en entornos cerrados y con mucho ruido. Dos aspectos de la señal FM explican este hecho: la localización precisa del objetivo conferida por la señal de banda ancha y la corta duración del llamado. El primero de estos es esencial porque en un entorno con mucho ruido, los murciélagos deben ser capaces de distinguir a sus presas de grandes cantidades de ruido de fondo. Las capacidades de localización 3D de la señal de banda ancha permiten al murciélago hacer exactamente eso, proporcionándole lo que Simmons y Stein (1980) llaman una "estrategia de rechazo del ruido". [31] Esta estrategia se mejora aún más mediante el uso de armónicos, que, como se dijo anteriormente, mejoran las propiedades de localización del llamado. La corta duración del llamado FM también es mejor en entornos cerrados y con mucho ruido porque permite al murciélago emitir muchos llamados extremadamente rápido sin superposición. Esto significa que el murciélago puede obtener un flujo casi continuo de información -esencial cuando los objetos están cerca, porque pasarán rápidamente- sin confundir qué eco corresponde a qué llamado. [33] [29]
Los murciélagos que buscan presas en vuelo en entornos abiertos y sin obstáculos suelen utilizar un componente CF, o que esperan posados a que aparezca su presa. El éxito de la primera estrategia se debe a dos aspectos del llamado CF, que confieren una excelente capacidad de detección de presas. En primer lugar, el mayor alcance del llamado permite a los murciélagos detectar objetivos presentes a grandes distancias, una situación común en entornos abiertos. En segundo lugar, la duración del llamado también es adecuada para objetivos a grandes distancias: en este caso, hay una menor probabilidad de que el llamado largo se superponga con el eco de retorno. La última estrategia es posible gracias a que el llamado largo y de banda estrecha permite al murciélago detectar los cambios Doppler, que se producirían si un insecto se moviera hacia o desde un murciélago posado. [33] [31] [29]
La ecolocalización se produce en una variedad de mamíferos y aves como se describe a continuación. [35] Evolucionó repetidamente, un ejemplo de evolución convergente . [29] [36]

Los murciélagos ecolocalizadores utilizan la ecolocalización para orientarse y buscar alimento, a menudo en total oscuridad. Por lo general, emergen de sus refugios en cuevas, áticos o árboles al anochecer y cazan insectos durante la noche. Mediante la ecolocalización, los murciélagos pueden determinar a qué distancia se encuentra un objeto, su tamaño, forma y densidad, y la dirección (si la hay) en la que se mueve. Su uso de la ecolocalización, junto con el vuelo propulsado, les permite ocupar un nicho en el que a menudo hay muchos insectos (que salen de noche, ya que hay menos depredadores), menos competencia por el alimento y menos especies que puedan cazar a los propios murciélagos. [37]
Los murciélagos ecolocalizadores generan ultrasonidos a través de la laringe y emiten el sonido a través de la boca abierta o, mucho más raramente, la nariz. [38] Esto último es más pronunciado en los murciélagos de herradura ( Rhinolophus spp. ). Las llamadas de ecolocalización de los murciélagos varían en frecuencia de 14.000 a más de 100.000 Hz, en su mayoría fuera del rango del oído humano (se considera que el rango de audición humana típico es de 20 Hz a 20.000 Hz). Los murciélagos pueden estimar la elevación de los objetivos interpretando los patrones de interferencia causados por los ecos que se reflejan desde el trago , un colgajo de piel en el oído externo. [39]
Las especies de murciélagos individuales se ecolocalizan dentro de rangos de frecuencia específicos que se adaptan a su entorno y tipos de presas. En ocasiones, los investigadores han utilizado esta técnica para identificar a los murciélagos que vuelan en una zona simplemente grabando sus llamadas con grabadoras ultrasónicas conocidas como "detectores de murciélagos". Sin embargo, las llamadas de ecolocalización no siempre son específicas de cada especie y algunos murciélagos se superponen en el tipo de llamadas que utilizan, por lo que las grabaciones de llamadas de ecolocalización no se pueden utilizar para identificar a todos los murciélagos. Los investigadores de varios países han desarrollado "bibliotecas de llamadas de murciélagos" que contienen grabaciones de "llamadas de referencia" de especies de murciélagos locales para ayudar con la identificación. [40] [41] [42]
Cuando buscan una presa, producen sonidos a una frecuencia baja (entre 10 y 20 clics por segundo). Durante la fase de búsqueda, la emisión de sonido se acopla a la respiración, que a su vez se acopla al aleteo. Este acoplamiento parece ahorrar energía de forma drástica, ya que el coste energético adicional de la ecolocalización para los murciélagos en vuelo es mínimo o nulo. [43] Después de detectar una presa potencial, los murciélagos ecolocalizadores aumentan la frecuencia de los pulsos, que terminan con el zumbido terminal , a frecuencias de hasta 200 clics por segundo. Durante la aproximación a un objetivo detectado, la duración de los sonidos disminuye gradualmente, al igual que la energía del sonido. [44]
Los murciélagos evolucionaron a principios del Eoceno , hace unos 64 millones de años . Los Yangochiroptera aparecieron hace unos 55 millones de años y los Rhinolophoidea hace unos 52 millones de años. [45]
Existen dos hipótesis sobre la evolución de la ecolocalización en los murciélagos. La primera sugiere que la ecolocalización laríngea evolucionó dos veces o más en los quirópteros, al menos una vez en los yangochiroptera y al menos una vez en los murciélagos de herradura (Rhinolophidae): [46]
La segunda propone que la ecolocalización laríngea tuvo un único origen en Chiroptera, es decir, que era basal para el grupo, y posteriormente se perdió en la familia Pteropodidae . [47] Más tarde, el género Rousettus en la familia Pteropodidae desarrolló un mecanismo diferente de ecolocalización utilizando un sistema de chasquidos con la lengua: [48]
Los murciélagos ecolocalizadores ocupan un conjunto diverso de condiciones ecológicas; se los puede encontrar viviendo en entornos tan diferentes como Europa y Madagascar , y cazando fuentes de alimento tan diferentes como insectos, ranas, néctar, fruta y sangre. Las características de un llamado de ecolocalización están adaptadas al entorno particular, al comportamiento de caza y a la fuente de alimento del murciélago en particular. La adaptación de los llamados de ecolocalización a los factores ecológicos está limitada por la relación filogenética de los murciélagos, lo que conduce a un proceso conocido como descendencia con modificación, y que da como resultado la diversidad de los quirópteros actuales. [29] [32] [31] Los murciélagos pueden interferir entre sí sin darse cuenta y, en algunas situaciones, pueden dejar de llamar para evitar interferir. [49]
Los insectos voladores son una fuente común de alimento para los murciélagos que utilizan la ecolocalización y algunos insectos (en particular las polillas) pueden oír los llamados de los murciélagos depredadores. Sin embargo, la evolución de los órganos auditivos en las polillas es anterior al origen de los murciélagos, por lo que, si bien muchas polillas escuchan la ecolocalización de los murciélagos que se acercan, sus oídos no evolucionaron originalmente en respuesta a las presiones selectivas de los murciélagos. [50] Estas adaptaciones de las polillas ejercen una presión selectiva sobre los murciélagos para mejorar sus sistemas de caza de insectos y este ciclo culmina en una " carrera armamentista evolutiva " entre polillas y murciélagos . [51] [52]
Debido a que los murciélagos utilizan la ecolocalización para orientarse y localizar objetos, sus sistemas auditivos están adaptados para este propósito, altamente especializados para detectar e interpretar los llamados de ecolocalización estereotipados característicos de su propia especie. Esta especialización es evidente desde el oído interno hasta los niveles más altos de procesamiento de información en la corteza auditiva. [53]
Tanto los murciélagos CF como los FM tienen oídos internos especializados que les permiten escuchar sonidos en el rango ultrasónico, muy fuera del rango de audición humana. Aunque en la mayoría de los otros aspectos, los órganos auditivos del murciélago son similares a los de la mayoría de los demás mamíferos, ciertos murciélagos ( murciélagos de herradura , Rhinolophus spp. y el murciélago bigotudo , Pteronotus parnelii ) con un componente de frecuencia constante (CF) en su llamado (conocidos como murciélagos de ciclo de trabajo alto) tienen algunas adaptaciones adicionales para detectar la frecuencia predominante (y los armónicos) de la vocalización CF. Estas incluyen una "sintonía" de frecuencia estrecha de los órganos del oído interno, con un área especialmente grande que responde a la frecuencia de los ecos de retorno del murciélago. [33]
La membrana basilar dentro de la cóclea contiene la primera de estas especializaciones para el procesamiento de la información del eco. En los murciélagos que utilizan señales de CF, la sección de la membrana que responde a la frecuencia de los ecos de retorno es mucho más grande que la región de respuesta para cualquier otra frecuencia. Por ejemplo, en el murciélago de herradura mayor, Rhinolophus ferrumequinum , hay una sección desproporcionadamente alargada y engrosada de la membrana que responde a sonidos alrededor de 83 kHz, la frecuencia constante del eco producido por el llamado del murciélago. Esta área de alta sensibilidad a un rango específico y estrecho de frecuencia se conoce como " fóvea acústica ". [54]
Los murciélagos que utilizan la ecolocalización tienen pelos cocleares que son especialmente resistentes al ruido intenso. Las células ciliadas cocleares son esenciales para la sensibilidad auditiva y pueden resultar dañadas por el ruido intenso. Como los murciélagos están expuestos regularmente a ruidos intensos a través de la ecolocalización, es necesaria la resistencia a la degradación por ruido intenso. [55]
Más adelante en la vía auditiva, el movimiento de la membrana basilar produce la estimulación de las neuronas auditivas primarias. Muchas de estas neuronas están específicamente "afinadas" (responden con mayor intensidad) al estrecho rango de frecuencia de los ecos que regresan de las llamadas de CF. Debido al gran tamaño de la fóvea acústica, la cantidad de neuronas que responden a esta región, y por lo tanto a la frecuencia del eco, es especialmente alta. [56]
En el colículo inferior , una estructura en el mesencéfalo del murciélago, la información de la parte inferior de la vía de procesamiento auditivo se integra y se envía a la corteza auditiva. Como George Pollak y otros demostraron en una serie de artículos en 1977, las interneuronas de esta región tienen un nivel muy alto de sensibilidad a las diferencias de tiempo, ya que el retraso de tiempo entre una llamada y el eco de retorno le dice al murciélago su distancia del objeto objetivo. Mientras que la mayoría de las neuronas responden más rápidamente a estímulos más fuertes, las neuronas coliculares mantienen su precisión de tiempo incluso cuando cambia la intensidad de la señal. [57] Estas interneuronas están especializadas para la sensibilidad al tiempo de varias maneras. Primero, cuando se activan, generalmente responden con solo uno o dos potenciales de acción . Esta corta duración de respuesta permite que sus potenciales de acción den una indicación específica del momento en que llegó el estímulo y respondan con precisión a estímulos que ocurren cerca en el tiempo uno del otro. Las neuronas tienen un umbral de activación muy bajo: responden rápidamente incluso a estímulos débiles. Por último, en el caso de las señales FM, cada interneurona está sintonizada con una frecuencia específica dentro del barrido, así como con esa misma frecuencia en el eco siguiente. También en este nivel hay especialización para el componente CF de la llamada. La alta proporción de neuronas que responden a la frecuencia de la fóvea acústica aumenta en realidad en este nivel. [57]
La corteza auditiva de los murciélagos es bastante grande en comparación con la de otros mamíferos. [58] Diferentes regiones de la corteza procesan diversas características del sonido, y cada una de ellas proporciona información diferente sobre la ubicación o el movimiento de un objeto objetivo. La mayoría de los estudios existentes sobre el procesamiento de la información en la corteza auditiva del murciélago han sido realizados por Nobuo Suga sobre el murciélago bigotudo, Pteronotus parnellii . El llamado de este murciélago tiene componentes de tono CF y de barrido FM. [59] [60]
Suga y sus colegas han demostrado que la corteza contiene una serie de "mapas" de información auditiva, cada uno de los cuales está organizado sistemáticamente en función de características del sonido, como la frecuencia y la amplitud . Las neuronas de estas áreas responden únicamente a una combinación específica de frecuencia y tiempo (retardo del eco del sonido) y se conocen como neuronas sensibles a la combinación. [59] [60]
Los mapas organizados sistemáticamente en la corteza auditiva responden a diversos aspectos de la señal de eco, como su retraso y su velocidad. Estas regiones están compuestas por neuronas "sensibles a la combinación" que requieren al menos dos estímulos específicos para generar una respuesta. Las neuronas varían sistemáticamente a lo largo de los mapas, que están organizados según las características acústicas del sonido y pueden ser bidimensionales. El murciélago utiliza las diferentes características del llamado y su eco para determinar características importantes de su presa. Los mapas incluyen: [59] [60]


El biosonar es valioso tanto para las ballenas dentadas (suborden Odontoceti ), incluidos los delfines , marsopas , delfines de río , orcas y cachalotes , como para las ballenas barbadas (suborden Mysticeti ), incluidas las ballenas francas , las ballenas de Groenlandia , las ballenas francas pigmeas , las ballenas grises y los rorcuales , porque viven en un hábitat submarino que tiene características acústicas favorables y donde la visión a menudo es extremadamente limitada en alcance debido a la absorción o la turbidez . [62] Los odontocetos generalmente pueden escuchar sonidos a frecuencias ultrasónicas , mientras que los misticetos escuchan sonidos dentro del régimen de frecuencia infrasónica . [63]
La evolución de los cetáceos consistió en tres radiaciones principales. A lo largo de los períodos del Eoceno medio y tardío (hace 49-31,5 millones de años), los arqueocetos , cetáceos dentados primitivos que surgieron de los mamíferos terrestres, fueron los únicos cetáceos. [64] [65] No ecolocalizaban, pero habían adaptado ligeramente la audición subacuática. [66] A finales del Eoceno medio, los huesos del oído acústicamente aislados habían evolucionado para dar a los arqueocetos basilosáuridos audición subacuática direccional en frecuencias bajas a medias. [67] Con la extinción de los arqueocetos al inicio del Oligoceno (hace 33,9-23 millones de años), dos nuevos linajes evolucionaron en una segunda radiación. Los primeros misticetos (ballenas barbadas) y odontocetos aparecieron en el Oligoceno medio en Nueva Zelanda. [65] Los odontocetos actuales son monofiléticos (un solo grupo evolutivo), pero la ecolocalización evolucionó dos veces, de manera convergente: una vez en Xenorophus , un odontoceto del tallo del Oligoceno , y otra vez en los odontocetos de la corona . [36]
La reestructuración física de los océanos ha jugado un papel en la evolución de la ecolocalización. El enfriamiento global en el límite Eoceno-Oligoceno causó un cambio de un mundo de invernadero a un mundo de hielo . Las aperturas tectónicas crearon el Océano Austral con una Corriente Circumpolar Antártica de flujo libre . [66] [67] [68] Estos eventos alentaron la selección de la capacidad de localizar y capturar presas en aguas turbias de ríos, lo que permitió a los odontocetos invadir y alimentarse a profundidades por debajo de la zona fótica . En particular, la ecolocalización por debajo de la zona fótica podría haber sido una adaptación de depredación a los cefalópodos que migran diariamente . [67] [69] La familia Delphinidae (delfines) se diversificó en el Neógeno (hace 23–2,6 millones de años), desarrollando una ecolocalización extremadamente especializada. [70] [66]
Cuatro proteínas desempeñan un papel importante en la ecolocalización de las ballenas dentadas. La prestina , una proteína motora de las células pilosas externas del oído interno de la cóclea de los mamíferos , está asociada con la sensibilidad auditiva. [71] Ha experimentado dos episodios claros de evolución acelerada en los cetáceos. [71] El primero está relacionado con la divergencia de los odontocetos, cuando se desarrolló por primera vez la ecolocalización, y el segundo con el aumento de la frecuencia de la ecolocalización entre los delfines. Tmc1 y Pjvk son proteínas relacionadas con la sensibilidad auditiva: Tmc1 está asociada con el desarrollo de las células pilosas y la audición de alta frecuencia, y Pjvk con la función de las células pilosas. [72] La evolución molecular de Tmc1 y Pjvk indica una selección positiva para la ecolocalización en los odontocetos. [72] Cldn14 , un miembro de las proteínas de unión estrecha que forman barreras entre las células del oído interno, muestra el mismo patrón evolutivo que la prestina. [73] Los dos eventos de evolución de proteínas, para Prestin y Cldn14, ocurrieron al mismo tiempo que la apertura tectónica del Pasaje de Drake (34-31 Ma) y el crecimiento del hielo antártico en la transición climática del Mioceno medio (14 Ma), con la divergencia de odontocetos y misticetos ocurriendo con el primero, y la especiación de Delphinidae con el segundo. [68]
La evolución de dos estructuras craneales puede estar vinculada a la ecolocalización. El telescopaje craneal (superposición entre los huesos frontal y maxilar , y desplazamiento hacia atrás de las fosas nasales [74] ) se desarrolló primero en los xenorófidos . Evolucionó más en los odontocetos del tallo, llegando al telescopaje craneal completo en los odontocetos de la corona. [75] El movimiento de las fosas nasales puede haber permitido un aparato nasal más grande y un melón para la ecolocalización. [75] Este cambio ocurrió después de la divergencia de los neocetos de los basilosáuridos. [76] El primer cambio hacia la asimetría craneal ocurrió en el Oligoceno Temprano, antes de los xenorófidos. [76] Un fósil de xenorófido ( Cotylocara macei ) tiene asimetría craneal y muestra otros indicadores de ecolocalización. [77] Sin embargo, los xenorófidos basales carecen de asimetría craneal, lo que indica que esto probablemente evolucionó dos veces. [76] Los odontocetos actuales tienen regiones nasofaciales asimétricas; generalmente, el plano medio está desplazado hacia la izquierda y las estructuras de la derecha son más grandes. [77] Tanto el telescopio craneal como la asimetría probablemente se relacionan con la producción de sonido para la ecolocalización. [75]
Trece especies de odontocetos actuales desarrollaron de manera convergente la ecolocalización de alta frecuencia de banda estrecha (NBHF) en cuatro eventos separados. Estas especies incluyen las familias Kogiidae (cachalotes pigmeos) y Phocoenidae (marsopas), así como algunas especies del género Lagenorhynchus , todas las de Cephalorhynchus y el delfín de La Plata . Se cree que la NBHF evolucionó como un medio de evasión de depredadores; las especies productoras de NBHF son pequeñas en relación con otros odontocetos, lo que las convierte en presas viables para especies grandes como la orca . Sin embargo, debido a que tres de los grupos desarrollaron NBHF antes de la aparición de la orca, la depredación por parte de otros odontocetos rapaces antiguos debe haber sido la fuerza impulsora para el desarrollo de NBHF, no la depredación por parte de la orca. Las orcas y, presumiblemente, los odontocetos rapaces antiguos como Acrophyseter , son incapaces de escuchar frecuencias superiores a 100 kHz. [78]
Otra razón para la variación en la ecolocalización es el hábitat. Para todos los sistemas de sonar, el factor limitante que decide si se detecta un eco de retorno es la relación eco-ruido (ENR). La ENR está dada por el nivel de fuente emitido (SL) más la intensidad del objetivo, menos la pérdida de transmisión bidireccional (absorción y propagación) y el ruido recibido. [79] Los animales se adaptarán para maximizar el alcance en condiciones de ruido limitado (aumentar el nivel de fuente) o para reducir el ruido en un hábitat poco profundo y/o lleno de basura (disminuir el nivel de fuente). En hábitats desordenados, como las áreas costeras, los rangos de presa son más pequeños, y especies como el delfín de Commerson ( Cephalorhynchus commersonii ) han reducido los niveles de fuente para adaptarse mejor a su entorno. [79]
Las ballenas dentadas emiten un haz enfocado de chasquidos de alta frecuencia en la dirección en la que apunta su cabeza. Los sonidos se generan al pasar aire desde las fosas nasales óseas a través de los labios fónicos . Estos sonidos se reflejan en el denso hueso cóncavo del cráneo y un saco de aire en su base. El haz enfocado es modulado por un gran órgano graso conocido como melón. Este actúa como una lente acústica porque está compuesto de lípidos de diferentes densidades. La mayoría de las ballenas dentadas utilizan chasquidos en una serie, o tren de chasquidos, para la ecolocalización, mientras que el cachalote puede producir chasquidos individualmente. Los silbidos de las ballenas dentadas no parecen usarse en la ecolocalización. Diferentes tasas de producción de chasquidos en un tren de chasquidos dan lugar a los familiares ladridos, chillidos y gruñidos del delfín mular . Un tren de chasquidos con una tasa de repetición de más de 600 por segundo se llama pulso de ráfaga. En los delfines mulares, la respuesta cerebral auditiva resuelve clics individuales hasta 600 por segundo, pero produce una respuesta graduada para tasas de repetición más altas. [80]
Se ha sugerido que la disposición de los dientes de algunas ballenas dentadas más pequeñas puede ser una adaptación para la ecolocalización. [81] Los dientes de un delfín mular, por ejemplo, no están dispuestos simétricamente cuando se los observa desde un plano vertical. Esta asimetría podría ser una ayuda para detectar si los ecos de su biosonar provienen de un lado o del otro; pero esto no se ha comprobado experimentalmente. [82]
Los ecos se reciben utilizando estructuras grasas complejas alrededor de la mandíbula inferior como vía de recepción primaria, desde donde se transmiten al oído medio a través de un cuerpo graso continuo. El sonido lateral puede recibirse a través de los lóbulos grasos que rodean las orejas con una densidad similar a la del agua. Algunos investigadores creen que cuando se acercan al objeto de interés, se protegen contra el eco más fuerte silenciando el sonido emitido. En los murciélagos se sabe que esto sucede, pero aquí la sensibilidad auditiva también se reduce cerca de un objetivo. [83] [84]
_hunting_by_echolocation.JPG/1280px-Palawan_swiftlet_(Aerodramus_palawanensis)_hunting_by_echolocation.JPG)
Se sabe que los guácharos y algunas especies de vencejos utilizan una forma de ecolocalización relativamente rudimentaria en comparación con la de los murciélagos y los delfines. Estas aves nocturnas emiten llamadas mientras vuelan y las utilizan para orientarse a través de los árboles y las cuevas donde viven. [85] [86]
Los mamíferos terrestres distintos de los murciélagos que se sabe o se cree que ecolocalizan incluyen musarañas , [87] [88] [89] los tenrecs de Madagascar , [90] lirones pigmeos chinos , [91] y solenodontes . [92] Los sonidos de las musarañas, a diferencia de los de los murciélagos, son de baja amplitud, banda ancha, multiarmónicos y modulados en frecuencia. [89] No contienen clics de ecolocalización con reverberaciones y parecen usarse para una orientación espacial simple y de corto alcance. A diferencia de los murciélagos, las musarañas usan la ecolocalización solo para investigar su hábitat en lugar de localizar comida. [89] Hay evidencia de que las ratas de laboratorio cegadas pueden usar la ecolocalización para navegar por laberintos. [93]

Algunos insectos que son depredados por murciélagos tienen adaptaciones antidepredadores , incluyendo la evitación de depredadores, [95] la desviación del ataque, [94] y chasquidos ultrasónicos que parecen funcionar como advertencias en lugar de bloquear la ecolocalización . [49] [96]
Las polillas tigre ( Arctiidae ) de diferentes especies (dos tercios de las especies analizadas) responden a ataques simulados mediante la ecolocalización de murciélagos mediante la producción de una serie acelerada de chasquidos. Se ha demostrado que la especie Bertholdia trigona bloquea la ecolocalización de los murciélagos: cuando se enfrentó a murciélagos pardos grandes ingenuos, el ultrasonido fue inmediatamente y consistentemente efectivo para prevenir el ataque de los murciélagos. Los murciélagos entraron en contacto con polillas de control silenciosas un 400% más a menudo que con B. trigona . [97]
Los ultrasonidos de las polillas también pueden servir para asustar al murciélago (una táctica de engaño), advertirle de que la polilla es desagradable (señalización honesta, aposematismo ) o imitar a especies que se defienden químicamente. Se ha demostrado que tanto el aposematismo como el mimetismo confieren una ventaja de supervivencia contra los ataques de murciélagos. [98] [99]
La polilla de cera mayor ( Galleria mellonella ) realiza acciones para evitar a los depredadores, como tirarse al suelo, dar vueltas y quedarse quieta cuando detecta ondas ultrasónicas, lo que indica que puede detectar y diferenciar entre las frecuencias ultrasónicas utilizadas por los depredadores y las señales de otros miembros de su especie. [95] Algunos miembros de la familia de polillas Saturniidae , que incluye polillas de seda gigantes, tienen colas largas en las alas traseras, especialmente las de los subgrupos Attacini y Arsenurinae . Las colas oscilan en vuelo, creando ecos que desvían el ataque del murciélago cazador del cuerpo de la polilla a las colas. La especie Argema mimosae (la polilla lunar africana), que tiene colas especialmente largas, fue la que tuvo más probabilidades de evadir la captura. [94]
De la pág. 140: De estos experimentos el autor concluye: … que el órgano del oído parece suplir al de la vista en el descubrimiento de cuerpos, y proporcionar a estos animales diferentes sensaciones para dirigir su vuelo y permitirles evitar los obstáculos que puedan presentarse.
Los murciélagos y los delfines son conocidos por su capacidad de utilizar la ecolocalización. ... algunas personas ciegas han aprendido a hacer lo mismo ...
