Experimento a largo plazo de la evolución de E. Coli
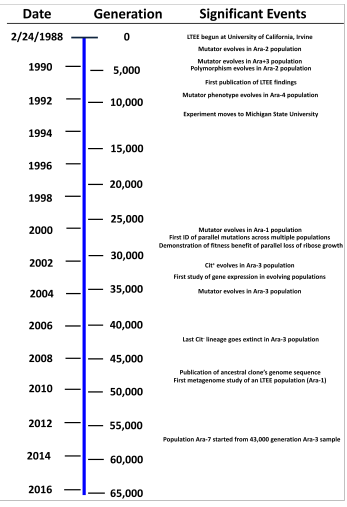
La adaptación más sorprendente reportada hasta ahora es la evolución de crecimiento aeróbico con citrato, el cual es raro en E. coli, en una población en algún momento entre las generaciones 31 000 y 31 500.
Además, debido al gran uso de E. coli como un organismo modelo principal en biología molecular, una gran variedad de herramientas, protocolos y procedimientos eran accesibles para estudiar los cambios a nivel genotípico, fenotípico y fisiológico.
[7] Lenski decidió realizar el experimento con la bacteria en un medio de cultivo con glucosa limitada llamado DM25,[10] el cual fue inicialmente desarrollado por Bernard Davis para aislar las mutantes auxótrofas de E. coli utilizando penicilina como un agente selectivo.
Las bacterias en estas muestras permanecen viables y pueden ser revividas en cualquier momento.
Estas diferencias permiten que se diferencien grupos de cepas en Tetrazolium Arabinose plates, sobre las cuales las colonias Ara- aparecen rojas mientras que las colonias Ara+ son de blancas a rosas.
Para las 20 000 generaciones, las poblaciones crecían aproximadamente un 70% más deprisa que la cepa ancestral.
[23] A lo largo del experimento, las poblaciones han evolucionado para especializarse en la glucosa en la que crecen.
Su análisis sugirió que este deterioro se debía a pleiotropía antagonística en la cual mutaciones que mejoraban la habilidad de crecer en glucosa habían reducido o eliminado la habilidad de crecer en otras substancias.
Este fue constatado por el descubrimiento de que la variante L tenía una ventaja en el crecimiento en glucosa, pero S tenía una ventaja durante la fase estacionaria, después de que la glucosa se había agotado.
Las dos variantes fueron encontradas que habían evolucionado inicialmente antes de las 6000 generaciones, y coexistido desde entonces.
[34] Este mutante había sido aislado durante la selección prolongada de otra nueva substancia para crecer en un caldo que también contenía citrato.
Cerca de la generación 33,127, se observó un incremento dramático en la población designada como Ara-3.
Esta capacidad metabólica permitió a la población crecer varias veces más grande de lo que lo había hecho previamente, debido a las grandes cantidades de citrato presentes en el medio.
[5] Lenski y sus compañeros concluyeron que la evolución de la función Cit+ en esta población en partiular surgió debido a una o más mutaciones previas, posiblemente no adaptativas, "potenciadoras" que incrementaron la velocidad de mutación a un nivel accesible.
Más generalmente, los autores sugieren que los resultados indican, siguiendo el argumento de Stephen Jay Gould, "que la contingencia histórica puede tener un profundo y duradero impacto" en el curso de la evolución.
[5] Estos resultados han sido considerados como una instancia significativa del impacto de la contingencia histórica sobre la evolución.
Sin embargo, el fenotipo Cit+ inicial proporcionado por la duplicación era muy débil y solo daba una aptitud benéfica de ~1%.
Mutaciones adicionales después de que la bacteria Cit+ fuera dominante en la población continuaron acumulándose para mejorar el crecimiento en citrato.
sugirió que este patrón puede ser típico de cómo nuevos caracteres evolucionan en general y propuso un modelos de tres pasos para la innovación evolutiva: Este modelo ha sido aceptado en biología evolutiva.
[39] Este método usa múltiples rondas de un proceso en donde conjugación basada en plásmidos F a 33 000 generation Cit+ clone, CZB154, and the Cit- founding clone of the LTEE to purge mutations not required for either manifestation of a weak or strong form of the Cit+ trait, which they refer to as Cit++.
Esta mutación, encontrada en una región reguladora de un gen llamado dctA, causó un aumento masivo en la expresión del transportador DctA, el cual funciona para importar dicarboxilatos C4 a la célula y había ocurrido en el linaje que lo condujo.
concluyeron que la mutación dctA no era involucrada en la potenciación, sino en el refinamiento.
Esto los condujo a sugerir que la evolución de Cit+ en la población Ara-3 pudo haber sido contingente de un perfil genético y ecología específica de la población que permitió las variantes tempranas y débiles Cit+ a mantenerse en la población lo suficiente para refinar mutaciones que surgieran y hacer el crecimiento en citrato suficientemente fuerte para proveer una aptitud significativamente benéfica.
Quandt y sus compañeros publicaron más tarde los resultados identificando definitivamente una mutación que potenció la evolución Cit+.
Después de que evolucionó el fenotipo fuerte Cit++, la actividad citrato sintasa se volvió perjudicial.
Concluyeron que las series e mutaciones en gltA primero potenciaron y después refinaron el crecimeinto en citrato.
En efecto, clones Cit- podían invadir las poblaciones Cit+ después del evento de extinción.
[43] Otros investigadores han experimentado hacer evolucionar E. coli aerobio que use citrato.
[44] John Roth y Sophie Maisnier-Patin revisaron ambas perspectivas tanto en las mutaciones tardadas del equipo de Lenski como en las mutaciones rápidas del equipo de Van Hofweges en E. coli.
[47] Sin embargo, él dice que la especiación es generalmente considerada por biólogos evolutivos como un proceso y no un evento.