Los cuatro géneros del virus de la influenza que infectan a los vertebrados, que se identifican por diferencias antigénicas en su nucleoproteína y proteína matriz , son los siguientes:
El virión del virus de la influenza es pleomórfico ; la envoltura viral puede presentarse en formas esféricas y filamentosas. En general, la morfología del virus es elipsoidal con partículas de 100 a 120 nm de diámetro, o filamentosa con partículas de 80 a 100 nm de diámetro y hasta 20 μm de largo. [5] Hay aproximadamente 500 proyecciones superficiales distintas en forma de espiga en la envoltura, cada una de las cuales se proyecta a 10 a 14 nm de la superficie con densidades superficiales variables. La espiga de la glucoproteína principal (HA) está interpuesta irregularmente por grupos de espigas de neuraminidasa (NA), con una relación de HA a NA de aproximadamente 10 a 1. [6]
La envoltura viral, compuesta por una membrana de bicapa lipídica en la que se anclan las espigas de glicoproteínas, encierra las nucleocápsides , nucleoproteínas de diferentes clases de tamaño con un bucle en cada extremo; la disposición dentro del virión es incierta. Las proteínas ribonucleares son filamentosas y miden entre 50 y 130 nm de largo y entre 9 y 15 nm de diámetro con simetría helicoidal. [ cita requerida ]
Las proteínas del virus de la influenza mejor caracterizadas son la hemaglutinina y la neuraminidasa , dos grandes glicoproteínas que se encuentran en el exterior de las partículas virales. La hemaglutinina es una lectina que media la unión del virus a las células diana y la entrada del genoma viral en la célula diana. [9] Por el contrario, la neuraminidasa es una enzima involucrada en la liberación del virus progenie de las células infectadas, al escindir los azúcares que se unen a las partículas virales maduras. Las proteínas hemaglutinina (H) y neuraminidasa (N) son objetivos clave para los anticuerpos y los medicamentos antivirales, [10] [11] y se utilizan para clasificar los diferentes serotipos de los virus de la influenza A, de ahí la H y la N en H5N1 .
La secuencia del genoma tiene secuencias repetidas terminales; repetidas en ambos extremos. Las repeticiones terminales en el extremo 5' tienen una longitud de 12 a 13 nucleótidos. Las secuencias de nucleótidos del extremo 3' son idénticas; las mismas en géneros de la misma familia; la mayoría en el ARN (segmentos) o en todas las especies de ARN. Las repeticiones terminales en el extremo 3' tienen una longitud de 9 a 11 nucleótidos. El ácido nucleico encapsidado es exclusivamente genómico. Cada virión puede contener copias interferentes defectuosas. En la influenza A (H1N1), PB1-F2 se produce a partir de un marco de lectura alternativo en PB1. Los genes M y NS producen dos genes diferentes a través de un empalme alternativo . [12]
Ciclo de replicación
Infección y replicación del virus de la gripe. Los pasos de este proceso se describen en el texto.
Por lo general, la gripe se transmite de mamíferos infectados a través del aire mediante la tos o los estornudos, que crean aerosoles que contienen el virus, y de aves infectadas a través de sus excrementos . La gripe también puede transmitirse por saliva , secreciones nasales , heces y sangre . Las infecciones se producen a través del contacto con estos fluidos corporales o con superficies contaminadas. Fuera de un huésped, los virus de la gripe pueden seguir siendo infecciosos durante aproximadamente una semana a la temperatura corporal humana, más de 30 días a 0 °C (32 °F) e indefinidamente a temperaturas muy bajas (como los lagos del noreste de Siberia ). Pueden inactivarse fácilmente con desinfectantes y detergentes . [13] [14] [15]
Los virus se unen a una célula a través de interacciones entre su glucoproteína hemaglutinina y azúcares de ácido siálico en las superficies de las células epiteliales en el pulmón y la garganta (Etapa 1 en la figura de infección). [16] La célula importa el virus por endocitosis . En el endosoma ácido , parte de la proteína hemaglutinina fusiona la envoltura viral con la membrana de la vacuola, liberando las moléculas de ARN viral (ARNv), las proteínas accesorias y la ARN polimerasa dependiente de ARN en el citoplasma (Etapa 2). [17] Estas proteínas y el ARNv forman un complejo que se transporta al núcleo celular , donde la ARN polimerasa dependiente de ARN comienza a transcribir el ARNc de sentido positivo complementario (Pasos 3a y b). [18] El ARNc se exporta al citoplasma y se traduce (paso 4), o permanece en el núcleo. Las proteínas virales recién sintetizadas se secretan a través del aparato de Golgi hacia la superficie celular (en el caso de la neuraminidasa y la hemaglutinina, paso 5b) o se transportan de regreso al núcleo para unirse al ARNv y formar nuevas partículas del genoma viral (paso 5a). Otras proteínas virales tienen múltiples acciones en la célula huésped, incluida la degradación del ARNm celular y el uso de los nucleótidos liberados para la síntesis del ARNv y también la inhibición de la traducción de los ARNm de la célula huésped. [19]
Los ARNv de sentido negativo que forman los genomas de los futuros virus, la ARN transcriptasa dependiente de ARN y otras proteínas virales se ensamblan en un virión. Las moléculas de hemaglutinina y neuraminidasa se agrupan en una protuberancia en la membrana celular. El ARNv y las proteínas del núcleo viral abandonan el núcleo y entran en esta protuberancia de la membrana (paso 6). El virus maduro se desprende de la célula en una esfera de membrana fosfolipídica del huésped, adquiriendo hemaglutinina y neuraminidasa con esta capa de membrana (paso 7). [20] Como antes, los virus se adhieren a la célula a través de la hemaglutinina; los virus maduros se desprenden una vez que su neuraminidasa ha escindido los residuos de ácido siálico de la célula huésped. [16] Después de la liberación del nuevo virus de la gripe, la célula huésped muere.
Transcripción de ARNm iniciada por la polimerasa viral mediante el método cap snatching
Los virus Orthomyxoviridae son uno de los dos virus de ARN que se replican en el núcleo (el otro es Retroviridae ). Esto se debe a que la maquinaria de los virus Orthomyxo no puede producir sus propios ARNm. Utilizan ARN celulares como cebadores para iniciar la síntesis de ARNm viral en un proceso conocido como arrebato de capuchón . [21] Una vez en el núcleo, la proteína ARN polimerasa PB2 encuentra un pre-ARNm celular y se une a su extremo con capuchón 5'. Luego, la ARN polimerasa PA escinde el ARNm celular cerca del extremo 5' y utiliza este fragmento con capuchón como cebador para transcribir el resto del genoma del ARN viral en ARNm viral. [22] Esto se debe a la necesidad de que el ARNm tenga un capuchón 5' para ser reconocido por el ribosoma de la célula para la traducción.
Como no existen enzimas correctoras de ARN , la transcriptasa de ARN dependiente de ARN comete un error de inserción de un solo nucleótido aproximadamente cada 10 mil nucleótidos, que es la longitud aproximada del ARNv de la gripe. Por lo tanto, casi todos los virus de la gripe recién fabricados contendrán una mutación en su genoma. [23] La separación del genoma en ocho segmentos separados de ARNv permite la mezcla ( reassortment ) de los genes si más de una variedad del virus de la gripe ha infectado la misma célula ( superinfección ). La alteración resultante en los segmentos del genoma empaquetados en la progenie viral confiere un nuevo comportamiento, a veces la capacidad de infectar nuevas especies hospedadoras o de superar la inmunidad protectora de las poblaciones hospedadoras a su antiguo genoma (en cuyo caso se denomina cambio antigénico ). [10]
Existen cuatro géneros de virus de la gripe, cada uno de los cuales contiene una única especie o tipo. Los virus de la gripe A y C infectan a una variedad de especies (incluidos los seres humanos), mientras que los de la gripe B infectan casi exclusivamente a los seres humanos y los de la gripe D infectan al ganado vacuno y a los cerdos. [26] [27] [28]
Gripe A
Diagrama de nomenclatura de la influenza
Los virus de la influenza A se clasifican además según las proteínas de superficie virales hemaglutinina (HA o H) y neuraminidasa (NA o N). Se han aislado en la naturaleza 18 subtipos HA (o serotipos) y 11 subtipos NA del virus de la influenza A. Entre ellos, los subtipos HA 1-16 y NA 1-9 se encuentran en aves acuáticas y playeras silvestres, y los subtipos HA 17-18 y NA 10-11 solo se han aislado de murciélagos. [29] [30]
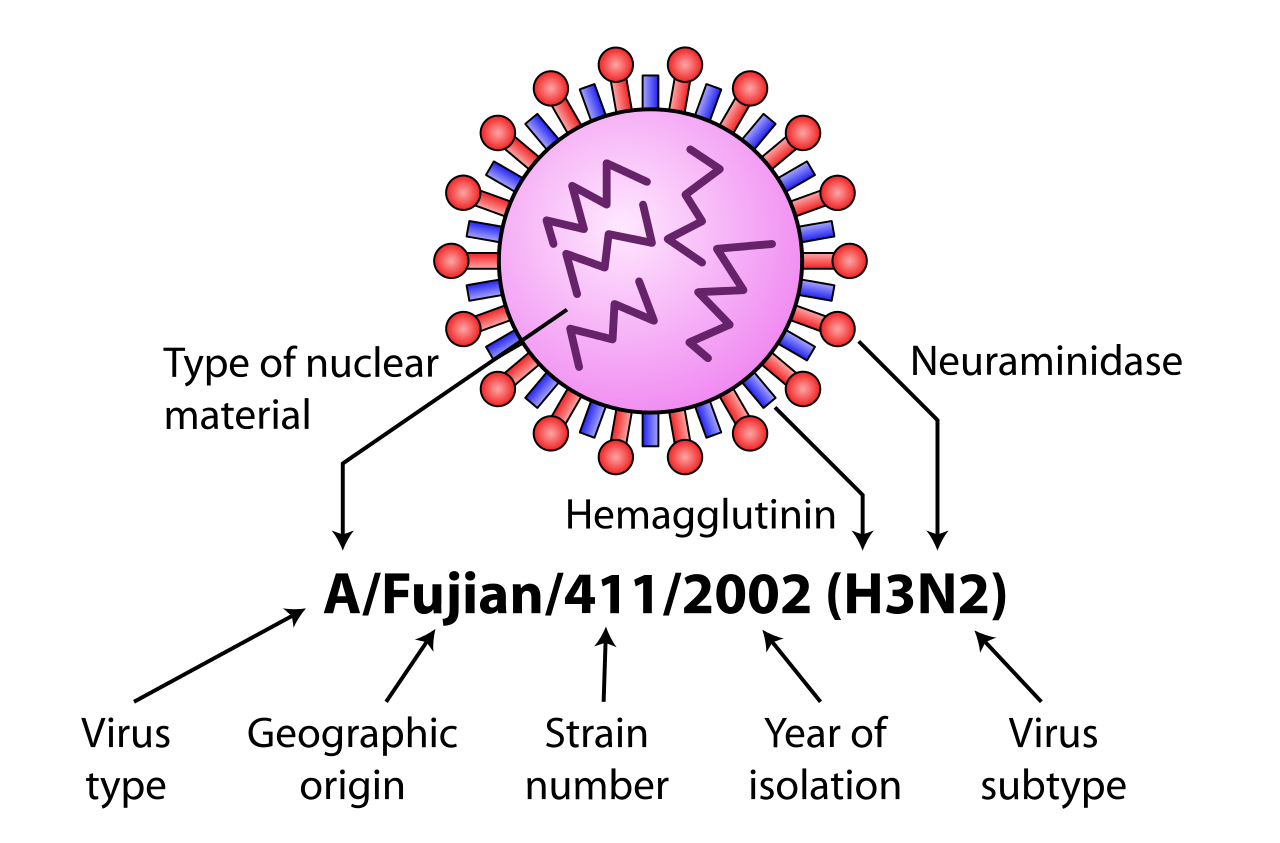
Existe una variación adicional; por lo tanto, los aislamientos de cepas de influenza específicas se identifican mediante la nomenclatura del virus de la influenza, [31] especificando el tipo de virus, la especie huésped (si no es humana), la ubicación geográfica donde se aisló por primera vez, la referencia de laboratorio, el año de aislamiento y los subtipos HA y NA. [32] [33]
Ejemplos de nomenclatura son:
A/Brisbane/59/2007 (H1N1) - aislado de un ser humano
A/cerdo/Dakota del Sur/152B/2009 (H1N2) - aislado de un cerdo
Los virus de influenza tipo A son los patógenos humanos más virulentos entre los tres tipos de influenza y causan la enfermedad más grave. Se cree que todos los virus de influenza A que causan brotes o pandemias se originan en aves acuáticas salvajes. [34] Todas las pandemias del virus de influenza A desde la década de 1900 fueron causadas por influenza aviar , a través de recombinación con otras cepas de influenza, ya sea aquellas que afectan a los humanos (gripe estacional) o aquellas que afectan a otros animales (ver pandemia de gripe porcina de 2009 ). [35] Los serotipos que se han confirmado en humanos , ordenados por el número de muertes humanas confirmadas, son:
El virus de la influenza B es casi exclusivamente un patógeno humano y es menos común que la influenza A. El único otro animal conocido que es susceptible a la infección por influenza B es la foca . [47] Este tipo de influenza muta a una tasa 2-3 veces menor que el tipo A [48] y, en consecuencia, es menos diverso genéticamente, con solo un serotipo de influenza B. [26] Como resultado de esta falta de diversidad antigénica , un grado de inmunidad a la influenza B generalmente se adquiere a una edad temprana. Sin embargo, la influenza B muta lo suficiente como para que no sea posible una inmunidad duradera. [49] Esta tasa reducida de cambio antigénico, combinada con su rango limitado de hospedadores (inhibiendo el cambio antigénico entre especies ), asegura que no ocurran pandemias de influenza B. [50]
Este es un género que fue clasificado en 2016, cuyos miembros fueron aislados por primera vez en 2011. [54] Este género parece estar más estrechamente relacionado con la influenza C, de la cual divergió hace varios cientos de años. [55] Hay al menos dos cepas existentes de este género. [56] Los principales huéspedes parecen ser el ganado, pero se sabe que el virus también infecta a los cerdos.
Viabilidad y desinfección
Los virus de la influenza de los mamíferos tienden a ser lábiles, pero pueden sobrevivir varias horas en el moco. [57] El virus de la influenza aviar puede sobrevivir durante 100 días en agua destilada a temperatura ambiente y 200 días a 17 °C (63 °F). El virus aviar se inactiva más rápidamente en el estiércol, pero puede sobrevivir hasta dos semanas en las heces de las jaulas. Los virus de la influenza aviar pueden sobrevivir indefinidamente cuando se congelan. [57] Los virus de la influenza son susceptibles a la lejía, al etanol al 70 %, a los aldehídos, a los agentes oxidantes y a los compuestos de amonio cuaternario. Se inactivan con calor de 133 °F (56 °C) durante un mínimo de 60 minutos, así como con un pH bajo <2. [57]
Vacunación y profilaxis
Objetivos de los agentes antigripales que están autorizados o bajo investigación
Existen vacunas y medicamentos para la profilaxis y el tratamiento de las infecciones por el virus de la gripe. Las vacunas están compuestas de viriones inactivados o vivos atenuados de los virus de la gripe A H1N1 y H3N2, así como de los virus de la gripe B. Debido a que las antigenicidades de los virus salvajes evolucionan, las vacunas se reformulan anualmente actualizando las cepas de siembra. [ cita requerida ]
Cuando las antigenicidades de las cepas de semillas y los virus salvajes no coinciden, las vacunas no protegen a los vacunados. [ cita requerida ] Además, incluso cuando coinciden, a menudo se generan mutantes de escape. [ cita requerida ]
Los fármacos disponibles para el tratamiento de la gripe incluyen amantadina y rimantadina , que inhiben la pérdida de la envoltura de los viriones al interferir con el canal de protones M2 , y oseltamivir (comercializado bajo la marca Tamiflu ), zanamivir y peramivir , que inhiben la liberación de viriones de las células infectadas al interferir con NA. Sin embargo, a menudo se generan mutantes de escape para el primer fármaco y con menor frecuencia para el segundo. [58]
^ Índice de virus del Comité Internacional de Taxonomía de Virus — Orthomyxovirus (2006). En: ICTVdB—The Universal Virus Database, versión 4. Büchen-Osmond, C (Ed), Universidad de Columbia, Nueva York.
^ Jones LD, Nuttall PA (1989). "Transmisión no virémica del virus Thogoto: influencia del tiempo y la distancia". Trans. R. Soc. Trop. Med. Hyg . 83 (5): 712–14. doi :10.1016/0035-9203(89)90405-7. PMID 2617637.
^ Ely B (1999). "Infectious Salmon Anaemia". Mill Hill Essays . Instituto Nacional de Investigación Médica . Archivado desde el original el 24 de agosto de 2007. Consultado el 14 de septiembre de 2007 .
^ Raynard RS, Murray AG, Gregory A (2001). "Virus de la anemia infecciosa del salmón en peces salvajes de Escocia". Dis. Aquat. Org . 46 (2): 93–100. doi : 10.3354/dao046093 . PMID: 11678233.
^ Noda T (3 de enero de 2012). "Morfología nativa de los viriones de influenza". Frontiers in Microbiology . 2 : 269. doi : 10.3389/fmicb.2011.00269 . PMC 3249889 . PMID 22291683.
^ Einav T, Gentles LE, Bloom JD (23 de julio de 2020). "Instantánea: la gripe en cifras" (PDF) . Cell . 182 (2): 532–532.e1. doi :10.1016/j.cell.2020.05.004. PMID 32707094. S2CID 220715148.
^ "Noveno informe del ICTV; publicación de taxonomía de 2009: Orthomyxoviridae". ICTV . Consultado el 19 de septiembre de 2020 .
^ Ghedin E, Sengamalay NA, Shumway M, Zaborsky J, Feldblyum T, Subbu V, Spiro DJ, Sitz J, Koo H, Bolotov P, Dernovoy D, Tatusova T, Bao Y, St George K, Taylor J, Lipman DJ, Fraser CM, Taubenberger JK, Salzberg SL (octubre de 2005). "La secuenciación a gran escala de la gripe humana revela la naturaleza dinámica de la evolución del genoma viral". Nature . 437 (7062): 1162–6. Bibcode :2005Natur.437.1162G. doi : 10.1038/nature04239 . PMID 16208317.
^ Suzuki Y (marzo de 2005). "Sialobiología de la gripe: mecanismo molecular de la variación del rango de hospedadores de los virus de la gripe". Boletín biológico y farmacéutico . 28 (3): 399–408. doi : 10.1248/bpb.28.399 . PMID 15744059.
^ abc Hilleman MR (agosto de 2002). "Realidades y enigmas de la gripe viral humana: patogénesis, epidemiología y control". Vaccine . 20 (25–26): 3068–87. doi :10.1016/S0264-410X(02)00254-2. PMID 12163258.
^ Wilson JC, von Itzstein M (julio de 2003). "Estrategias recientes en la búsqueda de nuevas terapias contra la gripe". Current Drug Targets . 4 (5): 389–408. doi :10.2174/1389450033491019. PMID 12816348.
^ Bouvier NM, Palese P (septiembre de 2008). "La biología de los virus de la gripe". Vaccine . 26 (Supl 4): D49–53. doi :10.1016/j.vaccine.2008.07.039. PMC 3074182 . PMID 19230160.
^ Suarez DL, Spackman E, Senne DA, Bulaga L, Welsch AC, Froberg K (2003). "El efecto de varios desinfectantes en la detección del virus de la influenza aviar mediante RT-PCR en tiempo real". Enfermedades aviares . 47 (3 Suppl): 1091–5. doi :10.1637/0005-2086-47.s3.1091. PMID 14575118. S2CID 8612187.
^ "Implicaciones de la influenza aviar (gripe aviar) para las enfermedades humanas. Características físicas de los virus de influenza A". CIDRAP - Centro de Investigación y Política de Enfermedades Infecciosas . Universidad de Minnesota. 12 de marzo de 2024.
^ "Los virus de la gripe 'pueden vivir décadas' en el hielo". The New Zealand Herald . Reuters . 30 de noviembre de 2006 . Consultado el 1 de noviembre de 2011 .
^ ab Wagner R, Matrosovich M, Klenk H (mayo-junio de 2002). "Equilibrio funcional entre la hemaglutinina y la neuraminidasa en las infecciones por el virus de la gripe". Rev Med Virol . 12 (3): 159–66. doi :10.1002/rmv.352. PMID 11987141. S2CID 30876482.
^ Lakadamyali M, Rust M, Babcock H, Zhuang X (5 de agosto de 2003). "Visualización de la infección de virus de influenza individuales". Proc Natl Acad Sci USA . 100 (16): 9280–85. Bibcode :2003PNAS..100.9280L. doi : 10.1073/pnas.0832269100 . PMC 170909 . PMID 12883000.
^ Cros J, Palese P (septiembre de 2003). "Tráfico de ARN genómico viral dentro y fuera del núcleo: virus de la influenza, de la enfermedad de Thogoto y de Borna". Virus Res . 95 (1–2): 3–12. doi :10.1016/S0168-1702(03)00159-X. PMID 12921991.
^ Kash J, Goodman A, Korth M, Katze M (julio de 2006). "Secuestro de la respuesta de la célula huésped y control de la traducción durante la infección por el virus de la gripe". Virus Res . 119 (1): 111–20. doi :10.1016/j.virusres.2005.10.013. PMID 16630668.
^ Nayak D, Hui E, Barman S (diciembre de 2004). "Ensamblaje y gemación del virus de la influenza". Virus Res . 106 (2): 147–65. doi :10.1016/j.virusres.2004.08.012. PMC 7172797 . PMID 15567494.
^ "Cap Snatching". ViralZone . expasy . Consultado el 11 de septiembre de 2014 .
^ Dias A, Bouvier D, Crépin T, McCarthy AA, Hart DJ, Baudin F, Cusack S, Ruigrok RW (abril de 2009). "La endonucleasa cap-snatching de la polimerasa del virus de la gripe reside en la subunidad PA". Nature . 458 (7240): 914–8. Bibcode :2009Natur.458..914D. doi :10.1038/nature07745. PMID 19194459. S2CID 4421958.
^ Drake J (1 de mayo de 1993). "Tasas de mutación espontánea entre virus ARN". Proc Natl Acad Sci USA . 90 (9): 4171–5. Bibcode :1993PNAS...90.4171D. doi : 10.1073/pnas.90.9.4171 . PMC 46468 . PMID 8387212.
^ Biere B, Bauer B, Schweiger B (abril de 2010). "Diferenciación de los linajes del virus de la influenza B Yamagata y Victoria mediante PCR en tiempo real" (PDF) . Journal of Clinical Microbiology . 48 (4): 1425–7. doi :10.1128/JCM.02116-09. PMC 2849545. PMID 20107085 .
^ Historia de la taxonomía ICTV, ICTV, 2014, archivado desde el original el 2 de abril de 2015 , consultado el 6 de junio de 2006
^ ab Hay A, Gregory V, Douglas A, Lin Y (29 de diciembre de 2001). "La evolución de los virus de la gripe humana". Philos Trans R Soc Lond B Biol Sci . 356 (1416): 1861–70. doi :10.1098/rstb.2001.0999. PMC 1088562. PMID 11779385 .
^ "Influenza aviar (gripe aviar)". Centros para el Control y la Prevención de Enfermedades . Consultado el 15 de septiembre de 2007 .
^ Kumar, Binod; Asha, Kumari; Khanna, Madhu; Ronsard, Larance; Meseko, Clement Adebajo; Sanicas, Melvin (abril de 2018). "La amenaza emergente del virus de la influenza: estado y nuevas perspectivas para su terapia y control". Archivos de Virología . 163 (4): 831–844. doi :10.1007/s00705-018-3708-y. ISSN 1432-8798. PMC 7087104 . PMID 29322273.
^ Alexander, Dennis J. (26 de julio de 2007). "Una visión general de la epidemiología de la influenza aviar". Vaccine . 25 (30): 5637–5644. doi :10.1016/j.vaccine.2006.10.051. ISSN 0264-410X. PMID 17126960.
^ Wu, Ying; Wu, Yan; Tefsen, Boris; Shi, Yi; Gao, George F. (abril de 2014). "Virus similares a la influenza derivados de murciélagos H17N10 y H18N11". Tendencias en microbiología . 22 (4): 183–191. doi :10.1016/j.tim.2014.01.010. ISSN 0966-842X. PMC 7127364 . PMID 24582528.
^ "Una revisión del sistema de nomenclatura de los virus de la gripe: un memorando de la OMS". Bull World Health Organ . 58 (4): 585–591. 1980. PMC 2395936 . PMID 6969132. Este memorando fue redactado por los firmantes enumerados en la página 590 con ocasión de una reunión celebrada en Ginebra en febrero de 1980.
^ Atkinson W, Hamborsky J, McIntyre L, Wolfe S, eds. (2007). Epidemiología y prevención de enfermedades prevenibles mediante vacunación (10.ª ed.). Washington DC: Centros para el Control y la Prevención de Enfermedades.
^ "Influenza aviar (gripe aviar): implicaciones para las enfermedades humanas". Centro de investigación y políticas sobre enfermedades infecciosas, Universidad de Minnesota . 27 de junio de 2007. Consultado el 14 de septiembre de 2007 .
^ Webster, RG; Bean, WJ; Gorman, OT; Chambers, TM; Kawaoka, Y. (marzo de 1992). "Evolución y ecología de los virus de la influenza A". Microbiological Reviews . 56 (1): 152–179. doi :10.1128/mr.56.1.152-179.1992. ISSN 0146-0749. PMC 372859 . PMID 1579108.
^ Taubenberger, Jeffery K.; Morens, David M. (abril de 2010). "Influenza: la pandemia pasada y futura". Public Health Reports . 125 (Supl 3): 16–26. doi :10.1177/00333549101250S305. ISSN 0033-3549. PMC 2862331 . PMID 20568566.
^ Wang TT, Palese P (junio de 2009). "Descifrando el misterio del virus de la gripe porcina". Cell . 137 (6): 983–85. doi : 10.1016/j.cell.2009.05.032 . PMID 19524497.
^ Taubenberger, JK, Morens, DM (abril de 2009). "Influenza pandémica: incluida una evaluación de riesgos del virus H5N1". Rev. Sci. Tech. Off. Int. Epiz . 28 (1): 187–202. doi :10.20506/rst.28.1.1879. PMC 2720801. PMID 19618626 .
^ Fouchier R, Schneeberger P, Rozendaal F, Broekman J, Kemink S, Munster V, Kuiken T, Rimmelzwaan G, Schutten M, Van Doornum G, Koch G, Bosman A, Koopmans M, Osterhaus A (2004). "Virus de la influenza aviar A (H7N7) asociado con conjuntivitis humana y un caso fatal de síndrome de dificultad respiratoria aguda". Proc Natl Acad Sci Estados Unidos . 101 (5): 1356–61. Código bibliográfico : 2004PNAS..101.1356F. doi : 10.1073/pnas.0308352100 . PMC 337057 . PMID 14745020.
^ Malik-Peiris JS, Poon LL, Guan Y (julio de 2009). "Aparición de un nuevo virus de influenza A de origen porcino (S-OIV) H1N1 en humanos". J Clin Virol . 45 (3): 169–173. doi :10.1016/j.jcv.2009.06.006. PMC 4894826 . PMID 19540800.
^ Potter CW (octubre de 2001). "Una historia de la gripe". Revista de microbiología aplicada . 91 (4): 572–9. doi :10.1046/j.1365-2672.2001.01492.x. PMID 11576290. S2CID 26392163.
^ "Diez cosas que debe saber sobre la gripe pandémica". Organización Mundial de la Salud. 14 de octubre de 2005. Archivado desde el original el 23 de septiembre de 2009. Consultado el 26 de septiembre de 2009 .
^ Valleron AJ, Cori A, Valtat S, Meurisse S, Carrat F, Boëlle PY (mayo de 2010). "Transmisibilidad y propagación geográfica de la pandemia de gripe de 1889". Proc. Natl. Sci. USA . 107 (19): 8778–81. Bibcode :2010PNAS..107.8778V. doi : 10.1073/pnas.1000886107 . PMC 2889325 . PMID 20421481.
^ Mills CE, Robins JM , Lipsitch M (diciembre de 2004). "Transmisibilidad de la gripe pandémica de 1918". Nature . 432 (7019): 904–06. Bibcode :2004Natur.432..904M. doi :10.1038/nature03063. PMC 7095078. PMID 15602562 .
^ Donaldson LJ, Rutter PD, Ellis BM, et al. (2009). "Mortalidad por influenza pandémica A/H1N1 2009 en Inglaterra: estudio de vigilancia de salud pública". BMJ . 339 : b5213. doi :10.1136/bmj.b5213. PMC 2791802 . PMID 20007665.
^ "Actualización diaria del ECDC: pandemia (H1N1) 2009, 18 de enero de 2010" (PDF) . Centro Europeo para la Prevención y el Control de Enfermedades . 18 de enero de 2010. Archivado desde el original (PDF) el 22 de enero de 2010. Consultado el 18 de enero de 2010 .
^ Dawood FS, Iuliano AD, Reed C, Meltzer MI, Shay DK, Cheng PY, Bandaranayake D, Breiman RF, Brooks WA, Buchy P, Feikin DR, Fowler KB, Gordon A, Hien NT, Horby P, Huang QS, Katz MA, Krishnan A, Lal R, Montgomery JM, Mølbak K, Pebody R, Presanis AM, Razuri H, Steens A, Tinoco YO, Wallinga J, Yu H, Vong S, Bresee J, Widdowson MA (septiembre de 2012). "Mortalidad global estimada asociada con los primeros 12 meses de circulación del virus de la influenza pandémica A H1N1 de 2009: un estudio de modelado". The Lancet. Enfermedades infecciosas (manuscrito enviado). 12 (9): 687–95. doi :10.1016/S1473-3099(12)70121-4. PMID 22738893. Archivado (PDF) del original el 9 de abril de 2024 – vía Zenodo.
^ Osterhaus AD, Rimmelzwaan GF, Martina BE, Bestebroer TM, Fouchier RA (mayo de 2000). "Virus de la influenza B en focas". Science . 288 (5468): 1051–3. Bibcode :2000Sci...288.1051O. doi :10.1126/science.288.5468.1051. PMID 10807575.
^ Nobusawa E, Sato K (abril de 2006). "Comparación de las tasas de mutación de los virus de la gripe humana A y B". Journal of Virology . 80 (7): 3675–8. doi :10.1128/JVI.80.7.3675-3678.2006. PMC 1440390 . PMID 16537638.
^ Webster RG, Bean WJ, Gorman OT, Chambers TM, Kawaoka Y (marzo de 1992). "Evolución y ecología de los virus de la influenza A". Microbiological Reviews . 56 (1): 152–79. doi :10.1128/MMBR.56.1.152-179.1992. PMC 372859 . PMID 1579108.
^ Zambon MC (noviembre de 1999). "Epidemiología y patogénesis de la gripe". The Journal of Antimicrobial Chemotherapy . 44 Suppl B (Suppl B): 3–9. doi : 10.1093/jac/44.suppl_2.3 . PMID 10877456. S2CID 15968981.
^ Matsuzaki Y, Sugawara K, Mizuta K, Tsuchiya E, Muraki Y, Hongo S, Suzuki H, Nakamura K (2002). "Caracterización antigénica y genética de los virus de la influenza C que causaron dos brotes en la ciudad de Yamagata, Japón, en 1996 y 1998". J Clin Microbiol . 40 (2): 422–29. doi :10.1128/JCM.40.2.422-429.2002. PMC 153379 . PMID 11825952.
^ Matsuzaki Y, Katsushima N, Nagai Y, Shoji M, Itagaki T, Sakamoto M, Kitaoka S, Mizuta K, Nishimura H (1 de mayo de 2006). "Características clínicas de la infección por el virus de la influenza C en niños". J Infect Dis . 193 (9): 1229–35. doi : 10.1086/502973 . PMID 16586359.
^ Katagiri S, Ohizumi A, Homma M (julio de 1983). "Un brote de influenza tipo C en un hogar de niños". J Infect Dis . 148 (1): 51–56. doi :10.1093/infdis/148.1.51. PMID 6309999.
^ Hause BM, Ducatez M, Collin EA, Ran Z, Liu R, Sheng Z, Armien A, Kaplan B, Chakravarty S, Hoppe AD, Webby RJ, Simonson RR, Li F (febrero de 2013). "Aislamiento de un nuevo virus de influenza porcina de Oklahoma en 2011 que está distantemente relacionado con los virus de influenza C humanos". PLOS Pathogens . 9 (2): e1003176. doi : 10.1371/journal.ppat.1003176 . PMC 3567177 . PMID 23408893.
^ Sheng Z, Ran Z, Wang D, Hoppe AD, Simonson R, Chakravarty S, Hause BM, Li F (febrero de 2014). "Caracterización genómica y evolutiva de un nuevo virus de tipo influenza C de cerdo". Archivos de Virología . 159 (2): 249–55. doi :10.1007/s00705-013-1815-3. PMC 5714291 . PMID 23942954.
^ Collin EA, Sheng Z, Lang Y, Ma W, Hause BM, Li F (enero de 2015). "Cocirculación de dos linajes genéticos y antigénicos distintos del virus de influenza D propuesto en el ganado". Journal of Virology . 89 (2): 1036–42. doi :10.1128/JVI.02718-14. PMC 4300623 . PMID 25355894.
^ abc Spickler AR (febrero de 2016). "Influenza" (PDF) . Centro para la Seguridad Alimentaria y la Salud Pública . Universidad Estatal de Iowa. pág. 7.
^ Suzuki Y (octubre de 2006). "Selección natural en el genoma del virus de la influenza". Biología molecular y evolución . 23 (10). Oxford Academic: 1902–11. doi : 10.1093/molbev/msl050 . PMID 16818477. Archivado desde el original el 21 de enero de 2022.
Lectura adicional
Hoyle, L. (1969). Los virus de la gripe. Monografías de virología. Vol. 4. Springer-Verlag. ISBN 978-3-211-80892-4. ISSN 0083-6591. OCLC 4053391.
Enlaces externos
Wikispecies tiene información relacionada con Orthomyxoviridae .
Portal Salud-UE: Trabajo de la UE para preparar una respuesta mundial a la gripe.
Base de datos de investigación sobre la influenza: base de datos de secuencias genómicas de la influenza e información relacionada.
Comisión Europea - Salud pública: coordinación de la UE en relación con la pandemia (H1N1) 2009
Estructuras relacionadas con el virus de la influenza en 3D del Banco de datos EM (EMDB)
Viralzone: Orthomyxoviridae
Taxonomía de virus: publicación de 2020: Comité Internacional de Taxonomía de Virus (ICTV)