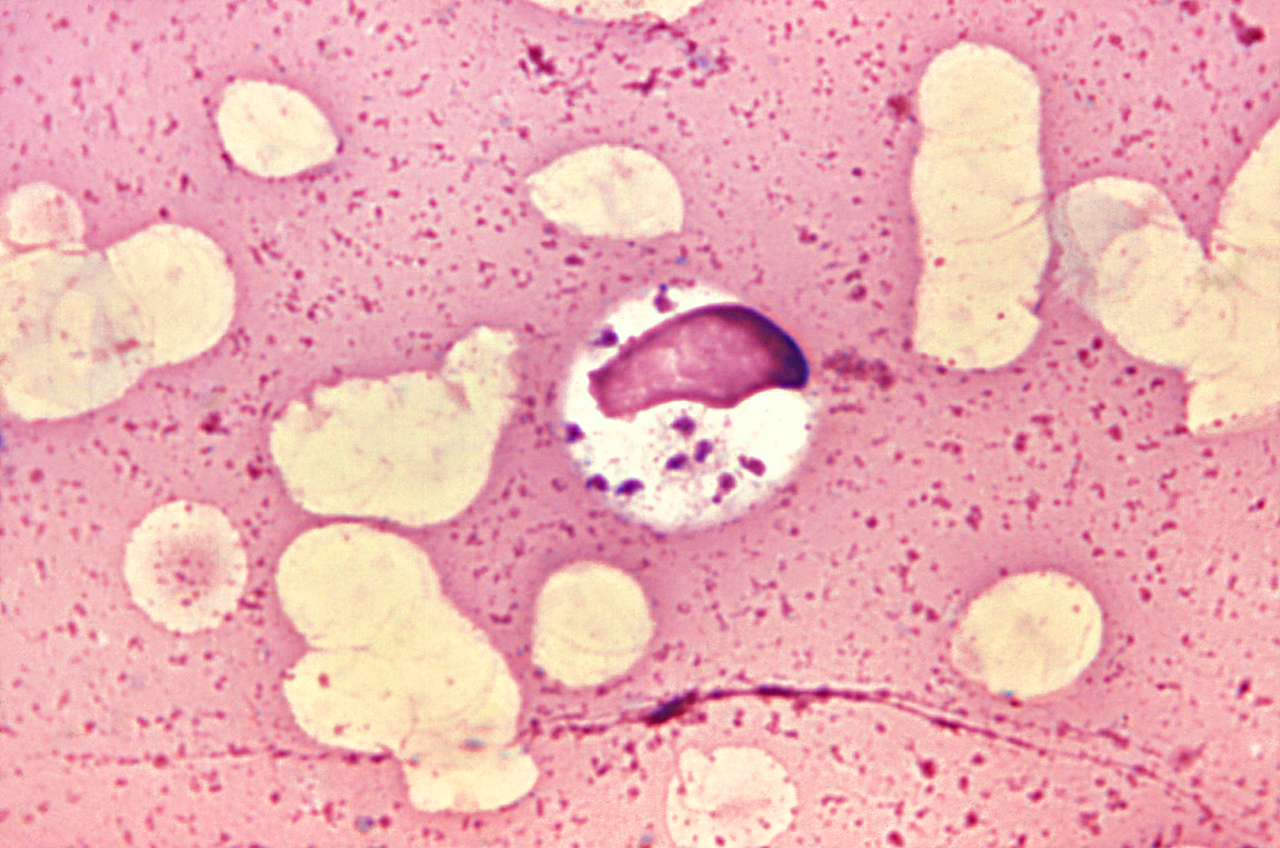
Trypanosomatida es un grupo de organismos unicelulares cinetoplastidos que se distinguen por tener un solo flagelo . El nombre se deriva del griego trypano (barrenador) y soma (cuerpo) debido al movimiento en espiral de algunas especies de tripanosomátidos. Todos los miembros son exclusivamente parásitos , se encuentran principalmente en insectos . [1] Algunos géneros tienen ciclos de vida que involucran un huésped secundario, que puede ser un vertebrado , invertebrado o planta . Estos incluyen varias especies que causan enfermedades importantes en humanos. [2] Algunos tripanosomátidos son parásitos intracelulares , con la importante excepción de Trypanosoma brucei .
Tres géneros son dixénicos (dos hospedadores en el ciclo de vida): Leishmania , Phytomonas y Trypanosoma . El resto son monoxénicos (un hospedador en el ciclo de vida). Paratrypanosoma parece ser la primera rama evolutiva de este orden. Se reconocen quince géneros en Trypanosomatidae y hay tres subfamilias: Blechomonadinae , Leishmaniinae y Strigomonadinae . Los géneros de la subfamilia Strigomonadinae se caracterizan por la presencia de bacterias intracelulares obligatorias del género Kinetoplastibacterium . [ cita requerida ]
Algunos tripanosomátidos sólo ocupan un único huésped , mientras que muchos otros son heteroxeno : viven en más de una especie huésped a lo largo de su ciclo de vida. Este ciclo de vida heteroxeno normalmente incluye el intestino de un insecto hematófago y la sangre y/o tejidos de un vertebrado . Los huéspedes más raros incluyen otros invertebrados hematófagos, como las sanguijuelas , [7] y otros organismos como las plantas . Diferentes especies pasan por una variedad de morfologías diferentes en diferentes etapas del ciclo de vida, y la mayoría tiene al menos dos morfologías diferentes. Normalmente, las formas promastigotes y epimastigotes se encuentran en huéspedes insectos, las formas tripomastigotes en el torrente sanguíneo de los mamíferos y los amastigotes en entornos intracelulares . [ cita requerida ]
Entre los ejemplos comúnmente estudiados, T. brucei , T. congolense y T. vivax son extracelulares, mientras que T. cruzi y Leishmania spp. son intracelulares. [8] Los tripanosomátidos con estadios intracelulares expresanProteínas δ-amastina en sus superficies. [8] de Paiva et al. , 2015 ilumina el papel de las δ-amastinas en el éxito intracelular. [8]
En los ciclos de vida de los tripanosomátidos aparecen diversas formas morfológicas diferentes, que se distinguen principalmente por la posición, la longitud y la unión del flagelo al cuerpo celular . El cinetoplasto se encuentra estrechamente asociado con el cuerpo basal en la base del flagelo y todas las especies de tripanosomátidos tienen un solo núcleo. La mayoría de estas morfologías se pueden encontrar como una etapa del ciclo de vida en todos los géneros de tripanosomátidos, sin embargo, ciertas morfologías son particularmente comunes en un género en particular. Las diversas morfologías se denominaban originalmente a partir del género en el que se encontraba comúnmente, aunque esta terminología ahora rara vez se usa debido a la posible confusión entre morfologías y género. La terminología moderna se deriva del griego "mastig", que significa látigo (refiriéndose al flagelo), y un prefijo que indica la ubicación del flagelo en la célula. Por ejemplo, la forma amastigote (prefijo "a-", que significa sin flagelo) también se conoce como la forma leishmanial, ya que todas las Leishmania tienen una etapa de ciclo de vida amastigote. [ cita requerida ]
Amastigote (leishmania). [10] Los amastigotes son una morfología común durante una etapa del ciclo de vida intracelular en un huésped mamífero. Todas las Leishmania tienen una etapa de amastigote en el ciclo de vida. Los amastigotes de Leishmania son particularmente pequeños y se encuentran entre las células eucariotas más pequeñas. El flagelo es muy corto y se proyecta solo ligeramente más allá del bolsillo flagelar.
Promastigote (leptomonad).[10]La forma promastigote es una morfología común en el insecto huésped. El flagelo se encuentra delante del núcleo y emerge directamente del cuerpo celular anterior. El cinetoplasto se encuentra delante del núcleo, cerca del extremo anterior del cuerpo.
Epimastigotes (crithidial).[10]Los epimastigotes son una forma común en el huésped insecto y Crithidia y Blastocrithidia , ambos parásitos de insectos, exhiben esta forma durante sus ciclos de vida. El flagelo sale de la célula por delante del núcleo y está conectado al cuerpo celular durante parte de su longitud por una membrana ondulada. El cinetoplasto está ubicado entre el núcleo y el extremo anterior.
Tripomastigote (tripanosoma).[10]Esta etapa es característica del género Trypanosoma en el torrente sanguíneo del huésped mamífero, así como de las etapas metacíclicas infecciosas en el vector, la mosca. En los tripomastigotes, elcinetoplastose encuentra cerca del extremo posterior del cuerpo y el flagelo se encuentra unido al cuerpo celular en la mayor parte de su longitud mediante una membrana ondulada.
Opistomastigote (herpetomonad).[10]Una morfología más rara donde el flagelo se encuentra detrás del núcleo, pasando a través de un surco largo en la célula.
Endomastigote .[11]Un morfotipo en el que el flagelo no se extiende más allá del bolsillo flagelar profundo.
Amastigote : Micrografía SEM en falso color de la forma amastigote de Leishmania mexicana . El cuerpo celular se muestra en naranja y el flagelo en rojo. 219 píxeles/μm.
Promastigote : Micrografía SEM en falso color de la forma promastigote de Leishmania mexicana . El cuerpo celular se muestra en naranja y el flagelo en rojo. 119 píxeles/μm.
Tripomastigote : micrografía SEM en falso color de la forma procíclica Trypanosoma brucei . El cuerpo celular se muestra en naranja y el flagelo en rojo. 84 píxeles/μm.
Otras características
Las características notables de los tripanosomátidos son la capacidad de realizar trans-splicing de ARN y la posesión de glicosomas , donde se limita gran parte de su glicólisis . El acidocalcisoma , otro orgánulo , fue identificado por primera vez en los tripanosomas. [12]
Endosimbionte bacteriano
Se sabe que seis especies de tripanosomátidos portan un endosimbionte proteobacteriano adicional , denominado TPE (endosimbiontes proteobacterianos tripanosomátidos). Estos tripanosomátidos ( Strigomonas oncopelti , S. culicis , S. galati , Angomonas desouzai y A. deanei ) se conocen a su vez como SHT, por tripanosomátidos que albergan simbiontes. Todos estos simbiontes tienen un origen evolutivo compartido y se clasifican en el género Candidatus " Kinetoplastibacterium ". [13]
Al igual que con muchos simbiontes, las bacterias tienen un genoma mucho más reducido en comparación con sus parientes de vida libre de los géneros Taylorella y Achromobacter . ( GTDB encuentra al género hermano de Proftella , un simbionte de Diaphorina citri ). [14] Como reflejo de su incapacidad para vivir solas, han perdido genes dedicados a funciones biológicas esenciales, dependiendo en su lugar del huésped. Han modificado su división para sincronizarse con el huésped. Al menos en S. culicis , el TPE ayuda al huésped sintetizando hemo [13] y produciendo enzimas esenciales, permaneciendo atado al cinetoplasto . [15]
Referencias
^ Podlipaev S (mayo de 2001). "Cuantos más insectos tripanosomátidos se estudian, más diversa parece la especie Trypanosomatidae". Revista Internacional de Parasitología . 31 (5–6): 648–52. doi :10.1016/S0020-7519(01)00139-4. PMID 11334958.
^ Simpson AG, Stevens JR, Lukes J (abril de 2006). "La evolución y diversidad de los flagelados cinetoplastídeos". Tendencias en parasitología . 22 (4): 168–74. doi :10.1016/j.pt.2006.02.006. PMID 16504583.
^ "Trypanosomiasis africana humana (enfermedad del sueño)". www.who.int . Archivado desde el original el 20 de abril de 2018 . Consultado el 14 de mayo de 2020 .
^
Rêgo, Felipe D.; Soares, Rodrigo Pedro (2021). "Lutzomyia longipalpis: una actualización sobre este vector del flebótomo". Anais da Academia Brasileira de Ciências . 93 (3): e20200254. doi : 10.1590/0001-37652021XXXX . hdl : 11336/166702 . PMID 33950136. S2CID 233743387. e20200254.
Esta revisión cita esta investigación.
Abbasi, Ibrahim; Trancoso Lopo de Queiroz, Artur; Kirstein, Óscar David; Nasereddin, Abdelmajeed; Horwitz, Ben Sión; Hailu, Asrat; Salah, Ikram; Mota, Tiago Feitosa; Fraga, Deborah Bittencourt Mothé (13 de noviembre de 2018). "Los flebótomos que se alimentan de plantas, vectores de la leishmaniasis, prefieren el Cannabis sativa". Actas de la Academia Nacional de Ciencias de los Estados Unidos de América . 115 (46): 11790–11795. Código Bib : 2018PNAS..11511790A. doi : 10.1073/pnas.1810435115 . PMC 6243281 . Número de modelo: PMID 30373823. Número de modelo: S2CID 53112660.
^ Poinar, G. (2008). "Lutzomyia adiketis sp. n. (Diptera: Phlebotomidae), un vector de Paleoleishmania neotropicum sp. n. (Kinetoplastida: Trypanosomatidae) en ámbar dominicano". Parásitos y vectores . 1 (1): 22. doi : 10.1186/1756-3305-1-22 . PMC 2491605 . PMID 18627624.
^ Poinar, G. (2005). " Triatoma dominicana sp. n. (Hemiptera: Reduviidae: Triatominae) y Trypanosoma antiquus sp. n. (Stercoraria: Trypanosomatidae), la primera evidencia fósil de una asociación entre vectores triatominos y tripanosomátidos". Enfermedades transmitidas por vectores y zoonóticas . 5 (1): 72–81. doi :10.1089/vbz.2005.5.72. PMID 15815152.
^ Hamilton, PB; Stevens, JR; Gidley, J.; Holz, P.; Gibson, WC (abril de 2005). "Un nuevo linaje de tripanosomas de vertebrados australianos y sanguijuelas hematófagas terrestres (Haemadipsidae)". Revista Internacional de Parasitología . 35 (4): 431–443. doi :10.1016/j.ijpara.2004.12.005. PMID 15777919.
^ abc Silva Pereira, Sara; Trindade, Sandra; De Niz, Mariana; Figueiredo, Luisa M. (2019). "Tropismo tisular en enfermedades parasitarias". Biología abierta . 9 (5): 190036. doi :10.1098/rsob.190036. PMC 6544988 . PMID 31088251.
^ ab Silva, Verônica Santana da; Machado, Carlos Renato (2022). "Sexo en protistos: una nueva perspectiva sobre los mecanismos de reproducción de los tripanosomátidos". Genética y Biología Molecular . 45 (3): e20220065. doi :10.1590/1678-4685-GMB-2022-0065. PMC 9552303 . PMID 36218381.
^ abcde Hoare, Cecil A.; Wallace, Franklin G. (1966). «Etapas de desarrollo de flagelados tripanosomátidos: una nueva terminología». Nature . 212 (5068): 1385–6. Código Bibliográfico :1966Natur.212.1385H. doi :10.1038/2121385a0. S2CID 4164112.
^ Merzlyak, Ekaterina; Yurchenko, Vyacheslav; Kolesnikov, Alexander A.; Alexandrov, Kirill; Podlipaev, Sergei A.; Maslov, Dmitri A. (1 de marzo de 2001). "Diversidad y filogenia de los tripanosomátidos de insectos basada en genes de ARNr de subunidades pequeñas: polifilia de Leptomonas y Blastocrithidia ". The Journal of Eukaryotic Microbiology . 48 (2): 161–169. doi :10.1111/j.1550-7408.2001.tb00298.x. PMID 12095103. S2CID 13880469.
^ Docampo R, de Souza W, Miranda K, Rohloff P, Moreno SN (marzo de 2005). "Acidocalcisomas: conservados desde las bacterias hasta el hombre". Nature Reviews Microbiology . 3 (3): 251–61. doi :10.1038/nrmicro1097. PMID 15738951. S2CID 31935658.
^ ab Alves, JM; Serrano, MG; Maia da Silva, F; Voegtly, LJ; Matveyev, AV; Teixeira, MM; Camargo, EP; Buck, GA (2013). "Evolución del genoma y análisis filogenómico de Candidatus Kinetoplastibacterium, los endosimbiontes betaproteobacterianos de Strigomonas y Angomonas". Genome Biology and Evolution . 5 (2): 338–50. doi : 10.1093/gbe/evt012 . PMC 3590767 . PMID 23345457.
^ "GTDB - Árbol en g__Kinetoplastibacterium". gtdb.ecogenomic.org . Archivado desde el original el 2022-12-20 . Consultado el 2022-12-20 .
^ de Souza, W.; Motta, MC (1999). "Endosimbiosis en protozoos de la familia Trypanosomatidae". FEMS Microbiology Letters . 173 (1): 1–8. doi : 10.1111/j.1574-6968.1999.tb13477.x . PMID 10220875.
Bütikofer P, Ruepp S, Boschung M, Roditi I (septiembre de 1997). "La prociclina 'GPEET' es la principal proteína de superficie de las formas de cultivo procíclicas de la cepa 427 de Trypanosoma brucei brucei". Revista bioquímica . 326 (Pt 2): 415–23. doi :10.1042/bj3260415. PMC 1218686 . PMID 9291113.
Dean S, Marchetti R, Kirk K, Matthews KR (mayo de 2009). "Una familia de transportadores de superficie transmite la señal de diferenciación de los tripanosomas". Nature . 459 (7244): 213–7. Bibcode :2009Natur.459..213D. doi :10.1038/nature07997. PMC 2685892 . PMID 19444208.
Engstler M, Boshart M; Boshart (noviembre de 2004). "El choque frío y la regulación del tráfico de proteínas de superficie transmiten sensibilización a los inductores de la diferenciación de etapas en Trypanosoma brucei". Genes & Development . 18 (22): 2798–811. doi :10.1101/gad.323404. PMC 528899 . PMID 15545633.
Hofer A, Steverding D, Chabes A, Brun R, Thelander L (mayo de 2001). "CTP sintetasa de Trypanosoma brucei: un objetivo para el tratamiento de la enfermedad del sueño africana". Actas de la Academia Nacional de Ciencias de los Estados Unidos de América . 98 (11): 6412–6. Bibcode :2001PNAS...98.6412H. doi : 10.1073/pnas.111139498 . PMC 33482 . PMID 11353848.
Janovy, J; Roberts, LS (2005). Fundamentos de parasitología (7.ª ed.). Nueva York, NY : McGraw Hill . Págs. 61–69.
Legros D, Ollivier G, Gastellu-Etchegorry M, et al. (julio de 2002). "Tratamiento de la tripanosomiasis africana humana: situación actual y necesidades de investigación y desarrollo". Lancet Infectious Diseases . 2 (7): 437–40. doi :10.1016/S1473-3099(02)00321-3. hdl : 10144/18268 . PMID 12127356.
Matthews KR (enero de 2005). "La biología celular del desarrollo de Trypanosoma brucei". Journal of Cell Science . 118 (Pt 2): 283–90. doi :10.1242/jcs.01649. PMC 2686837 . PMID 15654017.
Matthews KR, Gull K; Gull (junio de 1994). "Evidencia de una interacción entre la progresión del ciclo celular y el inicio de la diferenciación entre las formas del ciclo de vida de los tripanosomas africanos". Journal of Cell Biology . 125 (5): 1147–56. doi :10.1083/jcb.125.5.1147. PMC 2120053 . PMID 8195296.
Morrison LJ, Marcello L, McCulloch R (diciembre de 2009). "Variación antigénica en el tripanosoma africano: mecanismos moleculares y complejidad fenotípica" (PDF) . Microbiología celular . 11 (12): 1724–34. doi :10.1111/j.1462-5822.2009.01383.x. PMID 19751359. S2CID 26552797. Archivado (PDF) desde el original el 2021-10-20 . Consultado el 2019-07-10 .
Seed JR, Wenck MA; Wenck (junio de 2003). "Papel de la transición de largo delgado a corto rechoncho en el ciclo de vida de los tripanosomas africanos". Kinetoplastid Biology and Disease . 2 (1): 3. doi : 10.1186/1475-9292-2-3 . PMC 165594 . PMID 12844365.
Shadan S (mayo de 2009). "Microbiología: señales de cambio". Nature . 459 (7244): 175. Bibcode :2009Natur.459..175S. doi : 10.1038/459175a . PMID 19444199.
Sherwin T, Gull K; Gull (junio de 1989). "El ciclo de división celular de Trypanosoma brucei brucei : cronometraje de los marcadores de eventos y modulaciones del citoesqueleto". Philosophical Transactions of the Royal Society B . 323 (1218): 573–88. Bibcode :1989RSPTB.323..573S. doi :10.1098/rstb.1989.0037. PMID 2568647.
«Tripanosomiasis africana». Organización Mundial de la Salud . Agosto de 2006. Archivado desde el original el 4 de diciembre de 2016. Consultado el 5 de octubre de 2020 .
Woodcock, Harold Mellor (1911). "Trypanosomes" (Tripanosomas) . En Chisholm, Hugh (ed.). Encyclopædia Britannica . Vol. 27 (11.ª ed.). Cambridge University Press. págs. 340–347.(en línea) Un estudio exhaustivo de la historia natural de los organismos.
Enlaces externos
Wikimedia Commons alberga una categoría multimedia sobre Trypanosomatida .
Trykipedia, ontologías específicas de tripanosomátidos
_Trypanosoma_equiperdum.jpg/1280px-Trypanosoma_(248_09)_Trypanosoma_equiperdum.jpg)