Un osteoclasto (del griego antiguo ὀστέον (osteon) 'hueso' y κλαστός (clastos) 'roto') es un tipo de célula ósea que descompone el tejido óseo . Esta función es fundamental en el mantenimiento, la reparación y la remodelación de los huesos del esqueleto vertebral . El osteoclasto desmonta y digiere el compuesto de proteína hidratada y mineral a nivel molecular mediante la secreción de ácido y una colagenasa , un proceso conocido como resorción ósea . Este proceso también ayuda a regular el nivel de calcio en sangre .
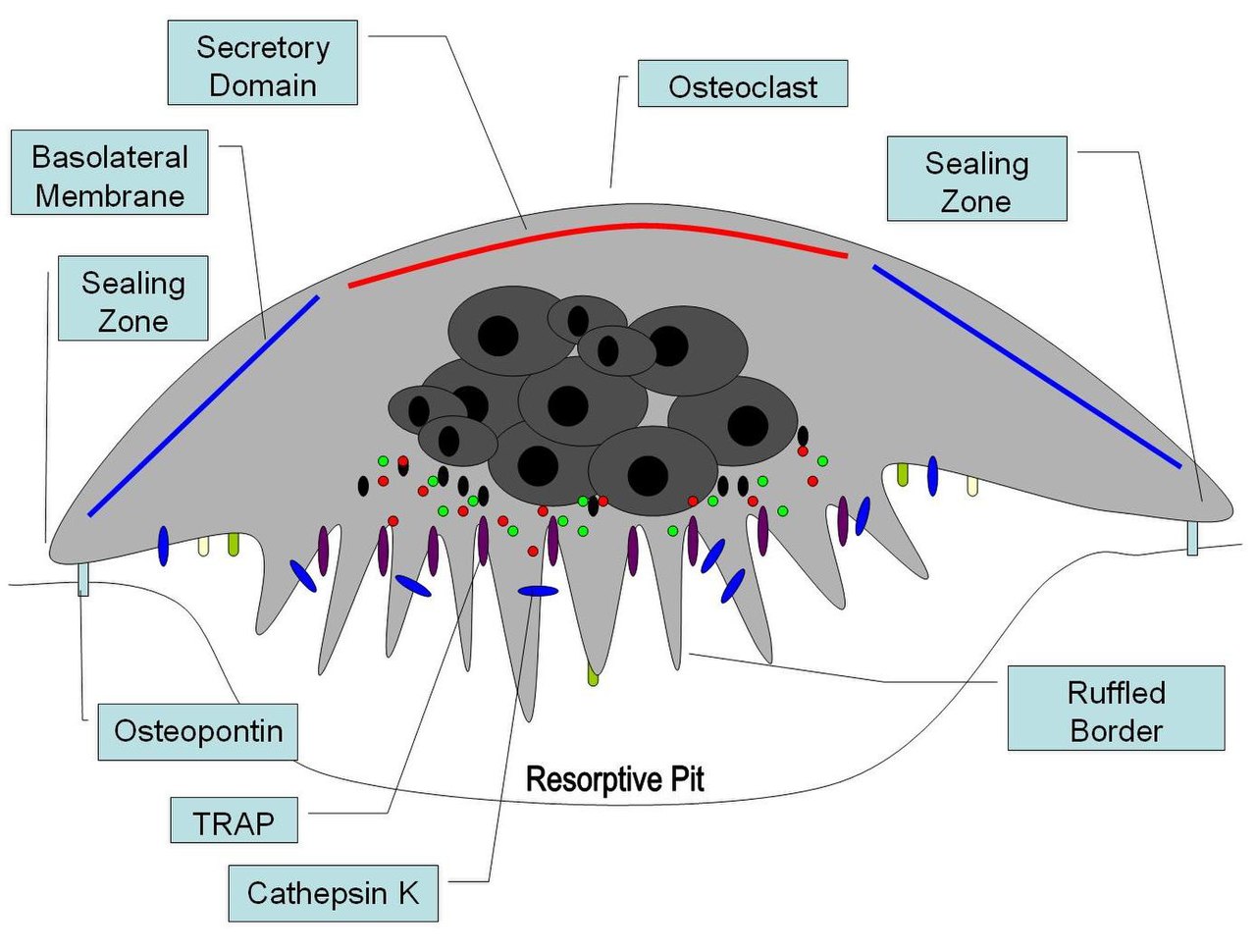
Los osteoclastos se encuentran en las superficies del hueso que están sufriendo reabsorción. En dichas superficies, los osteoclastos se encuentran en depresiones poco profundas llamadas bahías de reabsorción (lagunas de Howship) . Las bahías de reabsorción se crean por la acción erosiva de los osteoclastos sobre el hueso subyacente. El borde de la parte inferior de un osteoclasto exhibe procesos similares a dedos debido a la presencia de pliegues profundos de la membrana celular ; este borde se llama borde ondulado . El borde ondulado se encuentra en contacto con la superficie del hueso dentro de una bahía de reabsorción. La periferia del borde ondulado está rodeada por una zona de citoplasma en forma de anillo que está desprovista de orgánulos celulares pero es rica en filamentos de actina . Esta zona se llama zona clara o zona de sellado . Los filamentos de actina permiten que la membrana celular que rodea la zona de sellado se ancle firmemente a la pared ósea de las lagunas de Howship. De esta manera, se crea un compartimento subosteoclástico cerrado entre el borde ondulado y el hueso que está sufriendo reabsorción. Los osteoclastos secretan iones de hidrógeno , colagenasa , catepsina K y enzimas hidrolíticas en este compartimento. La reabsorción de la matriz ósea por los osteoclastos implica dos pasos: (1) disolución de componentes inorgánicos (minerales), y (2) digestión del componente orgánico de la matriz ósea. Los osteoclastos bombean iones de hidrógeno al compartimento subosteoclástico y crean así un microambiente ácido, que aumenta la solubilidad del mineral óseo, lo que da como resultado la liberación y reingreso de minerales óseos en el citoplasma de los osteoclastos para ser entregados a los capilares cercanos. Después de la eliminación de minerales, la colagenasa y la gelatinasa se secretan en el compartimento subosteoclástico. Estas enzimas digieren y degradan el colágeno y otros componentes orgánicos de la matriz ósea descalcificada. Los productos de degradación son fagocitados por los osteoclastos en el borde ondulado. Debido a sus propiedades fagocíticas, los osteoclastos se consideran un componente del sistema fagocítico mononuclear (SFM). La actividad de los osteoclastos está controlada por hormonas y citocinas. La calcitonina, una hormona de la glándula tiroides, suprime la actividad osteoclástica. Los osteoclastos no tienen receptores para la hormona paratiroidea (PTH). Sin embargo, la PTH estimula a los osteoblastos para que secreten la citocina denominada factor estimulante de osteoclastos, que es un potente estimulador de la actividad osteoclástica. [1]
Un odontoclasto (o-don´to-klast) es un osteoclasto asociado con la absorción de las raíces de los dientes deciduos . [2] [3] [4]

Un osteoclasto es una célula multinucleada de gran tamaño y los osteoclastos humanos en el hueso suelen tener cuatro núcleos [5] y un diámetro de entre 150 y 200 μm. Cuando se utilizan citocinas inductoras de osteoclastos para convertir los macrófagos en osteoclastos, se forman células muy grandes que pueden alcanzar los 100 μm de diámetro. Estas pueden tener docenas de núcleos y, por lo general, expresan las principales proteínas de los osteoclastos, pero tienen diferencias significativas con respecto a las células del hueso vivo debido al sustrato no natural. [6] [7] El tamaño del osteoclasto multinucleado ensamblado le permite concentrar las capacidades de transporte de iones, secreción de proteínas y transporte vesicular de muchos macrófagos en un área localizada del hueso.
En el hueso, los osteoclastos se encuentran en fosas en la superficie ósea que se denominan bahías de reabsorción o lagunas de Howship . Los osteoclastos se caracterizan por un citoplasma con un aspecto homogéneo y "espumoso". Este aspecto se debe a una alta concentración de vesículas y vacuolas . Estas vacuolas incluyen lisosomas llenos de fosfatasa ácida . Esto permite la caracterización de los osteoclastos mediante su tinción para una alta expresión de fosfatasa ácida resistente al tartrato (TRAP) y catepsina K. El retículo endoplasmático rugoso de los osteoclastos es escaso y el complejo de Golgi es extenso. [8] [9] [10]
En un sitio de resorción ósea activa, el osteoclasto forma una membrana celular especializada , el "borde ondulado", que se opone a la superficie del tejido óseo. Este borde muy plegado o ondulado facilita la eliminación ósea al aumentar drásticamente la superficie celular para la secreción y absorción del contenido del compartimento de resorción y es una característica morfológica de un osteoclasto que está reabsorbiendo hueso de forma activa.
Desde su descubrimiento en 1873 ha habido un considerable debate sobre su origen. Tres teorías fueron dominantes: de 1949 a 1970 fue popular la del origen del tejido conectivo, que afirmaba que los osteoclastos y los osteoblastos son del mismo linaje, y los osteoblastos se fusionan para formar osteoclastos. Después de años de controversia, ahora está claro que estas células se desarrollan a partir de la autofusión de macrófagos. [11] Fue a principios de 1980 cuando se reconoció al sistema fagocítico de monocitos como precursor de los osteoclastos. [12] La formación de osteoclastos requiere la presencia de RANKL (ligando del activador del receptor del factor nuclear κβ) y M-CSF (factor estimulante de colonias de macrófagos) . Estas proteínas unidas a la membrana son producidas por células estromales vecinas y osteoblastos , por lo que se requiere un contacto directo entre estas células y los precursores de osteoclastos .
El M-CSF actúa a través de su receptor en el osteoclasto, c-fms ( receptor del factor estimulante de colonias 1 ), un receptor transmembrana de la tirosina quinasa , lo que lleva a la activación del mensajero secundario de la tirosina quinasa Src. Ambas moléculas son necesarias para la osteoclastogénesis y están ampliamente involucradas en la diferenciación de células derivadas de monocitos/macrófagos.
RANKL es un miembro de la familia de necrosis tumoral ( TNF ), y es esencial en la osteoclastogénesis. Los ratones knock out de RANKL exhiben un fenotipo de osteopetrosis y defectos de erupción dental, junto con una ausencia o deficiencia de osteoclastos. RANKL activa NF-κβ (factor nuclear-κβ) y NFATc1 (factor nuclear de células T activadas, citoplasmático, dependiente de calcineurina 1) a través de RANK . La activación de NF-κβ se estimula casi inmediatamente después de que ocurre la interacción RANKL-RANK y no se regula al alza. Sin embargo, la estimulación de NFATc1 comienza ~24–48 horas después de que ocurre la unión y se ha demostrado que su expresión depende de RANKL.
La diferenciación de los osteoclastos se ve inhibida por la osteoprotegerina (OPG), que es producida por los osteoblastos y se une a RANKL, impidiendo así la interacción con RANK. Mientras que los osteoclastos derivan del linaje hematopoyético, los osteoblastos derivan de células madre mesenquimales. [13] [14]
Una vez activados, los osteoclastos se desplazan a las zonas de microfractura en el hueso mediante quimiotaxis . Los osteoclastos se encuentran en pequeñas cavidades llamadas lagunas de Howship, formadas a partir de la digestión del hueso subyacente. La zona de sellado es la unión de la membrana plasmática del osteoclasto al hueso subyacente. Las zonas de sellado están delimitadas por cinturones de estructuras de adhesión especializadas llamadas podosomas . La unión a la matriz ósea se ve facilitada por los receptores de integrina, como αvβ3, a través del motivo de aminoácidos específico Arg-Gly-Asp en las proteínas de la matriz ósea, como la osteopontina . El osteoclasto libera iones de hidrógeno a través de la acción de la anhidrasa carbónica ( H 2 O + CO 2 → HCO 3 − + H + ) a través del borde ondulado hacia la cavidad de reabsorción, acidificando y ayudando a la disolución de la matriz ósea mineralizada en Ca 2+ , H 3 PO 4 , H 2 CO 3 , agua y otras sustancias. Se ha documentado que la disfunción de la anhidrasa carbónica causa algunas formas de osteopetrosis. Los iones de hidrógeno son bombeados contra un alto gradiente de concentración por bombas de protones , específicamente una ATPasa vacuolar única . Esta enzima ha sido el objetivo en la prevención de la osteoporosis . Además, varias enzimas hidrolíticas , como los miembros de los grupos de catepsina y metaloproteasa de matriz (MMP), se liberan para digerir los componentes orgánicos de la matriz. Estas enzimas son liberadas en el compartimento por los lisosomas . De estas enzimas hidrolíticas, la catepsina K es la más importante.
La catepsina K es una proteasa de cisteína colagenolítica similar a la papaína que se expresa principalmente en los osteoclastos y se secreta en la fosa de reabsorción. La catepsina K es la proteasa principal implicada en la degradación del colágeno tipo I y otras proteínas no colágenas. Las mutaciones en el gen de la catepsina K están asociadas con la picnodisostosis , una enfermedad osteopetrótica hereditaria , caracterizada por una falta de expresión funcional de la catepsina K. Los estudios de eliminación de la catepsina K en ratones conducen a un fenotipo osteopetrótico, que se compensa parcialmente con una mayor expresión de proteasas distintas de la catepsina K y una osteoclastogénesis mejorada.
La catepsina K tiene una actividad enzimática óptima en condiciones ácidas. Se sintetiza como una proenzima con un peso molecular de 37 kDa y, tras la activación por escisión autocatalítica, se transforma en la forma madura y activa con un peso molecular de ~27 kDa.
Tras la polarización del osteoclasto sobre el sitio de resorción, la catepsina K se secreta desde el borde ondulado hacia la fosa de resorción. La catepsina K transmigra a través del borde ondulado mediante vesículas intercelulares y luego es liberada por el dominio secretor funcional . Dentro de estas vesículas intercelulares, la catepsina K, junto con las especies reactivas de oxígeno generadas por TRAP , degrada aún más la matriz extracelular ósea.
Varias otras catepsinas se expresan en los osteoclastos, incluidas las catepsinas B , C , D , E , G y L. La función de estas proteasas de cisteína y aspártico es generalmente desconocida dentro del hueso, y se expresan en niveles mucho más bajos que la catepsina K.
Los estudios realizados en ratones knock out para catepsina L han sido mixtos: hay un informe de hueso trabecular reducido en ratones knock out para catepsina L homocigotos y heterocigotos en comparación con los de tipo salvaje y otro informe que no encontró anomalías esqueléticas.
Las metaloproteinasas de matriz (MMP) comprenden una familia de más de 20 endopeptidasas dependientes del zinc. El papel de las metaloproteinasas de matriz (MMP) en la biología de los osteoclastos no está bien definido, pero en otros tejidos se las ha vinculado con actividades promotoras de tumores, como la activación de factores de crecimiento , y son necesarias para la metástasis tumoral y la angiogénesis.
La MMP9 está asociada con el microambiente óseo. La expresan los osteoclastos y se sabe que es necesaria para la migración de los osteoclastos y es una gelatinasa potente. Los ratones transgénicos que carecen de MMP-9 desarrollan defectos en el desarrollo óseo, la angiogénesis intraósea y la reparación de fracturas.
Se cree que la MMP-13 está involucrada en la resorción ósea y en la diferenciación de los osteoclastos, ya que los ratones knock-out revelaron una disminución en el número de osteoclastos, osteopetrosis y una disminución de la resorción ósea.
Las MMP expresadas por el osteoclasto incluyen MMP-9, -10, -12 y -14. Aparte de MMP-9, se sabe poco sobre su relevancia para el osteoclasto, sin embargo, se encuentran altos niveles de MMP-14 en la zona de sellado.
En los años 1980 y 1990 se estudió en detalle la fisiología de los osteoclastos típicos. Con el aislamiento del borde ondulado, se estudió directamente el transporte de iones a través de él en detalle bioquímico. Se verificó el transporte de ácido dependiente de la energía y se purificó la bomba de protones postulada. [15] [16] Con el cultivo exitoso de osteoclastos, se hizo evidente que están organizados para apoyar el transporte masivo de protones para la acidificación del compartimento de resorción y la solubilización del mineral óseo. Esto incluye la permeabilidad al Cl − del borde ondulado para controlar el potencial de membrana y el intercambio basolateral de Cl − /HCO 3 − para mantener el pH citosólico en rangos fisiológicamente aceptables. [17] [18] [19]
La eficacia de su secreción iónica depende de que el osteoclasto forme un sello eficaz alrededor del compartimento de resorción. La posición de esta "zona de sellado" parece estar mediada por integrinas expresadas en la superficie del osteoclasto. [20] Una vez establecida la zona de sellado, el osteoclasto multinucleado se reorganiza. El desarrollo de la membrana rizada altamente invaginada que se opone al compartimento de resorción permite una actividad secretora masiva. Además, permite la transcitosis vesicular del colágeno mineral y degradado desde el borde rizado hasta la membrana libre de la célula, y su liberación al compartimento extracelular. [21] [22] Esta actividad completa la resorción ósea, y tanto los componentes minerales como los fragmentos de colágeno se liberan a la circulación general.
Los osteoclastos están regulados por varias hormonas , entre ellas la hormona paratiroidea (PTH) de la glándula paratiroides, la calcitonina de la glándula tiroides y el factor de crecimiento interleucina 6 (IL-6). Esta última hormona, IL-6 , es uno de los factores de la enfermedad osteoporosis . La osteoporosis se produce cuando hay un desequilibrio entre las actividades de resorción ósea de los osteoclastos y las actividades de formación ósea de los osteoblastos. [23]
La actividad de los osteoclastos también está mediada por la interacción de dos moléculas producidas por los osteoblastos, a saber, la osteoprotegerina y el ligando RANK . Estas moléculas también regulan la diferenciación de los osteoclastos. [24] [25]
Un odontoclasto es un osteoclasto asociado con la absorción de las raíces de los dientes deciduos . [2] [3] [4]
Un osteoclasto también puede ser un instrumento utilizado para fracturar y restablecer los huesos (el origen es del griego osteon : hueso y klastos : roto). Para evitar confusiones, la célula se denominó originalmente osotoclasto. Cuando el instrumento quirúrgico dejó de utilizarse, la célula pasó a conocerse con su nombre actual.
Los osteoclastos gigantes pueden aparecer en algunas enfermedades, incluida la enfermedad ósea de Paget y la toxicidad por bisfosfonatos .
En los gatos, la actividad anormal de los odontoclastos puede causar lesiones reabsortivas odontoclásticas felinas , lo que requiere la extracción de los dientes afectados.
Los osteoclastos desempeñan un papel importante en el movimiento dentario ortodóncico y en la migración patológica de dientes periodontalmente comprometidos.
Los osteoclastos fueron descubiertos por Kölliker en 1873. [12]