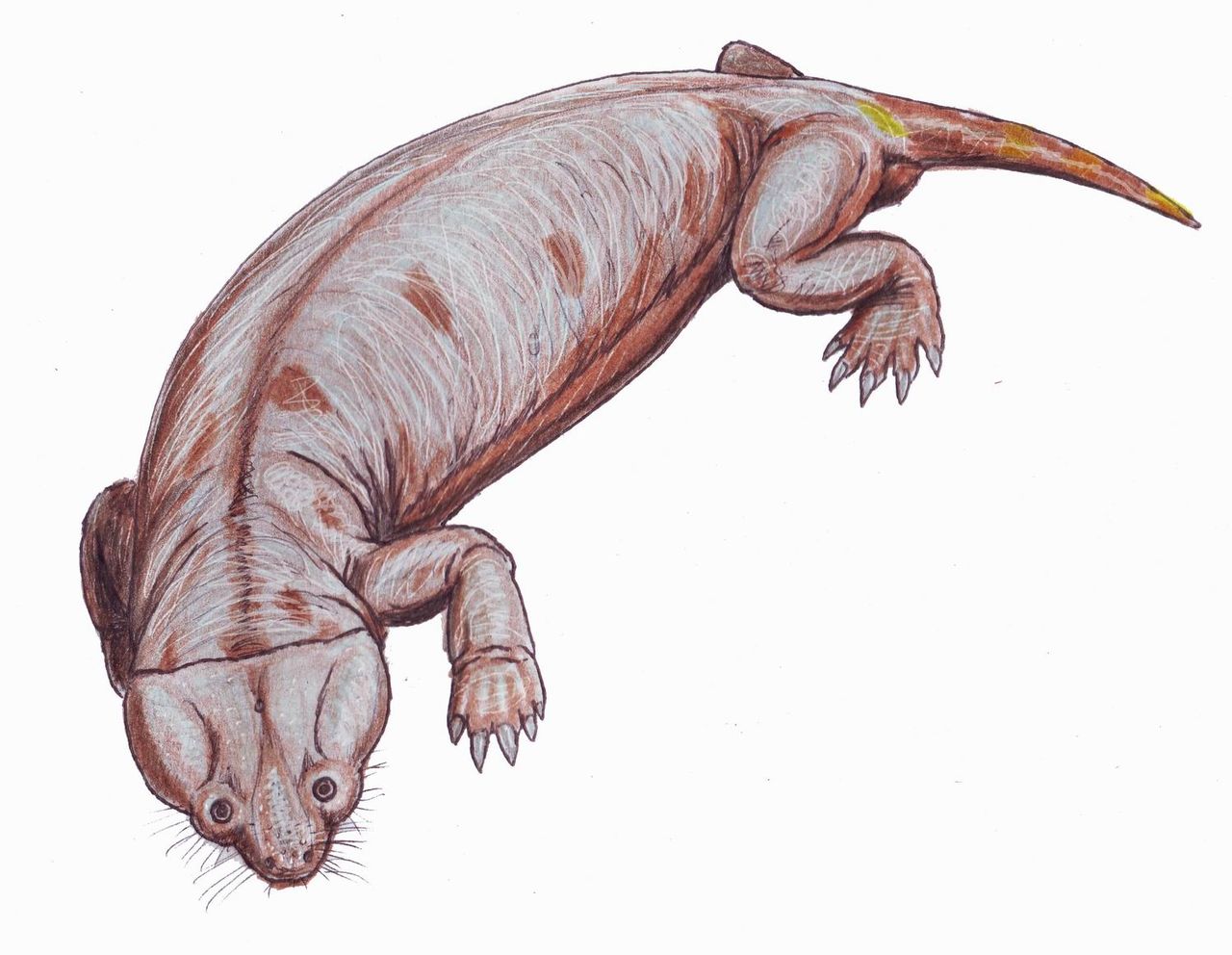
Dicynodontia es un clado extinto de anomodontos , un tipo extinto de terápsidos no mamíferos . Los dicinodontos eran herbívoros que normalmente tenían un par de colmillos, de ahí su nombre, que significa 'dos dientes de perro'. Los miembros del grupo poseían un pico córneo, normalmente sin dientes, único entre todos los sinápsidos . Los dicinodontos aparecieron por primera vez en el sur de Pangea durante el Pérmico medio , hace unos 270-260 millones de años, y se distribuyeron globalmente y se convirtieron en los animales herbívoros dominantes en el Pérmico tardío , hace unos 260-252 millones de años. Fueron devastados por la extinción masiva del final del Pérmico que acabó con la mayoría de los demás terápsidos hace unos 252 millones de años. Se recuperaron durante el Triásico, pero se extinguieron hacia el final de ese período. Fueron los terápsidos no mamíferos más exitosos y diversos, con más de 70 géneros conocidos, que variaban desde animales excavadores del tamaño de una rata hasta animales ramoneadores del tamaño de un elefante .


El cráneo de los dicinodontes es altamente especializado, ligero pero fuerte, con las aberturas temporales sinápsidas en la parte posterior del cráneo muy agrandadas para acomodar músculos mandibulares más grandes. La parte frontal del cráneo y la mandíbula inferior son generalmente estrechas y, en todas las formas primitivas, sin dientes. En cambio, la parte frontal de la boca está equipada con un pico córneo, como en las tortugas y los dinosaurios ceratopsianos . La comida se procesaba mediante la retracción de la mandíbula inferior cuando la boca se cerraba, produciendo una poderosa acción de corte, [2] que habría permitido a los dicinodontes lidiar con material vegetal duro. Los dicinodontes típicamente tenían un par de dientes caniniformes maxilares agrandados, análogos a los colmillos presentes en algunos mamíferos actuales. En los primeros géneros, eran simplemente dientes agrandados, pero en formas posteriores evolucionaron de forma independiente en dientes de crecimiento constante como los colmillos de los mamíferos varias veces. [3] En algunos dicinodontes, se ha sugerido que la presencia de colmillos es sexualmente dimórfica . [4] Algunos dicinodontes como Stahleckeria carecían de colmillos verdaderos y en su lugar tenían extensiones similares a colmillos en el costado del pico. [5] [6] : 139
El cuerpo es corto, fuerte y con forma de barril, con miembros fuertes. En los géneros grandes (como Dinodontosaurus ) las extremidades traseras se mantenían erguidas, pero las delanteras se doblaban a la altura del codo. Tanto la cintura escapular como el íleon son grandes y fuertes. La cola es corta. [ cita requerida ]
Las huellas de dicinodonte de Pentasauropus sugieren que estos animales tenían almohadillas carnosas en sus patas. [7] La piel momificada de especímenes de Lystrosaurus en Sudáfrica presenta numerosas protuberancias elevadas. [8]
Desde hace tiempo se ha sospechado que los dicinodontes son animales de sangre caliente . Sus huesos están muy vascularizados y poseen canales de Havers , y sus proporciones corporales favorecen la conservación del calor. [9] En los ejemplares jóvenes, los huesos están tan vascularizados que presentan densidades de canales más altas que la mayoría de los demás terápsidos. [10] Sin embargo, los estudios sobre coprolitos de dicinodontes del Triásico Tardío muestran, paradójicamente, patrones digestivos más típicos de animales con metabolismos lentos. [11]
Más recientemente, el descubrimiento de restos de pelo en coprolitos del Pérmico posiblemente reivindique el estatus de los dicinodontes como animales endotérmicos. Como estos coprolitos provienen de especies carnívoras y los huesos digeridos de dicinodontes son abundantes, se ha sugerido que al menos algunos de estos restos de pelo provienen de presas de dicinodontes. [12] Un nuevo estudio que utilizó análisis químicos pareció sugerir que tanto los cinodontes como los dicinodontes desarrollaron sangre caliente de forma independiente antes de la extinción del Pérmico. [13]

Un artículo de 2024 postuló que el arte rupestre de una criatura imaginaria superficialmente parecida a una morsa con colmillos curvados hacia abajo creado por el pueblo San de Sudáfrica antes de 1835 puede haberse inspirado en parte en cráneos fósiles de dicinodontes que se erosionan de las rocas de la zona. [14]
Los dicinodontes son conocidos por la ciencia desde mediados del siglo XIX. El geólogo sudafricano Andrew Geddes Bain dio la primera descripción de los dicinodontes en 1845. En ese momento, Bain era supervisor de la construcción de caminos militares bajo el Cuerpo de Ingenieros Reales y había encontrado muchos fósiles de reptiles durante sus estudios de Sudáfrica. Bain describió estos fósiles en una carta de 1845 publicada en Transactions of the Geological Society of London , llamándolos "bidentales" por sus dos colmillos prominentes. [15] Ese mismo año, el paleontólogo inglés Richard Owen nombró dos especies de dicinodontes de Sudáfrica: Dicynodon lacerticeps y Dicynodon bainii . Dado que Bain estaba preocupado por el Cuerpo de Ingenieros Reales, quería que Owen describiera sus fósiles de manera más extensa. Owen no publicó una descripción hasta 1876 en su Catálogo descriptivo e ilustrado de los reptiles fósiles de Sudáfrica en la colección del Museo Británico . [16] Para entonces, se habían descrito muchos más dicinodontos. En 1859, otra especie importante llamada Ptychognathus declivis fue nombrada de Sudáfrica. En el mismo año, Owen nombró al grupo Dicynodontia. [17] En su Catálogo descriptivo e ilustrado , Owen honró a Bain al erigir Bidentalia como un nombre de reemplazo para su Dicynodontia. [16] El nombre Bidentalia cayó rápidamente en desuso en los años siguientes, reemplazado por la popularidad de Dicynodontia de Owen. [18]

Los dicinodontos aparecieron por primera vez durante el Pérmico medio en el hemisferio sur, siendo Sudáfrica el centro de su diversidad conocida, y experimentaron una rápida radiación evolutiva , distribuyéndose globalmente y entre los vertebrados terrestres más exitosos y abundantes durante el Pérmico tardío . [19] [20] Durante este tiempo, incluyeron una gran variedad de ecotipos, incluidos herbívoros grandes, medianos y pequeños y excavadores similares a topos de extremidades cortas. [21]
Solo se sabe que cuatro linajes sobrevivieron a la Gran Mortandad ; los tres primeros representados con un solo género cada uno: Myosaurus , Kombuisia y Lystrosaurus , siendo este último el herbívoro más común y extendido del Induano ( Triásico temprano ). Ninguno de ellos sobrevivió mucho tiempo en el Triásico. El cuarto grupo fue el de los Kannemeyeriiformes , los únicos dicinodontos que se diversificaron durante el Triásico. [22] Estos animales robustos, del tamaño de un cerdo o un buey, fueron los herbívoros más abundantes en todo el mundo desde el Olenekiense hasta el Ladiniano . Para el Carniense habían sido suplantados por cinodontos traversodontes y reptiles rincosaurios . Durante el Noriano (mediados del Triásico Tardío), quizás debido a la creciente aridez, declinaron drásticamente, y el papel de los grandes herbívoros fue asumido por los dinosaurios sauropodomorfos . [ cita requerida ]
Los fósiles de un dicinodonte del tamaño de un elefante asiático, Lisowicia bojani, descubiertos en Polonia indican que los dicinodontes sobrevivieron al menos hasta finales del Noriense o principios del Rético (último Triásico); este animal también fue la especie de dicinodonte más grande conocida. [23] [24]
Seis fragmentos de hueso fósil descubiertos en Queensland , Australia, fueron interpretados como restos de un cráneo en 2003. Esto sugirió indicar que los dicinodontes sobrevivieron hasta el Cretácico en el sur de Gondwana . [25] La afinidad dicinodonte de estos especímenes fue cuestionada (incluida una propuesta de que pertenecían a un crocodiliforme baurusuquio por Agnolin et al. en 2010), [26] y en 2019 Knutsen y Oerlemans consideraron que este fósil era de edad Plioceno - Pleistoceno , y lo reinterpretaron como un fósil de un gran mamífero, probablemente un diprotodontido . [27]
Con el declive y extinción de los kannemeyeridos, no habría más herbívoros sinápsidos grandes dominantes hasta la época del Paleoceno medio (60 Ma), cuando los mamíferos , descendientes lejanos de los cinodontos , comenzaron a diversificarse después de la extinción de los dinosaurios no aviares.
Dicynodontia fue nombrada originalmente por el paleontólogo inglés Richard Owen . Fue erigida como una familia del orden Anomodontia e incluía los géneros Dicynodon y Ptychognathus . Otros grupos de Anomodontia incluían Gnathodontia, que incluía a Rhynchosaurus (ahora conocido por ser un archosauromorfo ) y Cryptodontia , que incluía a Oudenodon . Los cryptodontes se distinguían de los dicinodontos por su ausencia de colmillos. Aunque carece de colmillos, Oudenodon ahora se clasifica como un dicinodonte, y el nombre Cryptodontia ya no se usa. Thomas Henry Huxley revisó Dicynodontia de Owen como un orden que incluía a Dicynodon y Oudenodon . [28] Dicynodontia fue luego clasificada como un suborden o infraorden con el grupo más grande Anomodontia, que se clasifica como un orden. La clasificación de la dicinodoncia ha variado en estudios recientes: Ivakhnenko (2008) la considera un suborden, Ivanchnenko (2008) la considera un infraorden y Kurkin (2010) la considera un orden. [29]
Muchos taxones superiores, incluyendo infraórdenes y familias, han sido erigidos como un medio para clasificar el gran número de especies de dicinodontes. Cluver y King (1983) reconocieron varios grupos principales dentro de Dicynodontia, incluyendo Eodicynodontia (que contiene solo Eodicynodon ), Endothiodontia (que contiene solo Endothiodontidae ), Pristerodontia ( Pristerodontidae , Cryptodontidae , Aulacephalodontidae , Dicynodontidae , Lystrosauridae y Kannemeyeriidae ), Kingoriamorpha (que contiene solo Kingoriidae ), Diictodontia ( Diictodontidae , Robertiidae , Cistecephalidae , Emydopidae y Myosauridae ) y Venyukoviamorpha . [30] La mayoría de estos taxones ya no se consideran válidos. Kammerer y Angielczyk (2009) sugirieron que la problemática taxonomía y nomenclatura de Dicynodontia y otros grupos resulta de la gran cantidad de estudios conflictivos y la tendencia a establecer nombres inválidos por error. [18]






.jpg/1280px-Gordonia_traquairi_restoration_and_skull_(cropped).jpg)



A continuación se muestra un cladograma modificado de Angielczyk et al. (2021): [31]
Se conoce arte rupestre con serpiente cornuda en la cueva de La Belle France en Sudáfrica , que a menudo se confunde con el de Dingonek . Es posible que esté basado en fósiles de dicinodontes. [32]