
Los rapes son peces del orden de los teleósteos Lophiiformes ( / ˌlɒf iɪˈfɔːrm iːz / ). [ 1] Son peces óseos llamados así por su modo característico de depredación , en el que un rayo de aleta luminiscente modificado ( la esca o illicium ) actúa como un señuelo para otros peces . La luminiscencia proviene de bacterias simbióticas , que se cree que se adquieren del agua de mar, [2] [3] que habitan en el mar y sus alrededores.
Algunos rapes se caracterizan por un dimorfismo sexual extremo y una simbiosis sexual entre el macho pequeño y la hembra mucho más grande, como se observa en el suborden Ceratioidei , el rape de aguas profundas. En estas especies, los machos pueden ser varios órdenes de magnitud más pequeños que las hembras. [4]
Los rapes se encuentran en todo el mundo. Algunos son pelágicos (viven lejos del fondo marino), mientras que otros son bentónicos (viven cerca del fondo marino). Algunos viven en aguas profundas (como los ceratíidos ), mientras que otros viven en la plataforma continental , como los peces sapo y los lofíidos (rape o pez ganso). Las formas pelágicas suelen estar comprimidas lateralmente , mientras que las formas bentónicas suelen estar extremadamente comprimidas dorsoventralmente (deprimidas), a menudo con grandes bocas que apuntan hacia arriba. [ cita requerida ]
Los primeros fósiles de rape proceden de la formación Monte Bolca del Eoceno en Italia , y ya muestran una diversificación significativa en las familias modernas que componen el orden. [5] Dado esto, y su estrecha relación con los Tetraodontiformes , que se conocen a partir de fósiles del Cretácico , es probable que se originaran durante el Cretácico. [6] [7]
Un estudio filogenético del genoma mitocondrial de 2010 sugirió que los rapes se diversificaron en un corto período del Cretácico temprano a mediados, entre 130 y 100 millones de años atrás. [8] Una preimpresión más reciente reduce este tiempo al Cretácico tardío , entre 92 y 61 millones de años atrás. [7] Otros estudios indican que los rapes solo se originaron poco después del evento de extinción del Cretácico-Paleógeno como parte de una radiación adaptativa masiva de percomorfos , aunque esto choca con la extensa diversidad ya conocida del grupo en el Eoceno. [7] [9] Un estudio de 2024 encontró que todos los subórdenes de rape probablemente divergieron entre sí durante el Cretácico Superior y el Paleoceno , pero las múltiples familias de rape de aguas profundas ( Ceratioidei ), así como su parasitismo sexual característico , se originaron durante el Eoceno en una rápida radiación después del máximo térmico del Paleoceno-Eoceno . [10]
Los peces rape están clasificados según la quinta edición de Peces del mundo , como se establece a continuación, en cinco subórdenes y 18 familias. [11] [12] Los siguientes taxones se han organizado para mostrar sus relaciones evolutivas. [4] [13]
A continuación se muestran las relaciones de los subórdenes dentro de Lophiiformes tal como se establecen en Frogfishes of the world: systemics, zoogeography, and behavioral ecological ecology de Pietsch y Grobecker de 1987. [8]

_(16059517767).jpg/1280px-Hairy_frogfish_(Antennarius_striatus)_(16059517767).jpg)
Se ha descubierto en estudios filogenéticos que tanto los Lophiiformes como los Tetraodontiformes anidan dentro de los Acanthuriformes y, por lo tanto, se clasifican como clados dentro de ese taxón. [14]
Todos los peces rape son carnívoros y, por lo tanto, están adaptados para capturar presas. Las especies de aguas profundas, que varían en color desde el gris oscuro hasta el marrón oscuro, tienen cabezas grandes que albergan enormes bocas en forma de medialuna llenas de dientes largos en forma de colmillos, inclinados hacia adentro para atrapar presas de manera eficiente. Su longitud puede variar de 2 a 18 cm (1 a 7 pulgadas), y algunos tipos alcanzan los 100 cm (39 pulgadas), [15] pero esta variación se debe en gran medida al dimorfismo sexual, ya que las hembras son mucho más grandes que los machos. [16] Los peces sapo y otras especies de peces rape de aguas poco profundas son depredadores de emboscada y, a menudo, aparecen camuflados como rocas, esponjas o algas. [17]
Los peces rape tienen una aleta, o illicium, hacia el extremo distal de su cuerpo en su primera de dos aletas dorsales que se extiende hasta el hocico y actúa como un mecanismo de atracción donde la presa se acercará cara a cara. [18] El illicium se mueve hacia adelante y hacia atrás mediante cinco pares distintos de músculos: a saber, los músculos erectores y depresores más cortos que dictan el movimiento del hueso ilíaco, junto con los músculos inclinadores, transportadores y retractores que ayudan al movimiento del pterigióforo . [19]
En particular, si consideramos a Cryptopsaras couesii , este rape ceratioideo de aguas profundas tiene una biomecánica rotacional única en su musculatura. Los robustos músculos retractor y transportador se mueven en un patrón sinuoso en direcciones opuestas a lo largo del pterigióforo, que existe en una cresta longitudinal profunda a lo largo del cráneo. [19] Además, el inclinador largo y delgado del rape ceratioideo de aguas profundas permite un rango claramente amplio de movimiento anterior y posterior , lo que ayuda al movimiento del aparato de atracción para ayudar en la emboscada de la presa. [19]
La mayoría de las hembras adultas de rape ceratioideo tienen un órgano luminiscente llamado esca en la punta de un rayo dorsal modificado (el illicium o caña de pescar; derivado del latín ēsca , "cebo"). Se ha planteado la hipótesis de que el órgano sirve para atraer presas en entornos oscuros y profundos, pero también sirve para llamar la atención de los machos hacia las hembras para facilitar el apareamiento.
La fuente de luminiscencia son bacterias simbióticas que habitan en y alrededor de la esca, encerradas en un reflector en forma de copa que contiene cristales, probablemente compuestos de guanina . El rape hace uso de estas relaciones simbióticas con bacterias luminosas extracelulares. [20] Atípico de los simbiontes luminosos que viven fuera de las células del huésped, las bacterias que se encuentran en los señuelos del rape están experimentando un cambio evolutivo hacia genomas más pequeños y menos desarrollados (reducción genómica) asistida por expansiones de transposones . [20] Solo un puñado de especies de simbiontes luminiscentes pueden asociarse con los rapes de aguas profundas. [3] En algunas especies, las bacterias reclutadas en la esca son incapaces de producir luminiscencia independientemente del rape, lo que sugiere que han desarrollado una relación simbiótica y las bacterias son incapaces de sintetizar todos los químicos necesarios para la luminiscencia por sí mismas. Dependen del pez para compensar la diferencia. Si bien las hembras que se encuentran en la mayoría de las familias de rape tienen bioluminiscencia, existen excepciones, incluidas las familias Caulophrynidae y Neoceratiidae . [21]
Los simbiontes bacterianos no se encuentran en niveles consistentes a lo largo de las etapas de desarrollo del rape o en las diferentes profundidades del océano. [22] La secuenciación de organismos larvarios del suborden Ceratioidei muestra una ausencia de simbiontes bacterianos, mientras que la secuenciación del rape adulto mostró niveles más altos de simbiontes bacterianos bioluminiscentes . [22] Esto se correlaciona con la región mesopelágica que tiene los niveles más altos de relaciones simbióticas en las muestras de rape, ya que aquí es donde residen los rapes adultos durante la mayor parte de sus vidas después de su etapa larvaria. [22] La microscopía electrónica de estas bacterias en algunas especies revela que son bacilos Gram-negativos que carecen de cápsulas , esporas o flagelos . Tienen paredes celulares de doble capa y mesosomas . Un poro conecta la esca con el agua de mar, lo que permite la eliminación de bacterias muertas y desechos celulares, y permite que el pH y la tonicidad del medio de cultivo permanezcan constantes. Esto, así como la temperatura constante de la zona batipelágica habitada por estos peces, es crucial para la viabilidad a largo plazo de los cultivos bacterianos. [23]
La glándula de luz siempre está abierta al exterior, por lo que es posible que el pez adquiera las bacterias del agua de mar. Sin embargo, parece que cada especie utiliza sus propias especies particulares de bacterias, y estas bacterias nunca se han encontrado en el agua de mar. Haygood (1993) teorizó que las yescas descargan bacterias durante el desove y, de ese modo, las bacterias se transfieren a los huevos. [23]
Algunas evidencias muestran que algunos peces rape adquirieron sus simbiontes bioluminiscentes del entorno local. El material genético de las bacterias simbiontes se encuentra cerca del pez rape, lo que indica que es muy probable que el pez rape y sus bacterias asociadas no hayan evolucionado juntos y que las bacterias emprendan viajes difíciles para ingresar al huésped. [3] En un estudio sobre peces rape ceratioides en el Golfo de México, los investigadores notaron que los microbios bioluminiscentes asociados al huésped confirmados no están presentes en los especímenes larvarios ni durante el desarrollo del huésped. Los ceratioides probablemente adquirieron sus simbiontes bioluminiscentes del agua de mar. [2] Photobacterium phosphoreum y los miembros del clado kishitanii constituyen el simbionte bioluminiscente principal o único de varias familias de peces luminosos de aguas profundas. [24]
Se sabe que la composición genética de las bacterias simbiontes ha sufrido cambios desde que se asociaron con su anfitrión. [3] En comparación con sus parientes de vida libre, los genomas simbiontes del rape de aguas profundas se reducen en tamaño en un 50%. Se encuentran reducciones en las vías de síntesis de aminoácidos y en las capacidades para utilizar diversos azúcares. Sin embargo, los genes involucrados en la quimiotaxis y la motilidad que se cree que son útiles solo fuera del anfitrión se conservan en el genoma. El genoma simbionte contiene una gran cantidad de pseudogenes y muestra expansiones masivas de elementos transponibles . El proceso de reducción del genoma aún está en curso en estos linajes simbiontes, y la pérdida de genes puede conducir a la dependencia del anfitrión. [25]
En la mayoría de las especies, una boca ancha se extiende por toda la circunferencia anterior de la cabeza, y bandas de dientes inclinados hacia adentro recubren ambas mandíbulas. Los dientes pueden estar deprimidos de modo que no impidan que un objeto se deslice hacia el estómago, pero evitan que se escape de la boca. [26] El rape es capaz de distender tanto su mandíbula como su estómago, ya que sus huesos son delgados y flexibles, hasta alcanzar un tamaño enorme, lo que le permite tragar presas hasta dos veces más grandes que su cuerpo entero. [27]
En 2005, cerca de Monterey, California , a 1.474 metros de profundidad, un ROV filmó a una hembra de rape ceratioideo del género Oneirodes durante 24 minutos. Cuando se acercaba, el pez retrocedía rápidamente, pero en el 74% de las imágenes de vídeo, se desplazaba pasivamente, orientado en cualquier ángulo. Al avanzar, nadaba de forma intermitente a una velocidad de 0,24 longitudes corporales por segundo, batiendo sus aletas pectorales en sincronía. El comportamiento letárgico de este depredador de emboscada es adecuado para el entorno pobre en energía de las profundidades marinas. [28]
Otra observación in situ de tres rapes de nariz de látigo diferentes mostró un comportamiento inusual de natación invertida. Se observó que los peces flotaban invertidos completamente inmóviles con el illicium colgando rígidamente en un ligero arco frente al pez. El illicium colgaba sobre pequeñas madrigueras visibles. Se sugirió que esto es un esfuerzo por atraer presas y un ejemplo de búsqueda de alimento y depredación oportunista de baja energía. Cuando el ROV se acercó a los peces, exhibieron una natación explosiva, todavía invertida. [29]
La mandíbula y el estómago del rape pueden extenderse para permitirle consumir presas de hasta el doble de su tamaño. Debido a la cantidad limitada de alimento disponible en el entorno del rape, esta adaptación le permite almacenar alimentos cuando hay abundancia. [30]

El nombre de "pez rape" deriva del método característico de depredación de la especie. Los peces rape suelen tener al menos un filamento largo que brota de la mitad de sus cabezas, llamado illicium. El illicium son las tres primeras espinas separadas y modificadas de la aleta dorsal anterior . En la mayoría de las especies de peces rape, el filamento más largo es el primero. Esta primera espina sobresale por encima de los ojos del pez y termina en un crecimiento irregular de carne (la esca), y puede moverse en todas direcciones. Los peces rape pueden mover la esca para que parezca una presa, lo que atrae a la presa del pez rape lo suficientemente cerca para que el pez rape la devore entera. [31] Algunos peces rape de aguas profundas de la zona batipelágica también emiten luz desde su esca para atraer a sus presas. [32]
Debido a que los rapes son recolectores oportunistas, muestran una variedad de presas preferidas con peces en los extremos del espectro de tamaño, al mismo tiempo que muestran una mayor selectividad por ciertas presas. Un estudio que examinó el contenido estomacal de rapes de aleta de hilo en la costa del Pacífico de América Central encontró que estos peces comían principalmente dos categorías de presas bentónicas : crustáceos y peces teleósteos . La presa más frecuente fueron los camarones pandalidos . El 52% de los estómagos examinados estaban vacíos, lo que respalda las observaciones de que los rapes son consumidores de poca energía. [33]
.gif/1280px-Haplophryne_mollis_(female,_with_atrophied_male_attached).gif)

Algunos peces rape, como los ceratiidae, o diablos marinos, emplean un método de apareamiento inusual. [34] Debido a que los individuos son raros localmente, los encuentros también son muy raros. Por lo tanto, encontrar una pareja es problemático. Cuando los científicos comenzaron a capturar peces rape ceratioides, notaron que todos los especímenes eran hembras. Estos individuos tenían un tamaño de unos pocos centímetros y casi todos tenían lo que parecían ser parásitos adheridos a ellos. Resultó que estos "parásitos" eran ceratioides machos altamente dimórficos. Esto indica que algunos taxones de peces rape utilizan un sistema de apareamiento poliándrico . En algunas especies de peces rape, la fusión entre macho y hembra durante la reproducción es posible debido a la falta de claves del sistema inmunológico que permiten que los anticuerpos maduren y creen receptores para las células T. [35] Se supone que han desarrollado nuevas estrategias inmunológicas que compensan la pérdida de las funciones de los linfocitos B y T que se encuentran en un sistema inmunológico adaptativo . [36]
Algunos ceratioides dependen de la reproducción parabiótica . Los machos que viven en libertad y las hembras no parasitadas de estas especies nunca tienen gónadas completamente desarrolladas. Por lo tanto, los machos nunca maduran sin unirse a una hembra y mueren si no pueden encontrar una. [4] Al nacer, los ceratioides macho ya están equipados con órganos olfativos extremadamente bien desarrollados que detectan olores en el agua. Los machos de algunas especies también desarrollan ojos grandes y altamente especializados que pueden ayudar a identificar parejas en entornos oscuros. Los ceratioides machos son significativamente más pequeños que un rape hembra y pueden tener problemas para encontrar alimento en las profundidades marinas. Además, el crecimiento de los canales alimentarios de algunos machos se atrofia, lo que les impide alimentarse. Algunos taxones tienen mandíbulas que nunca son adecuadas o efectivas para capturar presas. [37] Estas características significan que el macho debe encontrar rápidamente un rape hembra para evitar la muerte. Los órganos olfativos sensibles ayudan al macho a detectar las feromonas que señalan la proximidad de un rape hembra.
Los métodos que utilizan los rapes para localizar a sus parejas varían. Algunas especies tienen ojos diminutos que no son aptos para identificar a las hembras, mientras que otras tienen fosas nasales poco desarrolladas, lo que hace que sea poco probable que encuentren hembras de manera efectiva mediante el olfato. [4] Cuando un macho encuentra a una hembra, le muerde la piel y libera una enzima que digiere la piel de su boca y el cuerpo de ella, fusionando a la pareja hasta el nivel de los vasos sanguíneos. [36] El macho se vuelve dependiente de la hembra huésped para sobrevivir al recibir nutrientes a través de su sistema circulatorio compartido y, a cambio, proporciona esperma a la hembra. Después de la fusión, los machos aumentan de volumen y se vuelven mucho más grandes en relación con los machos de vida libre de la especie. Viven y siguen siendo reproductivamente funcionales mientras viva la hembra, y pueden participar en múltiples desoves. [4] Este dimorfismo sexual extremo garantiza que cuando la hembra esté lista para desovar, tenga una pareja disponible de inmediato. [36] Se pueden incorporar varios machos en una sola hembra individual, con hasta ocho machos en algunas especies, aunque algunos taxones parecen tener una regla de "un macho por hembra". [4]
La simbiosis no es el único método de reproducción en el rape. De hecho, muchas familias, incluidas Melanocetidae , Himantolophidae , Diceratiidae y Gigantactinidae , no muestran evidencia de simbiosis masculina. [37] Las hembras de algunas de estas especies contienen ovarios grandes y desarrollados y los machos que viven en libertad tienen testículos grandes, lo que sugiere que estos individuos sexualmente maduros pueden desovar durante una unión sexual temporal que no implica fusión de tejidos. Los machos de estas especies también tienen mandíbulas bien dentadas que son mucho más efectivas para cazar que las que se observan en especies simbióticas. [37]
La simbiosis sexual puede ser una estrategia opcional en algunas especies de rape. [4] En Oneirodidae , se han reportado hembras portadoras de machos simbióticos en Leptacanthichthys y Bertella —y otros que aún no tenían gónadas completamente funcionales desarrolladas. [4] Una teoría sugiere que los machos se unen a las hembras independientemente de su propio desarrollo reproductivo si la hembra no es sexualmente madura, pero cuando tanto el macho como la hembra son maduros, desovan y luego se separan. [4]
Una explicación de la evolución de la simbiosis sexual es que la densidad relativamente baja de hembras en los ambientes de aguas profundas deja pocas oportunidades para la elección de pareja entre los peces rape. Las hembras siguen siendo grandes para permitir la fecundidad , como lo evidencian sus grandes ovarios y huevos. Se esperaría que los machos se encogieran para reducir los costos metabólicos en entornos pobres en recursos y desarrollarían habilidades altamente especializadas para encontrar hembras. Si un macho logra encontrar una hembra, entonces es más probable que el apego simbiótico mejore la aptitud de por vida en relación con la vida libre, particularmente cuando la perspectiva de encontrar futuras parejas es baja. Una ventaja adicional de la simbiosis es que el esperma del macho puede usarse en múltiples fertilizaciones, ya que siempre permanece disponible para la hembra para el apareamiento. Las densidades más altas de encuentros entre machos y hembras podrían correlacionarse con especies que demuestran simbiosis facultativa o simplemente usan un apareamiento de contacto temporal más tradicional. [38]
La puesta del rape del género Lophius consiste en una fina lámina de material gelatinoso transparente de 25 cm (10 in) de ancho y más de 10 m (33 ft) de largo. [39] Una lámina de huevos de este tipo es poco común entre los peces. Los huevos en esta lámina están en una sola capa, cada uno en su propia cavidad. La puesta se encuentra libre en el mar. Las larvas nadan libremente y tienen las aletas pélvicas alargadas en filamentos. [26]
Las especies de Lophius del noroeste de Europa son objeto de una intensa pesca y el CIEM las clasifica como "fuera de los límites biológicos seguros". [40] Además, se sabe que los rapes suben ocasionalmente a la superficie durante El Niño , dejando grandes grupos de rapes muertos flotando en la superficie. [40]
En 2010, Greenpeace Internacional añadió el rape americano ( Lophius americanus ), el rape ( Lophius piscatorius ) y el rape de vientre negro ( Lophius budegassa ) a su lista roja de mariscos, una lista de peces que se venden comúnmente en todo el mundo y que tienen una alta probabilidad de provenir de pesquerías no sostenibles. Peces de la Lista Roja [41] [42]
Una familia, los Lophiidae , es de interés comercial y se encuentra en la pesca en Europa occidental, el este de Norteamérica, África y el este de Asia. En Europa y Norteamérica, la carne de la cola de los peces del género Lophius , conocidos como rape o pez ganso (Norteamérica), se utiliza ampliamente en la cocina y a menudo se compara con la cola de langosta en cuanto a sabor y textura.
En África, los países de Namibia y la República de Sudáfrica registran las mayores capturas. [43] En Asia, especialmente en Japón, el hígado de rape, conocido como ankimo , se considera un manjar. [44] El rape se consume especialmente en Corea del Sur, donde aparece como ingrediente principal en platos como el Agujjim .
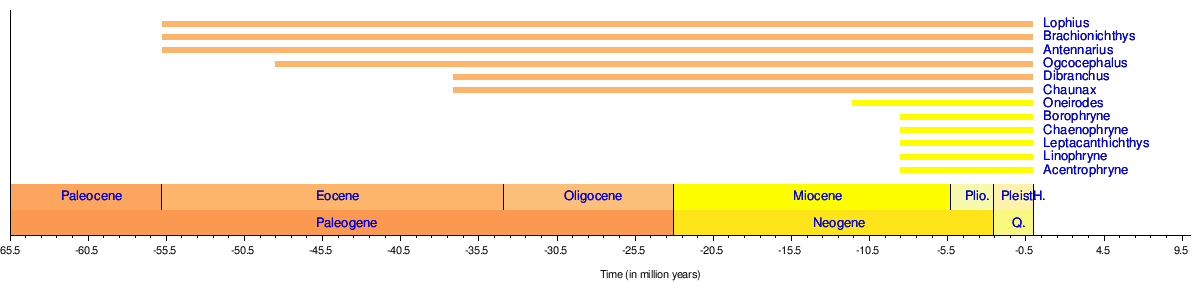
Los peces rape aparecen en el registro fósil de la siguiente manera: [45] [46] [47] [48]
Otros confían en la técnica adoptada por un lobo con piel de oveja: imitan a una especie inofensiva. ... Otros depredadores incluso imitan a la presa de sus presas: los rapes (Lophiiformes) y las tortugas caimanes (Macroclemys temmincki) pueden hacer crecer excrecencias carnosas de sus aletas o lenguas y atraer pequeños peces depredadores cerca de sus bocas.