La biología del desarrollo es el estudio del proceso mediante el cual los animales y las plantas crecen y se desarrollan. La biología del desarrollo también abarca la biología de la regeneración , la reproducción asexual , la metamorfosis y el crecimiento y la diferenciación de las células madre en el organismo adulto.
Los principales procesos implicados en el desarrollo embrionario de los animales son: la formación de patrones tisulares (a través de la especificación regional y la diferenciación celular pautada ); el crecimiento tisular ; y la morfogénesis tisular .
El desarrollo de las plantas implica procesos similares al de los animales. Sin embargo, las células vegetales son en su mayoría inmóviles, por lo que la morfogénesis se logra mediante un crecimiento diferencial, sin movimientos celulares. Además, las señales inductivas y los genes involucrados son diferentes a los que controlan el desarrollo animal.
La biología generativa es la ciencia generativa que explora la dinámica que guía el desarrollo y la evolución de una forma morfológica biológica. [1] [2] [3]

La diferenciación celular es el proceso por el cual surgen diferentes tipos de células funcionales en el desarrollo. Por ejemplo, las neuronas, las fibras musculares y los hepatocitos (células del hígado) son tipos bien conocidos de células diferenciadas. Las células diferenciadas suelen producir grandes cantidades de unas pocas proteínas que son necesarias para su función específica y esto les da la apariencia característica que permite reconocerlas bajo el microscopio óptico. Los genes que codifican estas proteínas son muy activos. Normalmente, su estructura de cromatina es muy abierta, lo que permite el acceso de las enzimas de transcripción, y los factores de transcripción específicos se unen a secuencias reguladoras en el ADN para activar la expresión génica. [4] [5] Por ejemplo, NeuroD es un factor de transcripción clave para la diferenciación neuronal, la miogenina para la diferenciación muscular y HNF4 para la diferenciación de los hepatocitos. La diferenciación celular suele ser la etapa final del desarrollo, precedida por varios estados de compromiso que no están visiblemente diferenciados. Un solo tejido, formado a partir de un solo tipo de célula progenitora o célula madre, a menudo consta de varios tipos de células diferenciadas. El control de su formación implica un proceso de inhibición lateral, [6] basado en las propiedades de la vía de señalización Notch . [7] Por ejemplo, en la placa neural del embrión este sistema opera para generar una población de células precursoras neuronales en las que NeuroD está altamente expresado.
La regeneración indica la capacidad de regenerar una parte faltante. [8] Esto es muy frecuente entre las plantas, que muestran un crecimiento continuo, y también entre los animales coloniales como los hidroides y las ascidias. Pero el mayor interés de los biólogos del desarrollo se ha mostrado en la regeneración de partes en animales que viven en libertad. En particular, cuatro modelos han sido objeto de mucha investigación. Dos de ellos tienen la capacidad de regenerar cuerpos enteros: la hidra , que puede regenerar cualquier parte del pólipo a partir de un pequeño fragmento, [9] y los gusanos planarios , que normalmente pueden regenerar tanto la cabeza como la cola. [10] Ambos ejemplos tienen una renovación celular continua alimentada por células madre y, al menos en las planarias, se ha demostrado que al menos algunas de las células madre son pluripotentes . [11] Los otros dos modelos muestran solo una regeneración distal de los apéndices. Estos son los apéndices de los insectos, normalmente las patas de los insectos hemimetábolos como el grillo, [12] y las extremidades de los anfibios urodelos . [13] Actualmente se dispone de una considerable cantidad de información sobre la regeneración de las extremidades de los anfibios y se sabe que cada tipo de célula se regenera a sí misma, excepto en el caso de los tejidos conectivos, donde hay una considerable interconversión entre el cartílago, la dermis y los tendones. En términos del patrón de estructuras, esto está controlado por una reactivación de señales activas en el embrión. Todavía se debate sobre la vieja cuestión de si la regeneración es una propiedad "prístina" o "adaptativa". [14] Si es así, con un mejor conocimiento, podríamos esperar poder mejorar la capacidad regenerativa en los seres humanos. Si es así, se supone que cada caso de regeneración surgió por selección natural en circunstancias particulares de la especie, por lo que no cabría esperar que existieran reglas generales.

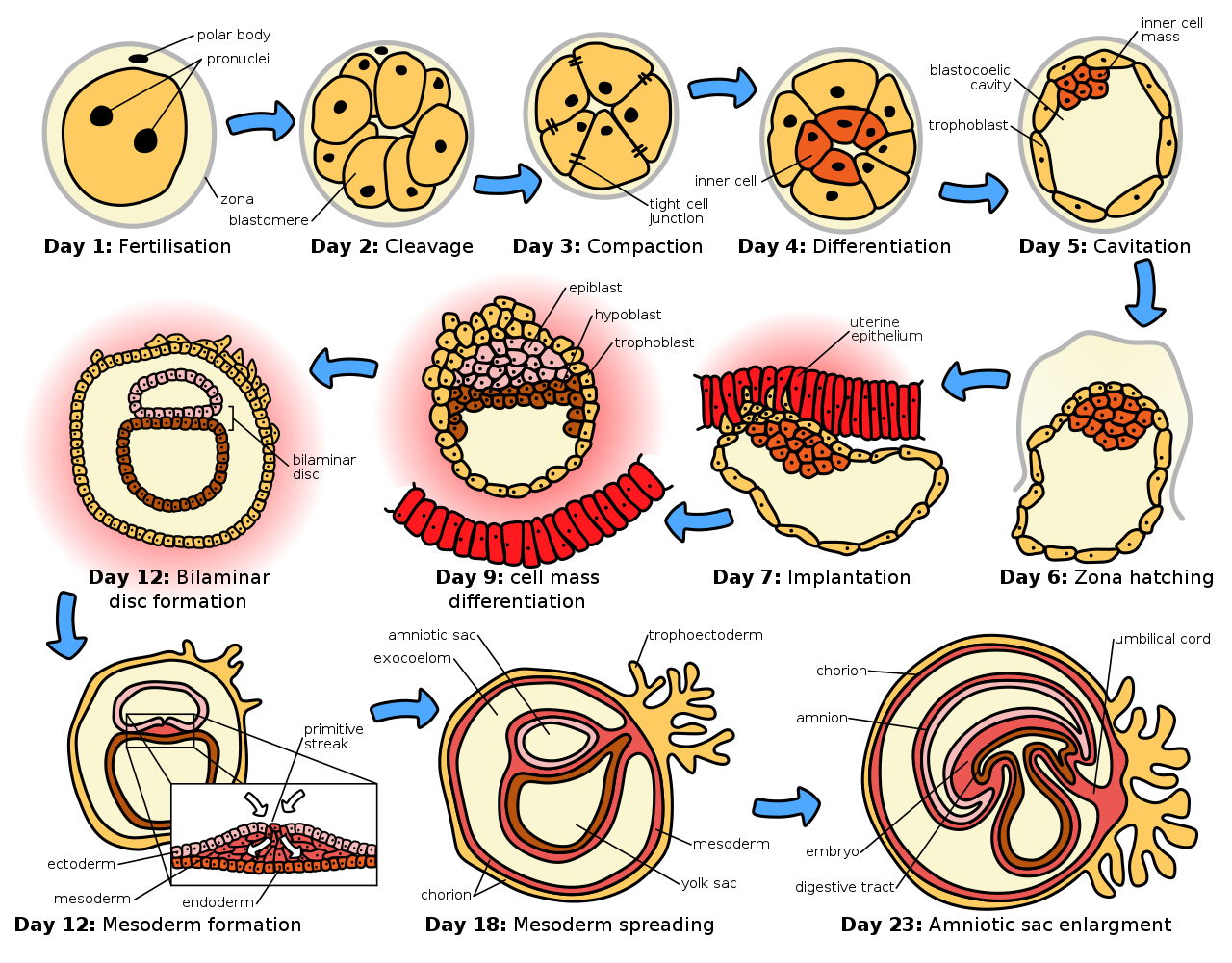
El espermatozoide y el óvulo se fusionan en el proceso de fertilización para formar un óvulo fertilizado, o cigoto . [15] Este atraviesa un período de divisiones para formar una bola o lámina de células similares llamada blástula o blastodermo . Estas divisiones celulares suelen ser rápidas y no hay crecimiento, por lo que las células hijas tienen la mitad del tamaño de la célula madre y el embrión completo se mantiene aproximadamente del mismo tamaño. Se denominan divisiones por segmentación .
Las células germinales primordiales del epiblasto del ratón (ver Figura: “Las etapas iniciales de la embriogénesis humana ”) experimentan una reprogramación epigenética extensa. [16] Este proceso implica la desmetilación del ADN en todo el genoma , la reorganización de la cromatina y el borrado de la impronta epigenética que conduce a la totipotencia . [16] La desmetilación del ADN se lleva a cabo mediante un proceso que utiliza la vía de reparación por escisión de bases del ADN. [17]
Los movimientos morfogenéticos convierten la masa celular en una estructura de tres capas que consiste en láminas multicelulares llamadas ectodermo , mesodermo y endodermo . Estas láminas se conocen como capas germinativas . Este es el proceso de gastrulación . Durante la segmentación y la gastrulación ocurren los primeros eventos de especificación regional. Además de la formación de las tres capas germinativas en sí, estas a menudo generan estructuras extraembrionarias, como la placenta de los mamíferos , necesaria para el soporte y la nutrición del embrión, [18] y también establecen diferencias de compromiso a lo largo del eje anteroposterior (cabeza, tronco y cola). [19]
La especificación regional se inicia con la presencia de determinantes citoplasmáticos en una parte del cigoto. Las células que contienen el determinante se convierten en un centro de señalización y emiten un factor inductor. Debido a que el factor inductor se produce en un lugar, se difunde y se desintegra, forma un gradiente de concentración, alto cerca de las células fuente y bajo más lejos. [20] [21] Las células restantes del embrión, que no contienen el determinante, son competentes para responder a diferentes concentraciones mediante la regulación positiva de genes de control del desarrollo específicos. Esto da como resultado que se establezca una serie de zonas, dispuestas a una distancia progresivamente mayor del centro de señalización. En cada zona se regula positivamente una combinación diferente de genes de control del desarrollo. [22] Estos genes codifican factores de transcripción que regulan positivamente nuevas combinaciones de actividad genética en cada región. Entre otras funciones, estos factores de transcripción controlan la expresión de genes que confieren propiedades adhesivas y de motilidad específicas a las células en las que están activos. Debido a estas diferentes propiedades morfogenéticas, las células de cada capa germinal se mueven para formar láminas de manera que el ectodermo termina en el exterior, el mesodermo en el medio y el endodermo en el interior. [23] [24]

Los movimientos morfogenéticos no solo cambian la forma y la estructura del embrión, sino que al llevar las láminas celulares a nuevas relaciones espaciales también hacen posibles nuevas fases de señalización y respuesta entre ellas. Además, los primeros movimientos morfogenéticos de la embriogénesis, como la gastrulación, la epibolia y la torsión , activan directamente las vías involucradas en la especificación del endomesodermo a través de procesos de mecanotransducción. [25] [26] Se sugirió que esta propiedad era heredada evolutivamente de la especificación del endomesodermo estimulada mecánicamente por el flujo hidrodinámico ambiental marino en los primeros organismos animales (primeros metazoos). [27] La torsión a lo largo del eje del cuerpo por una quiralidad zurda se encuentra en todos los cordados (incluidos los vertebrados) y se aborda mediante la teoría de la torsión axial . [28]
El crecimiento de los embriones es en su mayor parte autónomo. [29] En cada territorio de células, la tasa de crecimiento está controlada por la combinación de genes activos. Los embriones que viven en libertad no crecen en masa porque no tienen un suministro externo de alimentos. Pero los embriones alimentados por una placenta o un suministro de yema extraembrionaria pueden crecer muy rápido, y los cambios en la tasa de crecimiento relativa entre las partes de estos organismos ayudan a producir la anatomía general final.
Todo el proceso debe coordinarse en el tiempo y no se entiende cómo se controla. Puede haber un reloj maestro capaz de comunicarse con todas las partes del embrión que controle el curso de los acontecimientos, o el tiempo puede depender simplemente de secuencias causales locales de acontecimientos. [30]
Los procesos de desarrollo son muy evidentes durante el proceso de metamorfosis . Esto ocurre en varios tipos de animales. Ejemplos bien conocidos se ven en las ranas, que generalmente nacen como renacuajos y se metamorfosean en una rana adulta, y ciertos insectos que nacen como larvas y luego se remodelan a la forma adulta durante una etapa de pupa.
Todos los procesos de desarrollo enumerados anteriormente ocurren durante la metamorfosis. Algunos ejemplos que han sido especialmente estudiados incluyen la pérdida de la cola y otros cambios en el renacuajo de la rana Xenopus [31] [ 32] y la biología de los discos imaginales, que generan las partes del cuerpo adulto de la mosca Drosophila melanogaster [ 33] [34]
El desarrollo de las plantas es el proceso mediante el cual se originan y maduran las estructuras a medida que la planta crece. Se estudia en la anatomía y fisiología de las plantas , así como en la morfología de las plantas.
Las plantas producen constantemente nuevos tejidos y estructuras a lo largo de su vida a partir de meristemos [35] ubicados en las puntas de los órganos o entre los tejidos maduros. Por lo tanto, una planta viva siempre tiene tejidos embrionarios. Por el contrario, un embrión animal producirá muy pronto todas las partes del cuerpo que tendrá en su vida. Cuando el animal nace (o sale del huevo), tiene todas las partes de su cuerpo y a partir de ese momento solo crecerá más grande y madurará.
Las propiedades de organización que se observan en una planta son propiedades emergentes que son más que la suma de las partes individuales. “El ensamblaje de estos tejidos y funciones en un organismo multicelular integrado produce no sólo las características de las partes y procesos separados, sino también un conjunto completamente nuevo de características que no habrían sido predecibles sobre la base del examen de las partes separadas”. [36]
Una planta vascular comienza a partir de un cigoto unicelular , formado por la fecundación de un óvulo por un espermatozoide. A partir de ese momento, comienza a dividirse para formar un embrión vegetal a través del proceso de embriogénesis . A medida que esto sucede, las células resultantes se organizarán de manera que un extremo se convierta en la primera raíz, mientras que el otro extremo forma la punta del brote. En las plantas con semillas , el embrión desarrollará una o más "hojas de semilla" ( cotiledones ). Al final de la embriogénesis, la planta joven tendrá todas las partes necesarias para comenzar su vida.
Una vez que el embrión germina a partir de su semilla o planta madre, comienza a producir órganos adicionales (hojas, tallos y raíces) a través del proceso de organogénesis . Las nuevas raíces crecen a partir de los meristemos de la raíz ubicados en la punta de la raíz, y los nuevos tallos y hojas crecen a partir de los meristemos del brote ubicados en la punta del brote. [37] La ramificación ocurre cuando pequeños grupos de células que dejó el meristemo, y que aún no han experimentado una diferenciación celular para formar un tejido especializado, comienzan a crecer como la punta de una nueva raíz o brote. El crecimiento de cualquier meristemo de este tipo en la punta de una raíz o brote se denomina crecimiento primario y da como resultado el alargamiento de esa raíz o brote. El crecimiento secundario da como resultado el ensanchamiento de una raíz o brote a partir de divisiones de células en un cambium . [38]
Además del crecimiento por división celular , una planta puede crecer a través de la elongación celular . [39] Esto ocurre cuando las células individuales o grupos de células crecen más. No todas las células de la planta crecerán hasta la misma longitud. Cuando las células de un lado de un tallo crecen más y más rápido que las células del otro lado, el tallo se doblará hacia el lado de las células de crecimiento más lento como resultado. Este crecimiento direccional puede ocurrir a través de la respuesta de una planta a un estímulo particular, como la luz ( fototropismo ), la gravedad ( gravitropismo ), el agua ( hidrotropismo ) y el contacto físico ( tigmotropismo ).
El crecimiento y desarrollo de las plantas están mediados por hormonas vegetales específicas y reguladores del crecimiento vegetal (PGR) (Ross et al. 1983). [40] Los niveles de hormonas endógenas están influenciados por la edad de la planta, la resistencia al frío, la latencia y otras condiciones metabólicas; el fotoperíodo, la sequía, la temperatura y otras condiciones ambientales externas; y fuentes exógenas de PGR, por ejemplo, aplicadas externamente y de origen rizosférico.
Las plantas presentan variaciones naturales en su forma y estructura. Si bien todos los organismos varían de un individuo a otro, las plantas presentan un tipo adicional de variación. Dentro de un mismo individuo, se repiten partes que pueden diferir en forma y estructura de otras partes similares. Esta variación se observa con mayor facilidad en las hojas de una planta, aunque otros órganos como los tallos y las flores pueden mostrar variaciones similares. Existen tres causas principales de esta variación: efectos posicionales, efectos ambientales y juventud.
Los factores de transcripción y las redes reguladoras de la transcripción desempeñan un papel fundamental en la morfogénesis de las plantas y su evolución. Durante el aterrizaje de las plantas, surgieron muchas familias de factores de transcripción nuevos que se conectan preferentemente a las redes de desarrollo multicelular, reproducción y desarrollo de órganos, lo que contribuye a una morfogénesis más compleja de las plantas terrestres. [41]
La mayoría de las plantas terrestres comparten un ancestro común, las algas multicelulares. Un ejemplo de la evolución de la morfología vegetal se observa en los carófitos. Los estudios han demostrado que los carófitos tienen rasgos homólogos a los de las plantas terrestres. Existen dos teorías principales sobre la evolución de la morfología vegetal: la teoría homóloga y la teoría antitética. La teoría comúnmente aceptada para la evolución de la morfología vegetal es la teoría antitética. La teoría antitética afirma que las múltiples divisiones mitóticas que tienen lugar antes de la meiosis provocan el desarrollo del esporofito, que luego se desarrollará como un organismo independiente. [42]
Gran parte de la investigación en biología del desarrollo en las últimas décadas se ha centrado en el uso de un pequeño número de organismos modelo . Se ha comprobado que existe una gran conservación de los mecanismos de desarrollo en todo el reino animal. En el desarrollo temprano, las diferentes especies de vertebrados utilizan esencialmente las mismas señales inductivas y los mismos genes que codifican la identidad regional. Incluso los invertebrados utilizan un repertorio similar de señales y genes, aunque las partes del cuerpo formadas son significativamente diferentes. Cada organismo modelo tiene algunas ventajas experimentales particulares que les han permitido volverse populares entre los investigadores. En un sentido, son "modelos" para todo el reino animal y, en otro sentido, son "modelos" para el desarrollo humano, que es difícil de estudiar directamente por razones tanto éticas como prácticas. Los organismos modelo han sido muy útiles para dilucidar la naturaleza amplia de los mecanismos de desarrollo. Cuanto más detalles se buscan, más difieren entre sí y de los humanos.
También han sido populares para algunos propósitos los erizos de mar [51] [43] y las ascidias . [52] Para estudios de regeneración se utilizan anfibios urodelos como el ajolote Ambystoma mexicanum , [53] y también gusanos planarios como Schmidtea mediterranea . [10] También se ha demostrado que los organoides son un modelo eficiente para el desarrollo. [54] El desarrollo de plantas se ha centrado en el berro de thale Arabidopsis thaliana como organismo modelo. [55]
{{cite book}}: CS1 maint: location missing publisher (link){{cite book}}: |journal=ignorado ( ayuda )