
El espermatozoide ( pl.: espermatozoide o espermatozoides ) es la célula reproductora masculina , o gameto , en las formas anisógamas de reproducción sexual (formas en las que hay una célula reproductora femenina más grande y una masculina más pequeña). Los animales producen espermatozoides móviles con una cola conocida como flagelo , que se conocen como espermatozoides , mientras que algunas algas rojas y hongos producen células espermáticas no móviles, conocidas como espermatozoides . [1] Las plantas con flores contienen espermatozoides no móviles dentro del polen , mientras que algunas plantas más basales como los helechos y algunas gimnospermas tienen espermatozoides móviles. [2]
Los espermatozoides se forman durante el proceso conocido como espermatogénesis , que en los amniotas ( reptiles y mamíferos ) tiene lugar en los túbulos seminíferos de los testículos . [3] Este proceso implica la producción de varios precursores sucesivos de células espermáticas, comenzando por las espermatogonias , que se diferencian en espermatocitos . Luego, los espermatocitos experimentan meiosis , reduciendo su número de cromosomas a la mitad, lo que produce espermátidas . Luego, las espermátidas maduran y, en los animales, construyen una cola, o flagelo, que da lugar al espermatozoide maduro y móvil. Todo este proceso ocurre constantemente y demora alrededor de 3 meses desde su inicio hasta su finalización.
Los espermatozoides no pueden dividirse y tienen una vida útil limitada, pero después de fusionarse con los óvulos durante la fecundación , comienza a desarrollarse un nuevo organismo, comenzando como un cigoto totipotente . El espermatozoide humano es haploide , de modo que sus 23 cromosomas pueden unirse a los 23 cromosomas del óvulo femenino para formar una célula diploide con 46 cromosomas pareados. En los mamíferos , el esperma se almacena en el epidídimo y se libera a través del pene en el semen durante la eyaculación .
La palabra esperma se deriva de la palabra griega σπέρμα, esperma , que significa "semilla".
Se acepta generalmente que la isogamia es el ancestro de los espermatozoides y los óvulos. Dado que no existen registros fósiles de la evolución de los espermatozoides y los óvulos a partir de la isogamia, se hace mucho hincapié en los modelos matemáticos para comprender la evolución de los espermatozoides. [4]
Una hipótesis muy extendida afirma que los espermatozoides evolucionaron rápidamente, pero no hay evidencia directa de que los espermatozoides evolucionaran a un ritmo rápido o antes que otras características masculinas. [5]
La función principal del espermatozoide es llegar al óvulo y fusionarse con él para dar lugar a dos estructuras subcelulares: (i) el pronúcleo masculino que contiene el material genético y (ii) los centriolos que son estructuras que ayudan a organizar el citoesqueleto de microtúbulos . [ aclaración necesaria ]
El ADN nuclear de los espermatozoides es haploide , es decir, aportan solo una copia de cada par de cromosomas paternos . Las mitocondrias de los espermatozoides humanos no contienen ADN o contienen muy poco porque el ADNmt se degrada mientras los espermatozoides maduran, por lo que normalmente no aportan material genético a su descendencia. [6]


El espermatozoide de los mamíferos se puede dividir en dos partes conectadas por un cuello:
Durante la fertilización , el espermatozoide proporciona tres partes esenciales al ovocito : (1) un factor de señalización o activación, que hace que el ovocito metabólicamente inactivo se active; (2) el genoma paterno haploide ; (3) el centriolo, que es responsable de formar el centrosoma y el sistema de microtúbulos . [13]
Los espermatozoides de los animales se producen a través de la espermatogénesis dentro de las gónadas masculinas ( testículos ) a través de la división meiótica . El proceso inicial del espermatozoide tarda unos 70 días en completarse. El proceso comienza con la producción de espermatogonias a partir de precursores de células germinales . Estos se dividen y se diferencian en espermatocitos , que experimentan meiosis para formar espermátidas . En la etapa de espermátida, el espermatozoide desarrolla la conocida cola. La siguiente etapa en la que alcanza la madurez total tarda unos 60 días y se denomina espermatozoide . [14] Los espermatozoides salen del cuerpo masculino en un líquido conocido como semen . Los espermatozoides humanos pueden sobrevivir dentro del tracto reproductor femenino durante más de 5 días después del coito. [15] El semen se produce en las vesículas seminales , la glándula prostática y las glándulas uretrales .
En 2016, científicos de la Universidad Médica de Nanjing afirmaron haber producido células parecidas a las espermátidas de ratón a partir de células madre embrionarias de ratón de forma artificial. Inyectaron estas espermátidas en óvulos de ratón y produjeron crías. [16]
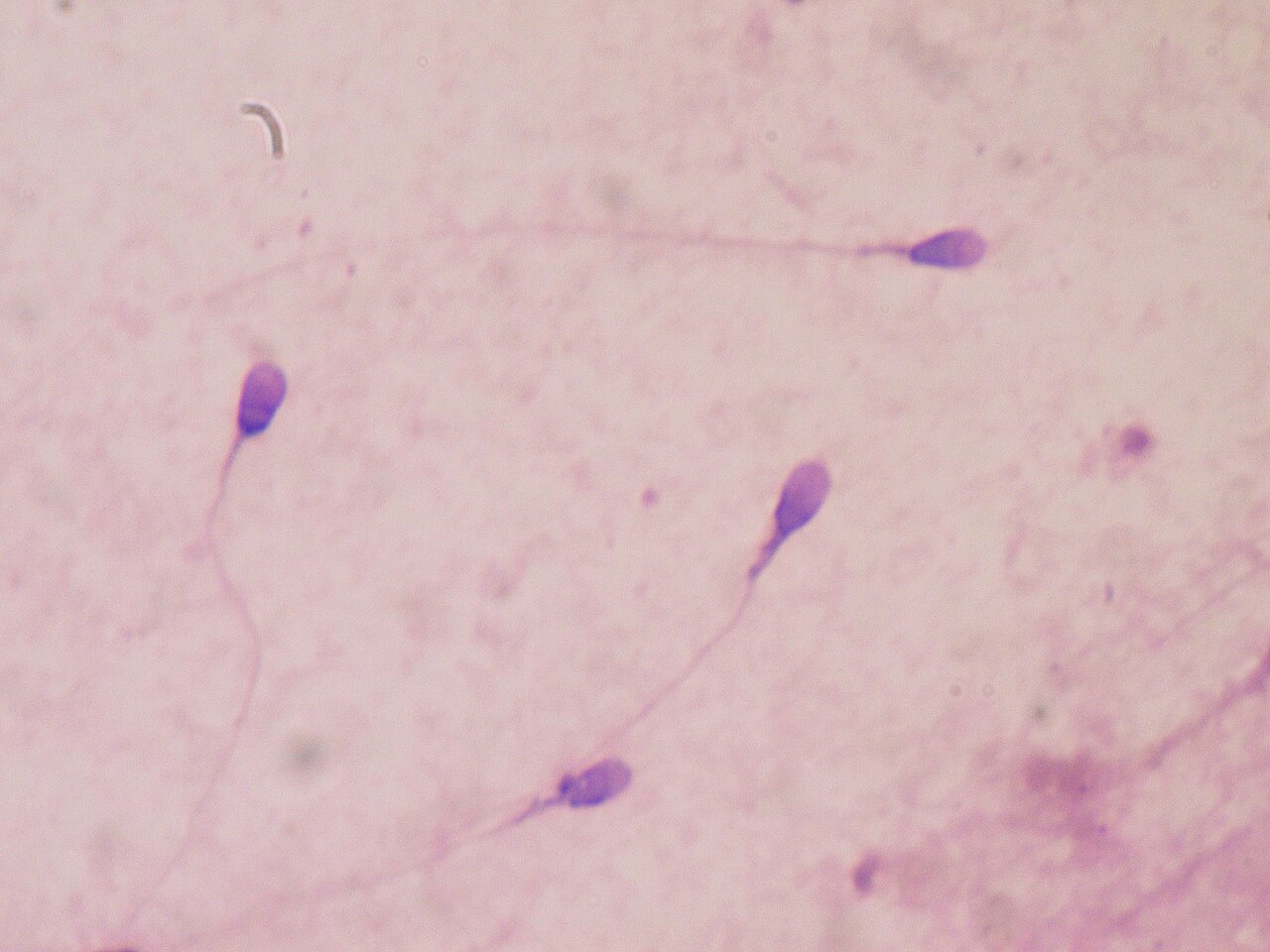
La cantidad y calidad de los espermatozoides son los principales parámetros de la calidad del semen, que es una medida de la capacidad del semen para lograr la fertilización . Por lo tanto, en los humanos, es una medida de la fertilidad en un hombre . La calidad genética de los espermatozoides, así como su volumen y motilidad, generalmente disminuyen con la edad . [17] Las roturas de doble cadena de ADN en los espermatozoides aumentan con la edad. [18] También la apoptosis disminuye con la edad, lo que sugiere que el aumento del ADN dañado de los espermatozoides a medida que los hombres envejecen se produce en parte como resultado de una selección celular menos eficiente (apoptosis) que opera durante o después de la espermatogénesis . [18]
Los daños en el ADN presentes en los espermatozoides en el período posterior a la meiosis pero anterior a la fecundación pueden repararse en el óvulo fecundado, pero si no se reparan, pueden tener efectos perjudiciales graves sobre la fertilidad y el embrión en desarrollo. Los espermatozoides humanos son particularmente vulnerables al ataque de radicales libres y a la generación de daño oxidativo en el ADN, [19] como el causado por la 8-Oxo-2'-desoxiguanosina .
La fase postmeiótica de la espermatogénesis del ratón es muy sensible a los agentes genotóxicos ambientales, porque a medida que las células germinales masculinas forman espermatozoides maduros pierden progresivamente la capacidad de reparar los daños en el ADN. [20] La irradiación de ratones machos durante la espermatogénesis tardía puede inducir daños que persisten durante al menos 7 días en los espermatozoides fertilizadores, y la interrupción de las vías de reparación de roturas de doble cadena del ADN materno aumenta las aberraciones cromosómicas derivadas de los espermatozoides. [21] El tratamiento de ratones machos con melfalán , un agente alquilante bifuncional empleado con frecuencia en quimioterapia, induce lesiones en el ADN durante la meiosis que pueden persistir en un estado no reparado a medida que las células germinales progresan a través de fases competentes para la reparación del ADN del desarrollo espermatogénico. [22] Estos daños no reparados en el ADN de los espermatozoides, después de la fertilización, pueden dar lugar a crías con diversas anomalías.
El tamaño de los espermatozoides está relacionado con la calidad de los mismos, al menos en algunos animales. Por ejemplo, los espermatozoides de algunas especies de moscas de la fruta ( Drosophila ) miden hasta 5,8 cm de largo, unas 20 veces más que la longitud de la propia mosca. Los espermatozoides más largos son mejores que sus contrapartes más cortas para desplazar a los competidores del receptáculo seminal de la hembra. El beneficio para las hembras es que sólo los machos sanos tienen genes "buenos" que pueden producir espermatozoides largos en cantidades suficientes para superar a sus competidores. [23] [24]
Algunos bancos de esperma pueden almacenar hasta 170 litros (37 galones imperiales; 45 galones estadounidenses) de esperma. [25]
Además de la eyaculación , es posible extraer espermatozoides mediante la extracción de espermatozoides testiculares .
En el mercado mundial, Dinamarca tiene un sistema bien desarrollado de exportación de esperma humano. Este éxito se debe principalmente a la reputación de los donantes de esperma daneses por ser de alta calidad [26] y, en contraste con la ley en los otros países nórdicos, les da a los donantes la opción de ser anónimos o no anónimos para la pareja receptora. [26] Además, los donantes de esperma nórdicos tienden a ser altos y altamente educados [27] y tienen motivos altruistas para sus donaciones, [27] en parte debido a la compensación monetaria relativamente baja en los países nórdicos. Más de 50 países en todo el mundo son importadores de esperma danés, incluidos Paraguay , Canadá , Kenia y Hong Kong . [26] Sin embargo, la Administración de Alimentos y Medicamentos (FDA) de los EE. UU. Ha prohibido la importación de cualquier esperma, motivada por un riesgo de transmisión de la enfermedad de Creutzfeldt-Jakob , aunque dicho riesgo es insignificante, ya que la inseminación artificial es muy diferente de la vía de transmisión de la enfermedad de Creutzfeldt-Jakob . [28] La prevalencia de la enfermedad de Creutzfeldt-Jakob en donantes es como máximo de uno en un millón, y si el donante fuera portador, las proteínas infecciosas todavía tendrían que cruzar la barrera hemato-testicular para hacer posible la transmisión. [28]
Los espermatozoides fueron observados por primera vez en 1677 por Antonie van Leeuwenhoek [29] usando un microscopio . Los describió como animálculos (pequeños animales), probablemente debido a su creencia en el preformacionismo , que pensaba que cada espermatozoide contenía un ser humano completamente formado pero pequeño. [ cita requerida ]
Los fluidos eyaculados se detectan mediante luz ultravioleta , independientemente de la estructura o el color de la superficie. [30] Las cabezas de esperma, por ejemplo, de hisopos vaginales, todavía se detectan mediante microscopía utilizando el método de "tinción de árbol de Navidad", es decir, tinción de Kernechtrot-Picroindigocarmín (KPIC). [31] [32]
Los espermatozoides de los gametofitos de las algas y de muchas plantas se producen en los gametangios masculinos ( anteridios ) mediante división mitótica . En las plantas con flores , los núcleos de los espermatozoides se producen dentro del polen . [33]

Los espermatozoides móviles se desplazan normalmente mediante flagelos y necesitan un medio acuoso para nadar hacia el óvulo para la fecundación. En los animales, la mayor parte de la energía necesaria para la movilidad de los espermatozoides se deriva del metabolismo de la fructosa transportada en el líquido seminal. Esto tiene lugar en las mitocondrias situadas en la parte media del espermatozoide (en la base de la cabeza del espermatozoide). Estas células no pueden nadar hacia atrás debido a la naturaleza de su propulsión. Los espermatozoides uniflagelados (con un flagelo) de los animales se denominan espermatozoides y se sabe que varían de tamaño. [ cita requerida ]
Los espermatozoides móviles también son producidos por muchos protistos y los gametofitos de briofitas , helechos y algunas gimnospermas como las cícadas y el ginkgo . Los espermatozoides son las únicas células flageladas en el ciclo de vida de estas plantas. En muchos helechos y licofitas , cícadas y ginkgo son multiflagelados (llevan más de un flagelo). [34]
En los nematodos , los espermatozoides son ameboides y se arrastran, en lugar de nadar, hacia el óvulo. [35]
Los espermatozoides inmóviles, llamados espermatozoides, carecen de flagelos y, por lo tanto, no pueden nadar. Los espermatozoides se producen en un espermatangio . [34]
Como los espermatozoides no pueden nadar, dependen de su entorno para llevarlos hasta el óvulo. Algunas algas rojas , como Polysiphonia , producen espermatozoides inmóviles que se esparcen por las corrientes de agua después de su liberación. [34] Los espermatozoides de los hongos de la roya están cubiertos de una sustancia pegajosa. Se producen en estructuras con forma de matraz que contienen néctar , que atrae a las moscas que transfieren los espermatozoides a las hifas cercanas para la fertilización en un mecanismo similar a la polinización por insectos en las plantas con flores . [36]
Las espermatias fúngicas (también llamadas picniosporas, especialmente en los Uredinales) pueden confundirse con los conidios . Los conidios son esporas que germinan independientemente de la fecundación, mientras que las espermatias son gametos necesarios para la fecundación. En algunos hongos, como Neurospora crassa , las espermatias son idénticas a los microconidios, ya que pueden realizar ambas funciones de fecundación y dar lugar a nuevos organismos sin fecundación. [37]
En casi todos los embriofitos , incluyendo la mayoría de las gimnospermas y todas las angiospermas , los gametofitos masculinos ( granos de polen ) son el modo principal de dispersión , por ejemplo a través del viento o la polinización de los insectos , eliminando la necesidad de agua para unir la brecha entre el macho y la hembra. Cada grano de polen contiene una célula espermatógena (generativa). Una vez que el polen aterriza en el estigma de una flor receptiva, germina y comienza a crecer un tubo polínico a través del carpelo . Antes de que el tubo llegue al óvulo , el núcleo de la célula generativa en el grano de polen se divide y da lugar a dos núcleos de esperma, que luego se descargan a través del tubo en el óvulo para la fertilización. [34]
En algunos protistos , la fertilización también implica que los núcleos de los espermatozoides, en lugar de las células, migren hacia el óvulo a través de un tubo de fertilización. Los oomicetos forman núcleos de espermatozoides en un anteridio sincítico que rodea a los óvulos. Los núcleos de los espermatozoides llegan a los óvulos a través de tubos de fertilización, similares al mecanismo del tubo polínico en las plantas. [34]
La mayoría de los espermatozoides tienen centriolos en el cuello del espermatozoide. [38] Los espermatozoides de muchos animales tienen dos centriolos típicos, conocidos como centriolo proximal y centriolo distal. Algunos animales (incluidos los humanos y los bovinos) tienen un solo centriolo típico, el centriolo proximal, así como un segundo centriolo con una estructura atípica. [11] Los ratones y las ratas no tienen centriolos espermáticos reconocibles. La mosca de la fruta Drosophila melanogaster tiene un solo centriolo y un centriolo atípico llamado centriolo proximal. [39]
La cola del espermatozoide es un tipo especializado de cilio (también conocido como flagelo). En muchos animales, la cola del espermatozoide se forma mediante el proceso único de ciliogénesis citosólica , en el que todo o parte del axonema de la cola del espermatozoide se forma en el citoplasma o queda expuesto al citoplasma. [40]
{{cite journal}}: CS1 maint: varios nombres: lista de autores ( enlace )