Las ecuaciones de Lotka-Volterra , también conocidas como modelo depredador-presa de Lotka-Volterra , son un par de ecuaciones diferenciales no lineales de primer orden , frecuentemente utilizadas para describir la dinámica de sistemas biológicos en los que interactúan dos especies, una como depredador y la otra como depredador . otro como presa. Las poblaciones cambian a través del tiempo según el par de ecuaciones:
dónde
La solución de las ecuaciones diferenciales es determinista y continua . Esto, a su vez, implica que las generaciones tanto del depredador como de la presa se superponen continuamente. [1]
El sistema de ecuaciones de Lotka-Volterra es un ejemplo de un modelo de Kolmogorov , [2] [3] [4] que es un marco más general que puede modelar la dinámica de los sistemas ecológicos con interacciones depredador-presa, competencia , enfermedades y mutualismo. .
Se supone que las presas tienen un suministro ilimitado de alimentos y se reproducen exponencialmente, a menos que estén sujetas a depredación; este crecimiento exponencial está representado en la ecuación anterior por el término αx . Se supone que la tasa de depredación de la presa es proporcional a la tasa a la que se encuentran los depredadores y la presa; esto está representado arriba por βxy . Si x o y son cero, entonces no puede haber depredación. Con estos dos términos, la ecuación de presa anterior se puede interpretar de la siguiente manera: la tasa de cambio de la población de la presa está dada por su propia tasa de crecimiento menos la tasa a la que es depredada.
El término δxy representa el crecimiento de la población de depredadores. (Obsérvese la similitud con la tasa de depredación; sin embargo, se utiliza una constante diferente, ya que la tasa a la que crece la población de depredadores no es necesariamente igual a la tasa a la que consume la presa). El término γy representa la tasa de pérdida de los depredadores debido a muerte natural o emigración; conduce a una decadencia exponencial en ausencia de presas. Por tanto, la ecuación expresa que la tasa de cambio de la población del depredador depende de la tasa a la que consume a sus presas, menos su tasa de mortalidad intrínseca.
El modelo depredador-presa de Lotka-Volterra hace una serie de suposiciones sobre el medio ambiente y la biología de las poblaciones de depredadores y presas: [5]
Es probable que ninguno de los supuestos anteriores se cumpla para las poblaciones naturales. Sin embargo, el modelo Lotka-Volterra muestra dos propiedades importantes de las poblaciones de depredadores y presas y estas propiedades a menudo se extienden a variantes del modelo en las que estos supuestos son relajados:
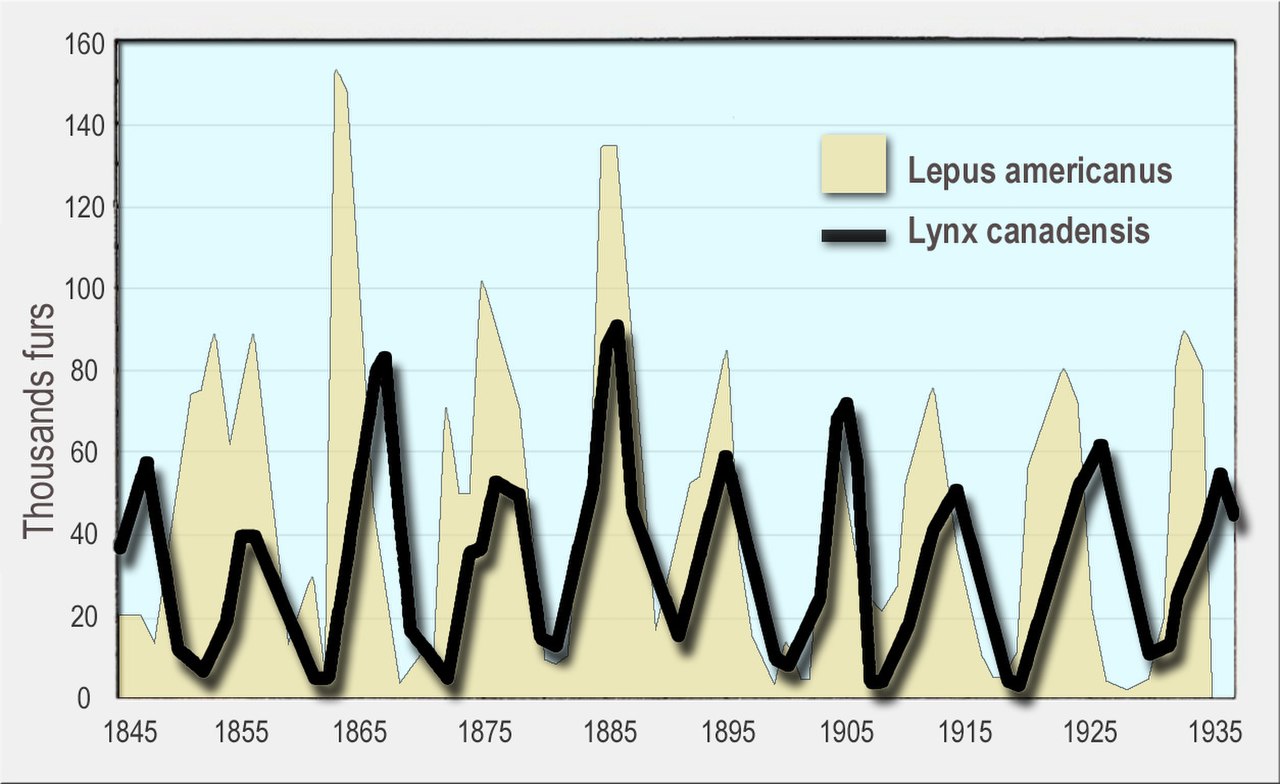
En primer lugar, la dinámica de las poblaciones de depredadores y presas tiende a oscilar. Se han observado números fluctuantes de depredadores y presas en poblaciones naturales, como los datos de linces y liebres con raquetas de nieve de la Compañía de la Bahía de Hudson [6] y las poblaciones de alces y lobos en el Parque Nacional Isle Royale . [7]
En segundo lugar, el equilibrio poblacional de este modelo tiene la propiedad de que la densidad de equilibrio de la presa (dada por ) depende de los parámetros del depredador, y la densidad de equilibrio del depredador (dada por ) de los parámetros de la presa. Esto tiene como consecuencia que un aumento en, por ejemplo, la tasa de crecimiento de las presas, conduce a un aumento en la densidad de equilibrio de los depredadores, pero no en la densidad de equilibrio de las presas. Mejorar el ambiente para la presa beneficia al depredador, no a la presa (esto está relacionado con la paradoja de los pesticidas y con la paradoja del enriquecimiento ). Una demostración de este fenómeno la proporciona el mayor porcentaje de peces depredadores capturados durante los años de la Primera Guerra Mundial (1914-18), cuando la tasa de crecimiento de las presas aumentó debido a un menor esfuerzo pesquero.
Otro ejemplo lo constituye la fertilización experimental del océano con hierro. En varios experimentos se disolvieron grandes cantidades de sales de hierro en el océano. La expectativa era que el hierro, que es un nutriente limitante para el fitoplancton, impulsaría el crecimiento del fitoplancton y secuestraría dióxido de carbono de la atmósfera. La adición de hierro suele provocar una breve floración del fioplancton, que es rápidamente consumido por otros organismos (como peces pequeños o zooplancton ) y limita el efecto del enriquecimiento principalmente a una mayor densidad de depredadores, lo que a su vez limita el secuestro de carbono. Esto es lo que predicen las densidades de población en equilibrio del modelo depredador-presa de Lotka-Volterra, y es una característica que se traslada a modelos más elaborados en los que se relajan los supuestos restrictivos del modelo simple. [8]
El modelo Lotka Volterra tiene aplicaciones adicionales en áreas como la economía [9] y el marketing. [10] [11] Puede usarse para describir la dinámica en un mercado con varios competidores, plataformas y productos complementarios, una economía colaborativa y más. Hay situaciones en las que uno de los competidores expulsa a los demás competidores del mercado y otras situaciones en las que el mercado alcanza un equilibrio en el que cada empresa estabiliza su participación de mercado. También es posible describir situaciones en las que existen cambios cíclicos en la industria o situaciones caóticas sin equilibrio y los cambios son frecuentes e impredecibles.
El modelo depredador-presa de Lotka-Volterra fue propuesto inicialmente por Alfred J. Lotka en la teoría de reacciones químicas autocatalíticas en 1910. [12] [13] Esta era efectivamente la ecuación logística , [14] derivada originalmente por Pierre François Verhulst . [15] En 1920 Lotka amplió el modelo, a través de Andrey Kolmogorov , a "sistemas orgánicos" utilizando una especie vegetal y una especie animal herbívora como ejemplo [16] y en 1925 utilizó las ecuaciones para analizar las interacciones depredador-presa en su libro. sobre biomatemáticas . [17] El mismo conjunto de ecuaciones fue publicado en 1926 por Vito Volterra , un matemático y físico, que se había interesado en la biología matemática . [13] [18] [19] La investigación de Volterra se inspiró en sus interacciones con el biólogo marino Umberto D'Ancona , quien estaba cortejando a su hija en ese momento y más tarde se convertiría en su yerno. D'Ancona estudió las capturas de peces en el mar Adriático y notó que el porcentaje de peces depredadores capturados había aumentado durante los años de la Primera Guerra Mundial (1914-18). Esto lo desconcertó, ya que el esfuerzo de pesca se había reducido mucho durante los años de la guerra y, como los peces de presa eran la captura preferida, uno intuitivamente esperaría que esto aumentara el porcentaje de peces de presa. Volterra desarrolló su modelo para explicar la observación de D'Ancona y lo hizo independientemente de Alfred Lotka. Le dio crédito al trabajo anterior de Lotka en su publicación, después de lo cual el modelo pasó a ser conocido como el "modelo Lotka-Volterra". [20]
Posteriormente, el modelo se amplió para incluir el crecimiento de presas dependiente de la densidad y una respuesta funcional de la forma desarrollada por CS Holling ; un modelo que se conoce como modelo Rosenzweig-MacArthur. [21] Tanto el modelo de Lotka-Volterra como el de Rosenzweig-MacArthur se han utilizado para explicar la dinámica de las poblaciones naturales de depredadores y presas.
A finales de la década de 1980, surgió una alternativa al modelo depredador-presa de Lotka-Volterra (y sus generalizaciones dependientes de presas comunes), el modelo dependiente de la proporción o modelo Arditi-Ginzburg . [22] La validez de los modelos dependientes de la presa o de la proporción ha sido muy debatida. [23]
Las ecuaciones de Lotka-Volterra tienen una larga historia de uso en la teoría económica ; su solicitud inicial se atribuye comúnmente a Richard Goodwin en 1965 [24] o 1967. [25] [26]
Las ecuaciones tienen soluciones periódicas . Estas soluciones no tienen una expresión sencilla en términos de las funciones trigonométricas habituales , aunque son bastante manejables. [27] [28] [29]
Si ninguno de los parámetros no negativos α , β , γ , δ desaparece, se pueden absorber tres en la normalización de variables para dejar solo un parámetro: dado que la primera ecuación es homogénea en x y la segunda en y , los parámetros β / α y δ / γ son absorbibles en las normalizaciones de y y x respectivamente, y γ en la normalización de t , de modo que sólo α / γ permanece arbitrario. Es el único parámetro que afecta la naturaleza de las soluciones.
Una linealización de las ecuaciones produce una solución similar al movimiento armónico simple [30] con la población de depredadores siguiendo a la de presas 90° en el ciclo.

.svg/1280px-Lotka-Volterra_model_(1.1,_0.4,_0.4,_0.1).svg.png)

Supongamos que hay dos especies de animales, un conejo (presa) y un zorro (depredador). Si las densidades iniciales son 10 conejos y 10 zorros por kilómetro cuadrado, se puede trazar la progresión de las dos especies a lo largo del tiempo; dados los parámetros que las tasas de crecimiento y muerte de los conejos son 1,1 y 0,4 mientras que las de los zorros son 0,1 y 0,4 respectivamente. La elección del intervalo de tiempo es arbitraria.
También se pueden trazar soluciones paramétricamente como órbitas en el espacio de fases , sin representar el tiempo, pero con un eje representando el número de presas y el otro eje representando las densidades de depredadores para todos los tiempos.
Esto corresponde a eliminar el tiempo de las dos ecuaciones diferenciales anteriores para producir una única ecuación diferencial.
relacionando las variables x e y . Las soluciones de esta ecuación son curvas cerradas. Es susceptible de separación de variables : integración
produce la relación implícita
donde V es una cantidad constante que depende de las condiciones iniciales y se conserva en cada curva.
Una acotación: estos gráficos ilustran una limitación potencial grave en la aplicación como modelo biológico: para esta elección específica de parámetros, en cada ciclo, la población de conejos se reduce a números extremadamente bajos, pero se recupera (mientras que la población de zorros sigue siendo considerable al final). densidad de conejos más baja). Sin embargo, en situaciones de la vida real, las fluctuaciones aleatorias de los números discretos de individuos podrían causar que los conejos se extingan y, en consecuencia, también los zorros. Este problema de modelado se ha denominado "problema del atto-zorro", siendo un atto- zorro un 10 −18 teórico de un zorro. [31] [32] Una densidad de 10 −18 zorros por kilómetro cuadrado equivale a un promedio de aproximadamente 5×10 −10 zorros en la superficie de la tierra, lo que en términos prácticos significa que los zorros están extintos.
Dado que la cantidad se conserva en el tiempo, desempeña el papel de una función hamiltoniana del sistema. [33] Para ver esto podemos definir el corchete de Poisson de la siguiente manera . Entonces las ecuaciones de Hamilton leen

Un ejemplo menos extremo cubre:
α = 2/3 , β = 4/3 , γ = 1 = δ . Supongamos que x , y cuantifican miles cada uno. Los círculos representan las condiciones iniciales de presa y depredador desde x = y = 0,9 a 1,8, en pasos de 0,1. El punto fijo está en (1, 1/2).
En el sistema modelo, los depredadores prosperan cuando las presas son abundantes pero, en última instancia, superan su suministro de alimentos y disminuyen. A medida que la población de depredadores sea baja, la población de presas aumentará nuevamente. Estas dinámicas continúan en un ciclo poblacional de crecimiento y disminución.
El equilibrio de la población ocurre en el modelo cuando ninguno de los niveles de población cambia, es decir, cuando ambas derivadas son iguales a 0:
El sistema de ecuaciones anterior produce dos soluciones:
Por tanto, hay dos equilibrios.
La primera solución representa efectivamente la extinción de ambas especies. Si ambas poblaciones están en 0, seguirán estando así indefinidamente. La segunda solución representa un punto fijo en el que ambas poblaciones mantienen sus números actuales distintos de cero y, en el modelo simplificado, lo hacen indefinidamente. Los niveles de población en los que se logra este equilibrio dependen de los valores elegidos de los parámetros α , β , γ y δ .
La estabilidad del punto fijo en el origen se puede determinar realizando una linealización utilizando derivadas parciales .
La matriz jacobiana del modelo depredador-presa es
Cuando se evalúa en el estado estacionario de (0, 0) , la matriz jacobiana J se convierte en
Los valores propios de esta matriz son
En el modelo, α y γ son siempre mayores que cero y, como tal, el signo de los valores propios anteriores siempre será diferente. Por tanto, el punto fijo en el origen es un punto de silla .
La inestabilidad de este punto fijo es importante. Si fuera estable, poblaciones distintas de cero podrían verse atraídas hacia él y, como tal, la dinámica del sistema podría conducir a la extinción de ambas especies en muchos casos de niveles poblacionales iniciales. Sin embargo, como el punto fijo en el origen es un punto de silla y, por tanto, inestable, se deduce que la extinción de ambas especies es difícil en el modelo. (De hecho, esto sólo podría ocurrir si la presa fuera completamente erradicada artificialmente, provocando que los depredadores murieran de hambre. Si los depredadores fueran erradicados, la población de presas crecería sin límites en este modelo simple.) Las poblaciones de presas y depredadores pueden acercarse infinitamente a cero y aun así recuperarse.
La evaluación de J en el segundo punto fijo conduce a
Los valores propios de esta matriz son
Como los valores propios son puramente imaginarios y conjugados entre sí, este punto fijo debe ser un centro de órbitas cerradas en las proximidades locales o una espiral atractiva o repulsiva. En sistemas conservadores, debe haber órbitas cerradas en las proximidades locales de puntos fijos que existen en los mínimos y máximos de la cantidad conservada. La cantidad conservada se deduce arriba para estar en órbitas. Así, las órbitas alrededor del punto fijo son cerradas y elípticas , por lo que las soluciones son periódicas, oscilando en una pequeña elipse alrededor del punto fijo, con una frecuencia y un período .
Como se ilustra en las oscilaciones circulantes en la figura anterior, las curvas de nivel son órbitas cerradas que rodean el punto fijo: los niveles de las poblaciones de depredadores y presas ciclan y oscilan sin amortiguación alrededor del punto fijo con frecuencia .
El valor de la constante de movimiento V , o, equivalentemente, K = exp(− V ) , se puede encontrar para las órbitas cerradas cerca del punto fijo.
Al aumentar K, se acerca una órbita cerrada al punto fijo. El mayor valor de la constante K se obtiene resolviendo el problema de optimización.



































