
La cóclea es la parte del oído interno implicada en la audición . Es una cavidad en forma de espiral en el laberinto óseo , que en los humanos realiza 2,75 vueltas alrededor de su eje, el modiolo . [2] [3] Un componente central de la cóclea es el órgano de Corti , el órgano sensorial de la audición, que se distribuye a lo largo del tabique que separa las cámaras de líquido en el tubo cónico en espiral de la cóclea.
El nombre cóclea deriva del griego antiguo κοχλίας (kokhlias) 'espiral, concha de caracol'.
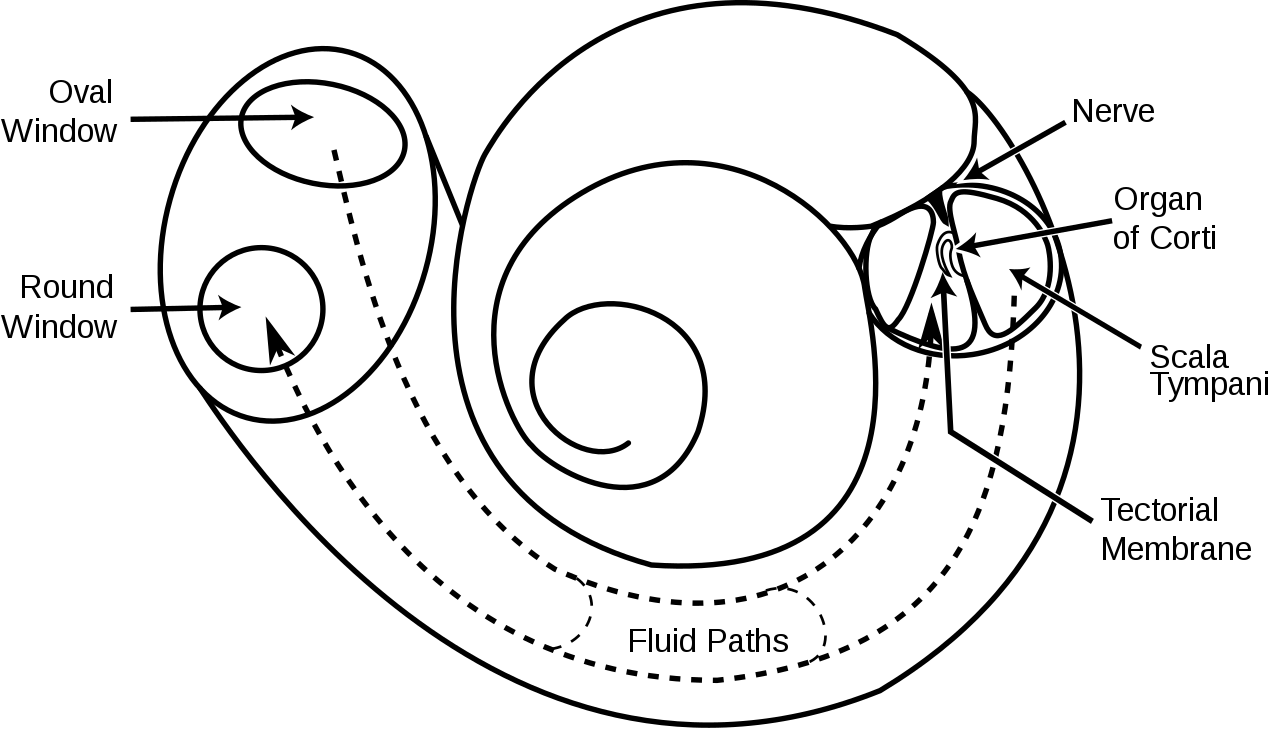
La cóclea ( pl.: cochleae) es una cámara ósea cónica, hueca y en espiral, en la que las ondas se propagan desde la base (cerca del oído medio y la ventana oval ) hasta el ápice (la parte superior o centro de la espiral). El canal espiral de la cóclea es una sección del laberinto óseo del oído interno que mide aproximadamente 30 mm de largo y da 2 3 ⁄ 4 vueltas alrededor del modiolo. Las estructuras cocleares incluyen:
La cóclea es una parte del oído interno que parece una concha de caracol ( coclea es la palabra griega para caracol). [4] La cóclea recibe el sonido en forma de vibraciones, que hacen que los estereocilios se muevan. Los estereocilios luego convierten estas vibraciones en impulsos nerviosos que son llevados al cerebro para ser interpretados. Dos de las tres secciones de fluido son canales y el tercero es el "órgano de Corti", que detecta los impulsos de presión que viajan a lo largo del nervio auditivo hasta el cerebro. Los dos canales se llaman canal vestibular y canal timpánico.
Las paredes de la cóclea hueca están hechas de hueso, con un revestimiento fino y delicado de tejido epitelial . Este tubo enrollado está dividido en la mayor parte de su longitud por una partición membranosa interna. Dos espacios exteriores llenos de líquido (conductos o escalas ) están formados por esta membrana divisoria. En la parte superior de los tubos enrollados en forma de concha de caracol, hay una inversión de la dirección del líquido, cambiando así el conducto vestibular al conducto timpánico. Esta área se llama helicotrema. Esta continuación en el helicotrema permite que el líquido que es empujado hacia el conducto vestibular por la ventana oval vuelva a salir a través del movimiento en el conducto timpánico y la desviación de la ventana redonda; dado que el líquido es casi incompresible y las paredes óseas son rígidas, es esencial que el volumen de líquido conservado salga por algún lado.
El tabique longitudinal que divide la mayor parte de la cóclea es en sí mismo un tubo lleno de líquido, el tercer "conducto". Esta columna central se llama conducto coclear. Su líquido, la endolinfa, también contiene electrolitos y proteínas, pero es químicamente muy diferente de la perilinfa. Mientras que la perilinfa es rica en iones de sodio, la endolinfa es rica en iones de potasio, lo que produce un potencial eléctrico iónico .
Las células ciliadas están dispuestas en cuatro filas en el órgano de Corti a lo largo de toda la espiral coclear. Tres filas están formadas por células ciliadas externas (CCE) y una fila está formada por células ciliadas internas (CCI). Las células ciliadas internas proporcionan la principal salida neuronal de la cóclea. Las células ciliadas externas, en cambio, "reciben" principalmente la entrada neuronal del cerebro, que influye en su motilidad como parte del "preamplificador" mecánico de la cóclea. La entrada a las CCE proviene del cuerpo olivar a través del haz olivococlear medial.
El conducto coclear es casi tan complejo por sí solo como el oído mismo. El conducto coclear está delimitado por tres lados por la membrana basilar , la estría vascular y la membrana de Reissner. La estría vascular es un lecho rico en capilares y células secretoras; la membrana de Reissner es una membrana delgada que separa la endolinfa de la perilinfa; y la membrana basilar es una membrana mecánicamente algo rígida, que sostiene el órgano receptor de la audición, el órgano de Corti, y determina las propiedades de propagación de ondas mecánicas del sistema coclear.
Entre los hombres y las mujeres, existen diferencias en la forma de la cóclea humana. La variación está en la torsión al final de la espiral. Debido a esta diferencia, y debido a que la cóclea es uno de los huesos más duraderos del cráneo, se utiliza para determinar el sexo de los restos humanos encontrados en yacimientos arqueológicos. [5] [6]
La cóclea está llena de un líquido acuoso, la endolinfa , que se mueve en respuesta a las vibraciones que llegan desde el oído medio a través de la ventana oval. A medida que el líquido se mueve, el tabique coclear (membrana basilar y órgano de Corti) se mueve; miles de células ciliadas detectan el movimiento a través de sus estereocilios y convierten ese movimiento en señales eléctricas que se comunican mediante neurotransmisores a muchos miles de células nerviosas. Estas neuronas auditivas primarias transforman las señales en impulsos electroquímicos conocidos como potenciales de acción , que viajan a lo largo del nervio auditivo hasta las estructuras del tronco encefálico para su posterior procesamiento.
El hueso del estribo (hueso del estribo) del oído medio transmite vibraciones a la fenestra oval (ventana oval) en el exterior de la cóclea, que hace vibrar la perilinfa en el conducto vestibular (cámara superior de la cóclea). Los huesecillos son esenciales para el acoplamiento eficiente de las ondas sonoras en la cóclea, ya que el entorno de la cóclea es un sistema de membrana-líquido y se necesita más presión para mover el sonido a través de las ondas de membrana-líquido que a través del aire. Se logra un aumento de presión al reducir la relación del área de la membrana timpánica (tímpano) a la ventana oval ( hueso del estribo ) en 20. Como presión = fuerza/área, resulta en una ganancia de presión de aproximadamente 20 veces con respecto a la presión de la onda sonora original en el aire. Esta ganancia es una forma de adaptación de impedancia : para hacer coincidir la onda sonora que viaja a través del aire con la que viaja en el sistema de membrana-líquido.
En la base de la cóclea, cada "conducto" termina en un portal membranoso que mira hacia la cavidad del oído medio: el conducto vestibular termina en la ventana oval , donde se asienta la plataforma del estribo . La plataforma vibra cuando la presión se transmite a través de la cadena osicular. La onda en la perilinfa se aleja de la plataforma y se dirige hacia el helicotrema . Dado que esas ondas fluidas mueven el tabique coclear que separa los conductos hacia arriba y hacia abajo, las ondas tienen una parte simétrica correspondiente en la perilinfa del conducto timpánico, que termina en la ventana redonda, abultándose cuando la ventana oval se abulta hacia adentro.
La perilinfa del conducto vestibular y la endolinfa del conducto coclear actúan mecánicamente como un solo conducto, manteniéndose separadas únicamente por la muy delgada membrana de Reissner . Las vibraciones de la endolinfa del conducto coclear desplazan la membrana basilar en un patrón que alcanza su punto máximo a cierta distancia de la ventana oval, dependiendo de la frecuencia de la onda sonora. El órgano de Corti vibra debido a que las células pilosas externas amplifican aún más estas vibraciones. Las células pilosas internas son entonces desplazadas por las vibraciones del líquido y se despolarizan por un influjo de K+ a través de sus canales conectados por el enlace de punta , y envían sus señales a través de neurotransmisores a las neuronas auditivas primarias del ganglio espiral . [7]
Las células pilosas del órgano de Corti están sintonizadas con ciertas frecuencias de sonido por su ubicación en la cóclea, debido al grado de rigidez de la membrana basilar. [8] Esta rigidez se debe, entre otras cosas, al grosor y ancho de la membrana basilar, [9] que a lo largo de la cóclea es más rígida cerca de su inicio en la ventana oval, donde el estribo introduce las vibraciones que vienen del tímpano. Como su rigidez es alta allí, solo permite que las vibraciones de alta frecuencia muevan la membrana basilar y, por lo tanto, las células pilosas. Cuanto más viaja una onda hacia el ápice de la cóclea (el helicotrema ), menos rígida es la membrana basilar; por lo tanto, las frecuencias más bajas viajan por el tubo, y la membrana menos rígida se mueve más fácilmente por ellas donde la rigidez reducida lo permite: es decir, a medida que la membrana basilar se vuelve cada vez menos rígida, las ondas se ralentizan y responde mejor a las frecuencias más bajas. Además, en los mamíferos, la cóclea está enrollada, lo que se ha demostrado que mejora las vibraciones de baja frecuencia a medida que viajan a través de la bobina llena de líquido. [10] Esta disposición espacial de la recepción del sonido se conoce como tonotopía .
En el caso de frecuencias muy bajas (por debajo de los 20 Hz), las ondas se propagan a lo largo de todo el recorrido de la cóclea, de forma diferencial por el conducto vestibular y el conducto timpánico hasta llegar al helicotrema . Las frecuencias tan bajas siguen activando el órgano de Corti en cierta medida, pero son demasiado bajas para provocar la percepción de un tono . Las frecuencias más altas no se propagan al helicotrema debido a la tonotopía mediada por la rigidez.
Un movimiento muy fuerte de la membrana basilar debido a un ruido muy fuerte puede provocar la muerte de las células ciliadas. Esta es una causa común de pérdida auditiva parcial y es la razón por la que los usuarios de armas de fuego o maquinaria pesada suelen usar orejeras o tapones para los oídos .
Para transmitir la sensación del sonido al cerebro, donde puede procesarse y convertirse en la percepción auditiva , las células ciliadas de la cóclea deben convertir su estimulación mecánica en patrones de señalización eléctrica del sistema nervioso. Las células ciliadas son neuronas modificadas , capaces de generar potenciales de acción que pueden transmitirse a otras células nerviosas. Estas señales de potencial de acción viajan a través del nervio vestibulococlear para finalmente llegar al bulbo raquídeo anterior , donde hacen sinapsis y se procesan inicialmente en los núcleos cocleares . [11]
Parte del procesamiento ocurre en los propios núcleos cocleares, pero las señales también deben viajar al complejo olivar superior del puente , así como a los colículos inferiores para su posterior procesamiento. [11]
La cóclea no sólo "recibe" sonidos, sino que una cóclea sana los genera y amplifica cuando es necesario. Cuando el organismo necesita un mecanismo para oír sonidos muy débiles, la cóclea los amplifica mediante la transducción inversa de las células OHC, convirtiendo las señales eléctricas de nuevo en mecánicas en una configuración de retroalimentación positiva. Las células OHC tienen un motor proteico llamado prestina en sus membranas externas; genera movimiento adicional que se acopla de nuevo a la onda de fluido-membrana. Este "amplificador activo" es esencial para la capacidad del oído de amplificar sonidos débiles. [12] [13]
El amplificador activo también provoca el fenómeno de vibraciones de ondas sonoras que se emiten desde la cóclea hacia el canal auditivo a través del oído medio (emisiones otoacústicas).
Las otoemisiones acústicas se deben a una onda que sale de la cóclea a través de la ventana oval y se propaga de nuevo a través del oído medio hasta el tímpano y sale por el canal auditivo, donde puede ser captada por un micrófono. Las otoemisiones acústicas son importantes en algunos tipos de pruebas de pérdida auditiva , ya que están presentes cuando la cóclea está funcionando bien y menos cuando sufre una pérdida de actividad de la CCE. Las otoemisiones acústicas también presentan dimorfismos sexuales, ya que las mujeres tienden a mostrar magnitudes más altas de otoemisiones acústicas. Los hombres tienden a experimentar una reducción en las magnitudes de las otoemisiones acústicas a medida que envejecen. Las mujeres, por otro lado, no experimentan un cambio en las magnitudes de las otoemisiones acústicas con la edad. [14]
Las proteínas de unión en hendidura, llamadas conexinas , expresadas en la cóclea desempeñan un papel importante en el funcionamiento auditivo. [15] Se ha descubierto que las mutaciones en los genes de unión en hendidura causan sordera sindrómica y no sindrómica. [16] Ciertas conexinas, incluidas la conexina 30 y la conexina 26 , prevalecen en los dos sistemas de unión en hendidura distintos que se encuentran en la cóclea. La red de unión en hendidura de células epiteliales acopla células epiteliales no sensoriales, mientras que la red de unión en hendidura de tejido conectivo acopla células de tejido conectivo. [17] Los canales de unión en hendidura reciclan iones de potasio de regreso a la endolinfa después de la mecanotransducción en las células ciliadas . [18] Es importante destacar que los canales de unión en hendidura se encuentran entre las células de soporte cocleares, pero no entre las células ciliadas auditivas . [19]
El daño a la cóclea puede ser resultado de diferentes incidentes o condiciones, como una lesión grave en la cabeza, un colesteatoma , una infección y/o exposición a ruidos fuertes que podrían matar las células ciliadas de la cóclea.
La pérdida auditiva asociada a la cóclea suele ser resultado de daño o muerte de las células pilosas externas e internas. Las células pilosas externas son más susceptibles a sufrir daños, lo que puede resultar en una menor sensibilidad a los sonidos débiles. La sensibilidad a la frecuencia también se ve afectada por el daño coclear, lo que puede perjudicar la capacidad del paciente para distinguir entre las diferencias espectrales de las vocales. Los efectos del daño coclear en diferentes aspectos de la pérdida auditiva, como la integración temporal, la percepción del tono y la determinación de la frecuencia, aún se están estudiando, dado que se deben tener en cuenta múltiples factores en relación con la investigación coclear. [20]
En 2009, ingenieros del Instituto Tecnológico de Massachusetts crearon un chip electrónico que puede analizar rápidamente una amplia gama de frecuencias de radio utilizando sólo una fracción de la energía necesaria para las tecnologías existentes; su diseño imita específicamente una cóclea. [21] [22]
La forma enrollada de la cóclea es exclusiva de los mamíferos . En las aves y en otros vertebrados no mamíferos , el compartimento que contiene las células sensoriales para la audición a veces también se llama "cóclea", a pesar de que no está enrollado. En cambio, forma un tubo con extremos ciegos, también llamado conducto coclear. Esta diferencia aparentemente evolucionó en paralelo con las diferencias en el rango de frecuencia de la audición entre mamíferos y vertebrados no mamíferos. El rango de frecuencia superior en los mamíferos se debe en parte a su mecanismo único de preamplificación del sonido por vibraciones activas del cuerpo celular de las células ciliadas externas . Sin embargo, la resolución de frecuencia no es mejor en los mamíferos que en la mayoría de los lagartos y aves, pero el límite superior de frecuencia es, a veces, mucho más alto. La mayoría de las especies de aves no oyen por encima de 4-5 kHz, y el máximo conocido actualmente es de ~ 11 kHz en la lechuza común. Algunos mamíferos marinos oyen hasta 200 kHz. Un compartimento largo y enrollado, en lugar de uno corto y recto, proporciona más espacio para octavas adicionales de rango auditivo y ha hecho posible algunos de los comportamientos altamente derivados que involucran la audición de los mamíferos. [23]
Como el estudio de la cóclea debe centrarse fundamentalmente en el nivel de las células pilosas, es importante observar las diferencias anatómicas y fisiológicas entre las células pilosas de las distintas especies. En las aves, por ejemplo, en lugar de células pilosas externas e internas, hay células pilosas altas y cortas. Hay varias similitudes notables con respecto a estos datos comparativos. Por un lado, la célula pilosa alta es muy similar en función a la célula pilosa interna, y la célula pilosa corta, que carece de inervación de fibras nerviosas auditivas aferentes, se parece a la célula pilosa externa. Sin embargo, una diferencia inevitable es que, mientras que todas las células pilosas están unidas a una membrana tectorial en las aves, solo las células pilosas externas están unidas a la membrana tectorial en los mamíferos.
El nombre cóclea se deriva de la palabra latina para concha de caracol , que a su vez proviene del griego κοχλίας kokhlias ("caracol, tornillo"), de κόχλος kokhlos ("concha en espiral") [24] en referencia a su forma enrollada; la cóclea está enrollada en los mamíferos con excepción de los monotremas .




{{cite book}}: Mantenimiento de CS1: otros ( enlace )