
Las células vegetales son las células presentes en las plantas verdes , eucariotas fotosintéticos del reino Plantae . Sus características distintivas incluyen paredes celulares primarias que contienen celulosa, hemicelulosas y pectina, la presencia de plastidios con capacidad de realizar la fotosíntesis y almacenar almidón, una gran vacuola que regula la presión de turgencia, la ausencia de flagelos o centriolos , excepto en los gametos, y una Método único de división celular que implica la formación de una placa celular o fragmoplasto que separa las nuevas células hijas.
Las células vegetales se diferencian de las células meristemáticas indiferenciadas (análogas a las células madre de los animales) para formar las principales clases de células y tejidos de raíces , tallos , hojas , flores y estructuras reproductivas, cada una de las cuales puede estar compuesta por varios tipos de células.
Las células del parénquima son células vivas que tienen funciones que van desde el almacenamiento y soporte hasta la fotosíntesis ( células del mesófilo ) y la carga del floema ( células de transferencia ). Además del xilema y el floema en sus haces vasculares, las hojas se componen principalmente de células parenquimatosas. Algunas células del parénquima, como en la epidermis, están especializadas en la penetración de la luz y en la concentración o regulación del intercambio de gases , pero otras se encuentran entre las células menos especializadas del tejido vegetal y pueden seguir siendo totipotentes , capaces de dividirse para producir nuevas poblaciones de células indiferenciadas. A lo largo de sus vidas. [17] Las células del parénquima tienen paredes primarias delgadas y permeables que permiten el transporte de pequeñas moléculas entre ellas, y su citoplasma es responsable de una amplia gama de funciones bioquímicas como la secreción de néctar o la fabricación de productos secundarios que desalentan la herbivoría . Las células del parénquima que contienen muchos cloroplastos y se ocupan principalmente de la fotosíntesis se denominan células de clorénquima . Las células de clorénquima son células del parénquima implicadas en la fotosíntesis. [18] Otros, como la mayoría de las células del parénquima de los tubérculos de patata y los cotiledones de las semillas de las legumbres , tienen una función de almacenamiento.
Las células del colénquima están vivas en la madurez y tienen paredes celulares de celulosa engrosadas. [19] Estas células maduran a partir de derivados de meristemo que inicialmente se parecen al parénquima, pero las diferencias rápidamente se vuelven evidentes. Los plastidios no se desarrollan y el aparato secretor (RE y Golgi) prolifera para secretar pared primaria adicional. La pared suele ser más gruesa en las esquinas, donde entran en contacto tres o más células, y más delgada donde sólo dos células entran en contacto, aunque son posibles otras disposiciones del engrosamiento de la pared. [19] La pectina y la hemicelulosa son los componentes dominantes de las paredes celulares del colénquima de las angiospermas dicotiledóneas , que pueden contener tan solo un 20% de celulosa en las petasites . [20] Las células del colénquima suelen ser bastante alargadas y pueden dividirse transversalmente para dar una apariencia septada. La función de este tipo de células es sostener la planta en ejes que aún crecen en longitud y conferir flexibilidad y resistencia a la tracción a los tejidos. La pared primaria carece de lignina, lo que la haría resistente y rígida, por lo que este tipo de células proporciona lo que podría llamarse soporte plástico: soporte que puede sostener un tallo o pecíolo joven en el aire, pero en células que pueden estirarse como las células que los rodean. alargado. El soporte estirable (sin cierre elástico) es una buena manera de describir lo que hace el colénquima. Partes de las cuerdas del apio son colénquima.
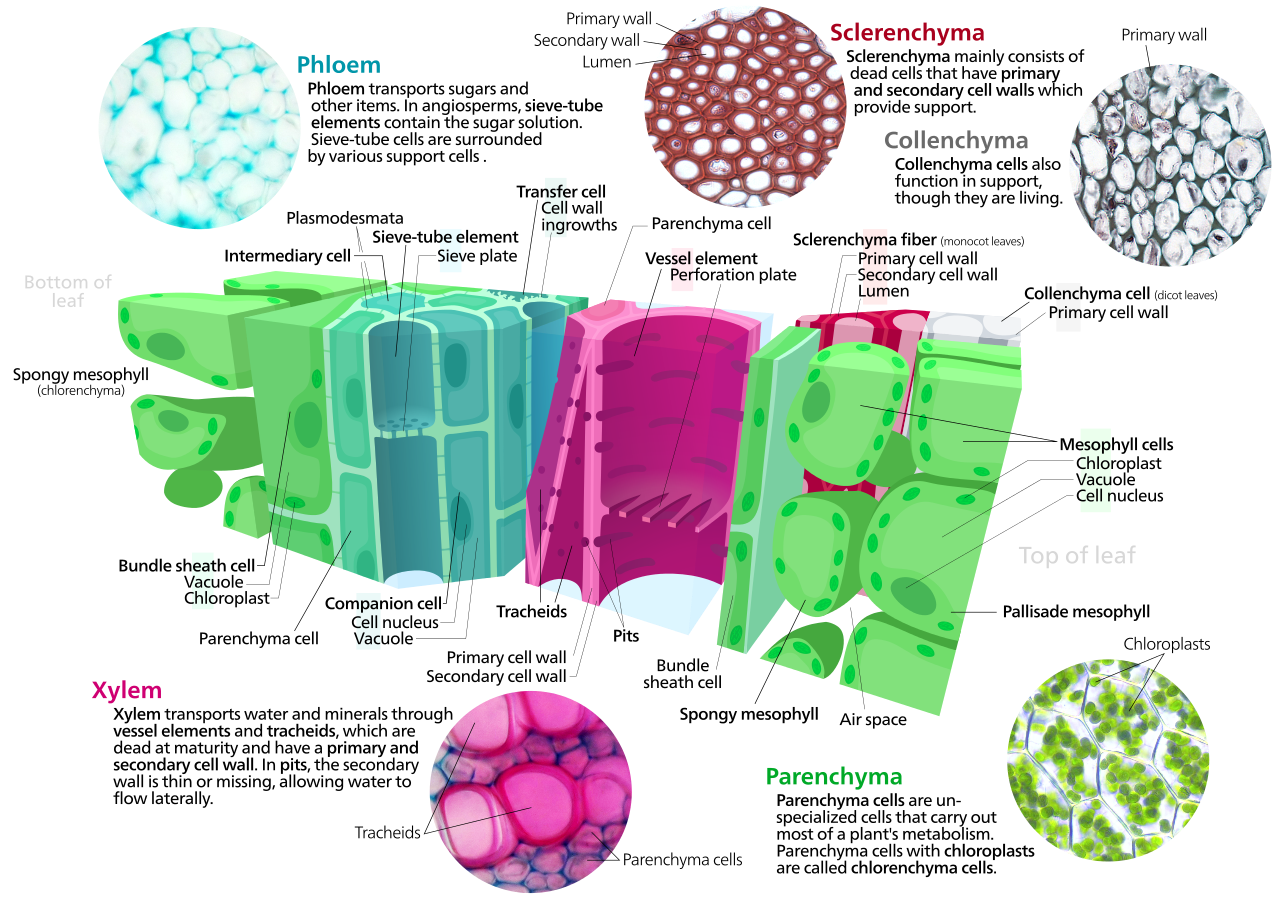
El esclerénquima es un tejido compuesto por dos tipos de células, esclereidas y fibras que tienen paredes secundarias engrosadas y lignificadas [19] : 78 colocadas dentro de la pared celular primaria . Las paredes secundarias endurecen las células y las hacen impermeables al agua. En consecuencia, las esclereidas y las fibras suelen estar muertas en la madurez funcional y falta el citoplasma, lo que deja una cavidad central vacía. Las esclereidas o células pétreas (del griego skleros, duras ) son células duras y resistentes que dan a las hojas o frutos una textura arenosa. Pueden desalentar la herbivoría al dañar los conductos digestivos en las etapas larvarias de insectos pequeños. Las esclereidas forman la dura pared del hueso de los melocotones y muchas otras frutas, proporcionando protección física al grano en desarrollo. Las fibras son células alargadas con paredes secundarias lignificadas que proporcionan soporte de carga y resistencia a la tracción a las hojas y tallos de las plantas herbáceas. Las fibras del esclerénquima no participan en la conducción, ni de agua y nutrientes (como en el xilema ) ni de compuestos de carbono (como en el floema ), pero es probable que evolucionaran como modificaciones de las iniciales de xilema y floema en las primeras plantas terrestres.

El xilema es un tejido vascular complejo compuesto por traqueidas o elementos vasculares conductores de agua , junto con fibras y células de parénquima. Las traqueidas [21] son células alargadas con engrosamiento secundario lignificado de las paredes celulares, especializadas en la conducción de agua, y aparecieron por primera vez en las plantas durante su transición a la tierra en el período Silúrico hace más de 425 millones de años (ver Cooksonia ). La posesión de traqueidas xilemáticas define a las plantas vasculares o traqueofitas. Las traqueidas son células del xilema alargadas y puntiagudas, las más simples de las cuales tienen paredes celulares primarias continuas y engrosamientos lignificados de la pared secundaria en forma de anillos, aros o redes reticuladas. Las traqueidas más complejas con perforaciones en forma de válvulas llamadas fosas bordeadas caracterizan a las gimnospermas. Los helechos y otros pteridofitos y las gimnospermas tienen sólo traqueidas xilemáticas , mientras que las plantas con flores también tienen vasos xilemáticos . Los elementos vasculares son células huecas del xilema sin paredes extremas que están alineadas de extremo a extremo para formar tubos largos y continuos. Los briofitos carecen de tejido xilemático verdadero, pero sus esporofitos tienen un tejido conductor de agua conocido como hidroma que está compuesto por células alargadas de construcción más simple.
El floema es un tejido especializado para el transporte de alimentos en las plantas superiores, transportando principalmente sacarosa a lo largo de gradientes de presión generados por ósmosis, proceso llamado translocación . El floema es un tejido complejo, que consta de dos tipos de células principales, los tubos cribosos y las células acompañantes íntimamente asociadas , junto con las células del parénquima, las fibras del floema y las esclereidas. [19] : 171 Los tubos cribosos están unidos de extremo a extremo con placas extremas perforadas conocidas como placas cribosas , que permiten el transporte de fotosintato entre los elementos cribosos. Los elementos del tubo criboso carecen de núcleos y ribosomas , y su metabolismo y funciones están regulados por las células nucleadas acompañantes adyacentes. Las células compañeras, conectadas a los tubos cribosos a través de plasmodesmos , son responsables de cargar el floema con azúcares . Los briofitos carecen de floema, pero los esporofitos de musgo tienen un tejido más simple con función análoga conocido como leptoma.

La epidermis vegetal es un tejido especializado, compuesto por células de parénquima, que recubre las superficies externas de hojas, tallos y raíces. Pueden estar presentes varios tipos de células en la epidermis. Entre ellos se destacan las células protectoras de los estomas que controlan la tasa de intercambio de gases entre la planta y la atmósfera, los pelos o tricomas glandulares y de la ropa , y los pelos radiculares de las raíces primarias. En la epidermis de los brotes de la mayoría de las plantas, sólo las células protectoras tienen cloroplastos. Los cloroplastos contienen el pigmento verde clorofila, necesario para la fotosíntesis. Las células epidérmicas de los órganos aéreos surgen de la capa superficial de células conocida como túnica (capas L1 y L2) que cubre el ápice del brote de la planta , [19] mientras que la corteza y los tejidos vasculares surgen de la capa más interna del ápice del brote conocida como corpus (capa L3). La epidermis de las raíces se origina en la capa de células inmediatamente debajo de la cofia de la raíz. La epidermis de todos los órganos aéreos, pero no de las raíces, está cubierta por una cutícula hecha de cutina de poliéster o cután de polímero (o ambos), con una capa superficial de ceras epicuticulares . Se cree que las células epidérmicas del brote primario son las únicas células vegetales con capacidad bioquímica para sintetizar cutina. [22]
{{cite book}}: Mantenimiento CS1: falta el editor de la ubicación ( enlace )